
Abstract
17β-Estradiol (E2) was shown to exert neuroprotective effects both in in vitro and in vivo models of stroke. Although these effects of E2 are known to require estrogen receptor-α (ERα), the cellular target of estrogen-mediated neuroprotection remains unknown. Using cell type-specific ER mutant mice in an in vivo model of stroke, we specifically investigated the role of ERα in neuronal cells versus its role in the microglia in the mediation of neuroprotection by estrogens. We generated and analyzed two different tissue-specific knockout mouse lines lacking ERα either in cells of myeloid lineage, including microglia, or in the neurons of the forebrain. Both E2-treated and E2-untreated mutant and control mice were subjected to a permanent middle cerebral artery occlusion for 48 h, and the infarct volume was quantified. Although the infarct volume of E2-treated female myeloid-specific ERα knockout mice was similar to that of E2-treated control mice, both male and female neuron-specific ERα mutant mice had larger infarcts than did control mice after E2 treatment. We conclude that neuronal ERα in female and male mice mediates neuroprotective estrogen effects in an in vivo mouse model of stroke, whereas microglial ERα is dispensable.
Introduction
17β-Estradiol (E2) has been shown to have protective effects against brain injury and neurodegeneration in several in vivo studies (Merchenthaler et al, 2003). 17β-Estradiol treatment of animals leads to smaller infarct size compared with untreated animals (Dubal et al, 1999). Studies in genetically modified animals showed that estrogen receptor-α (ERα) is essential for neuroprotection by estrogens in models of middle cerebral artery occlusion (MCAO) (Dubal et al, 2001). However, the cellular mechanism remains elusive, as several estrogen-responsive cell types could mediate this protective effect. In vitro studies showed direct protective actions of E2 on neurons, e.g., in serum deprivation and glutamate-induced excitotoxicity (McEwen and Alves, 1999). Alternatively, microglial cells have been described to have an important role in inflammatory response and apoptosis in acute injury models of the brain both in vitro and in vivo (Lalancette-Hebert et al, 2007; Vegeto et al, 2003). Furthermore, microglial ERα exerts an important antiinflammatory effect in in vitro models of neuronal damage (Ghisletti et al, 2005). To dissect the neuroprotective action of estrogens on a cellular level, we generated two different cell type-specific ERα knockout mouse lines using the Cre-loxP system, deleting ERα in the microglia or in neurons of the brain, respectively, and subjected the mice to permanent focal cerebral ischemia.
Material and methods
Animals
All mouse strains were crossed to a C57BL/6 background. C57BL/6 mice were obtained from Charles River (Sulzfeld, Germany). Crossing the mice to a C57BL/6 background for four generations guarantees a nearly complete C57BL/6 background.
Myeloid-specific ERα mutant mice were generated by breeding mice expressing the Cre recombinase under control of the lysozyme M promoter (Clausen et al, 1999) with mice harboring loxP site-modified ERα alleles (ERfl/fl) (Wintermantel et al, 2006). To generate mice lacking ERα in the neurons of the forebrain, mice expressing the Cre recombinase under control of the CaMKII promoter were bred with ERfl/fl mice (Wintermantel et al, 2006).
Microglial Cell Culture
Microglial cells were isolated from newborn mice (P1) as described previously (Burudi et al, 1999). Flasks (one 75-cm2 flask for three brain samples) were coated with poly-L-lysine (0.01%). The brains of 1-day-old mice were isolated and collected in a cell culture dish containing Hank's balanced salt solution. The meninges were removed. The brains were then homogenized and lysates were incubated for 20 mins at room temperature after adding 1 mL of 1% trypsin. After 20 mins, Falcon tubes containing the cell lysate were filled to a volume of 50 mL with complete medium (complete Dulbecco's modified Eagle's medium (cDMEM): DMEM, 10% fetal bovine serum, 100 Units/mL penicillin, 100 μg/mL streptomycin, and 2 mmol/L glutamine) and centrifuged for 10 mins at 180 g. The precipitate was resuspended with 1 mL cDMEM for every three brains. The cell lysate was distributed with 1 mL per flask and incubated at 37°C, 5% CO2. The medium was changed at days 1, 2, and 7. After 14 days, the secondary culture for selection of microglial cells was performed. All steps were performed at room temperature. Flasks were shaken vigorously to detach oligodendrocytes and microglial cells bound to the surface of the cell layer. The medium was discarded, and the flasks were rinsed once with 10 mL cDMEM each and incubated with 3 mL trypsine-EDTA per flask for 3 mins at room temperature. Trypsine-EDTA was discarded and 10 mL of cDMEM and 0.5 mL of DNase were added. The cells were resuspended and centrifuged for 10 mins at 180 g. The cell precipitate was resuspended with 4 mL of cDMEM. A volume of 1 mL of the resuspended cells was added to a Petri dish and incubated at 37°C, 5% CO2 for 20 mins. After 20 mins of incubation, 6 mL of cDMEM was added to the cultures and the cells were grown at 37°C, 5% CO2. The medium was changed once a week. Microglial cells were used after 3 weeks in vitro for further analysis.
Estrogen Receptor-α Immunocytochemistry
Coverslips with attached microglial cells were washed with phosphate-buffered saline (PBS)/MgCl2 and fixed with 4% paraformaldehyde. Thereafter, the cells were treated for 10 mins with 50 mmol/L NH4Cl/PBS and permeabilized for 15 mins with 0.1% Triton-X-100/PBS. The cells were washed with PBS/MgCl2 and incubated for 20 mins with 5% NSS (normal sheep serum). Incubation with anti-ERα antibody (polyclonal anti-ERα MC-20, Santa Cruz (Heidelberg, Germany) sc-542, diluted 1:500 in 5% NSS) was performed overnight at 4°C. The cells were then washed thrice for 5 mins and incubated for 30 mins with the secondary antibody (anti-rabbit AlexaFluor 594, Invitrogen (Karlsruhe, Germany), diluted 1:500 in 5% NSS). After washing the cells thrice for 5 mins with PBS/MgCl2, they were incubated overnight with isolectin GS-IB4 AlexaFluor 488 from Griffonia simplicifolia (Invitrogen, diluted 1:20 in 5% NSS) at 4°C. Finally, the cells were washed thrice for 5 mins with PBS/MgCl2 and mounted with Mowiol (Roth, Karlsruhe, Germany). Estrogen receptor-α-positive cells and isolectin B4-positive cells were counted using fluorescence microscopy. Recombination efficiency was determined by the ratio of ERα-positive to isolectin B4-positive cells. Overall, 400 cells were evaluated. Microglial cells of ERαfl/fl mice were stained as a control.
For detection of ERα in sections, the brains were fixed in 4% paraformaldehyde for 48 h. After blocking of endogenous peroxidase activity, coronal vibratome sections (40-μm-thick) were incubated overnight with anti-ERα. The primary antibody was detected with biotinylated anti-rabbit antibody, the ABC-peroxidase system (Vector, Burlingame, CA, USA), and 3,3′-diaminobenzidine (Sigma, Munich, Germany) as substrate.
Real-Time Reverse Transcription-PCR
Cortices were dissected and homogenized using an Ultra-Turrax T8 homogenizer (IKA Werke, Staufen, Germany). For RNA isolation, we used the RNeasy Mini Kit (Qiagen, Hilden, Germany). We removed genomic DNA from the lysate by using RNase-free DNase (Qiagen). RNA was transcribed using the SuperScript II Reverse Transcriptase Kit (Invitrogen). For real-time PCR, the Absolute QPCR-mix (Thermo Scientific, Karlsruhe, Germany) and the following primers and probes were used: ERα, Mm 00433149_m1; hypoxanthine guanine phosphoribosyl transferase (HPRT), Mm 00446968_m1 (Applied Biosystems, Darmstadt, Germany). Results were normalized to HPRT expression.
Middle Cerebral Artery Occlusion
Eight-week-old female mice were anesthetized with intraperitoneal injection of 150 μL of 2.5% tribromoethanol per 10 g body weight and ovariectomized. Both female and male mice received a pellet containing 0.025 mg E2 with a release time of 21 days (Innovative Research of America, Sarasota, FL, USA). Control animals received a placebo pellet. Mice were randomly assigned to treatment groups. Ten days after ovariectomy and implantation of pellets, E2 plasma levels were determined by a chemiluminescent immunoassay and the ADVIA centaur (Siemens, Nürnberg, Germany). 17β-Estradiol-treated ovariectomized mice had E2 levels in the physiologic range (ovariectomized, E2-treated, 68.4 ± 13.3 pg/mL, n = 7; female controls, 58.1 ± 4.7 pg/mL, n = 4). After 10 days of recovery from ovariectomy or implantation of the pellet, respectively, mice were anesthetized with intraperitoneal injection of 150 μL of 2.5% tribromoethanol per 10 g body weight. Middle cerebral artery occlusion was performed in mice as described previously (Herrmann et al, 2005; Zhang et al, 2005). Briefly, a skin incision was made between the ear and the orbit on the left side. The temporal muscle was removed by electrical coagulation. The stem of the MCA was exposed through a burr hole and was occluded by microbipolar coagulation. Mice were kept at a body temperature of 37°C on a heating pad. The body temperature was monitored continuously during the surgery using a rectal thermometer. To determine the infarct volume, mice were killed 48 h after the MCAO.
Coronal serial frozen sections of the forebrain were prepared. Silver staining was performed to determine the infarct volume as described previously (Schneider et al, 1999). Stained sections were scanned at 300 d.p.i., and the infarct area was measured using the Scion ImageJ software (Scion, Frederick, MD, USA). To correct for edema formation, the difference between areas of the left and right hemispheres was subtracted from the measured silver-negative area (Swanson et al, 1990). For calculating the whole brain infarct volume, the infarct areas were added and multiplied by the distance between the sections (0.4 mm). Surgery was performed, and ischemic damage was measured by an investigator who had no knowledge of the treatment group or the genotype. Animal experiments were approved by the Regierungspräsidium Karlsruhe.
Measurement of Physiologic Parameters
Measurement of arterial blood pressure, pulse, and blood gases was carried out both before and after ischemia in a separate cohort of mice. Mice were kept under tribromoethanol anesthesia on a heating pad at 37°C, and the body temperature was measured using a rectal thermometer. For measuring blood pressure and pulse, a cannula was inserted into the right femoral artery. A blood sample of 150 μL per mouse was collected in a heparin-coated glass capillary for analysis of arterial blood gas, hemoglobin, and glucose levels. The catheter was washed with 200 μL NaCl solution mixed with 50 IE (international units) of heparin before measuring blood pressure. For laser Doppler measurements, the electrode (P415-205; Perimed, Jarfalla, Sweden) was placed 3 mm lateral and 6 mm posterior to the bregma. Relative perfusion units were determined (Periflux 4001; Perimed).
Statistical Analysis
Student's t-test was used to compare two groups and one-way ANOVA (analysis of variance) for comparing more than two groups. Data are expressed as means ± s.d.
Results
17β-Estradiol treatment is neuroprotective in various rodent models of stroke (Gibson et al, 2006). To verify the efficacy of E2 treatment in a model of permanent distal MCAO using C57BL/6 mice, we implanted ovariectomized female mice with E2 or placebo pellets. Ten days later, mice were subjected to MCAO and killed after 48 h to determine the infarct volume. 17β-Estradiol treatment significantly reduced the infarct volume (Figure 1).
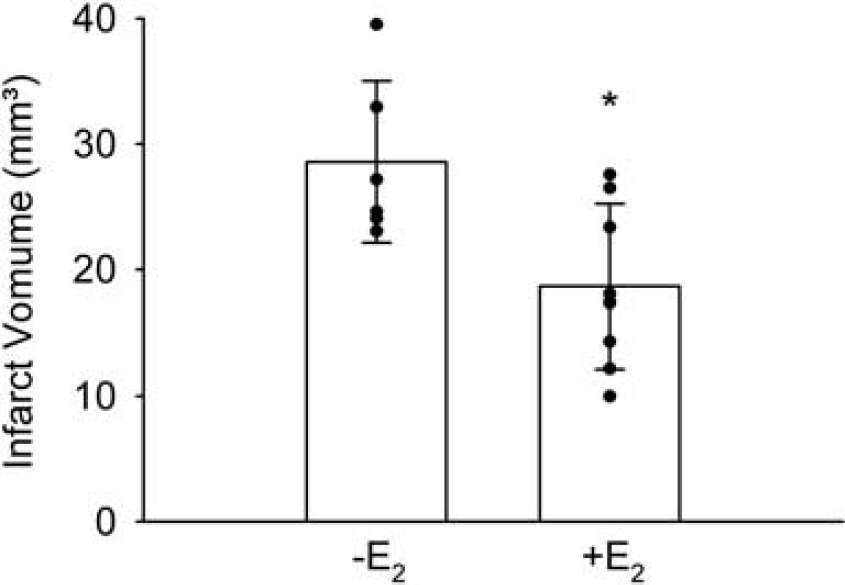
17β-Estradiol (E2) reduced the infarct volume in ovariectomized mice that were subjected to permanent distal MCAO. The infarct volume was determined 48 h after MCAO. Means ± s.d. and individual values are depicted (n = 6 to 8). *P < 0.05 (t-test).
To investigate the role of ERα in myeloid cells including microglia, we generated myeloid lineage-specific ERα mutant mice by breeding mice expressing the Cre recombinase (Cre) under control of the lysozyme M promoter (Clausen et al, 1999) with mice harboring a conditional ERα allele (LysCre/ERfl/fl) (Wintermantel et al, 2006). To investigate the recombination efficiency, we detected nuclear ERα expression in isolated microglial cells by immunocytochemistry. All the microglia obtained from control mice were stained (Figures 2A and 2C), whereas 92% of the microglial cells isolated from LysCre/ERfl/fl mice showed no immunoreactivity for ERα (Figures 2B and 2C). 17β-Estradiol treatment reduced the infarct volume of control mice (ERfl/fl). After E2 treatments, the infarct size of LysCre/ERfl/fl mice and control mice did not differ (Figure 3), indicating that the absence of microglial ERα does not affect ischemic brain damage in E2-treated mice.
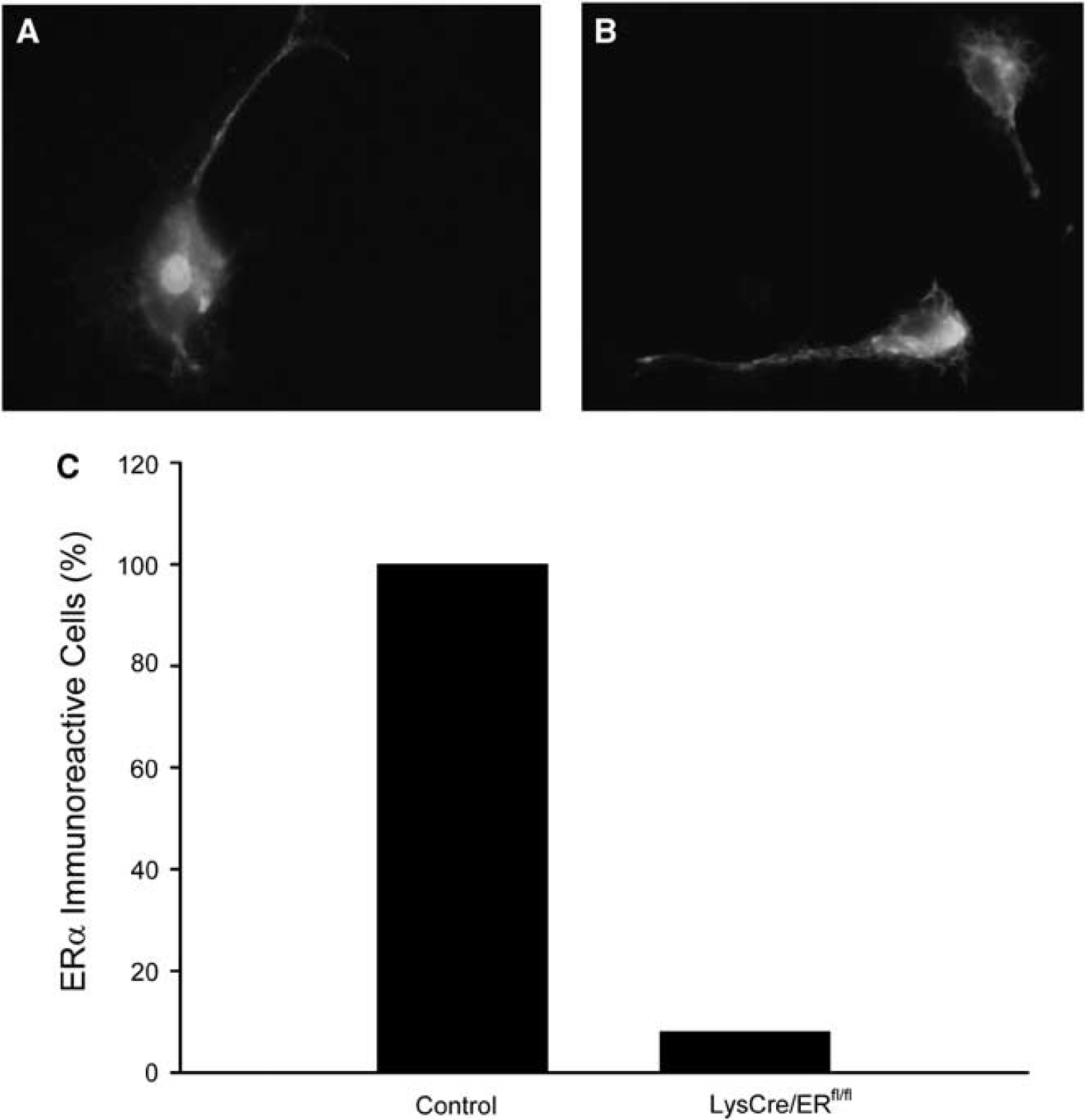
Deletion of ERα in microglial cells of LysCre/ERfl/fl mice. (
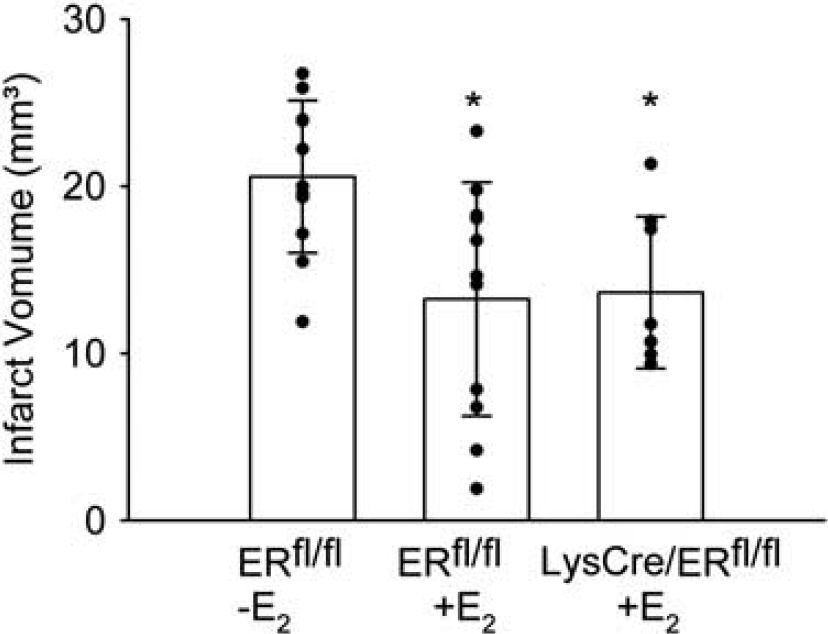
Deletion of ERα in microglial cells did not interfere with the neuroprotective effect of 17β-estradiol (E2). The infarct volume of E2-treated female LysCre/ERfl/fl (n = 8) and control ERfl/fl mice (n = 11) did not differ. In both groups, the infarct volume was significantly lesser than in female control ERfl/fl mice treated with placebo pellets (n = 11). ANOVA, F(2/27) =5.739, P = 0.008. *P < 0.05 (Student–Newman–Keuls method) compared with ERfl/fl-E2. Means ± s.d. and individual values are depicted.
To analyze the contribution of neuronal ERα, we deleted ERα in the neurons of the forebrain by breeding mice expressing the Cre recombinase under control of the CaMKII promoter with ERfl/fl mice (Wintermantel et al, 2006). As described previously for ERα expression in hypothalamic neurons, we found a marked reduction of ERα immunoreactivity in the cortices of CaMKIICre/ERfl/fl mice (Figure 4A). Quantification of ERα mRNA by real-time reverse transcription PCR 24 h after MCAO confirmed this finding. Estrogen receptor-α mRNA was upregulated in the ischemic ipsilateral cortex compared with the contralateral side (Figure 4B) in accordance with a report that cerebral ischemia induces ERα expression (Dubal et al, 2006). However, in CaMKIICre/ERfl/fl mice, ERα expression was markedly reduced both in the ischemic and the contralateral sides (Figure 4B).
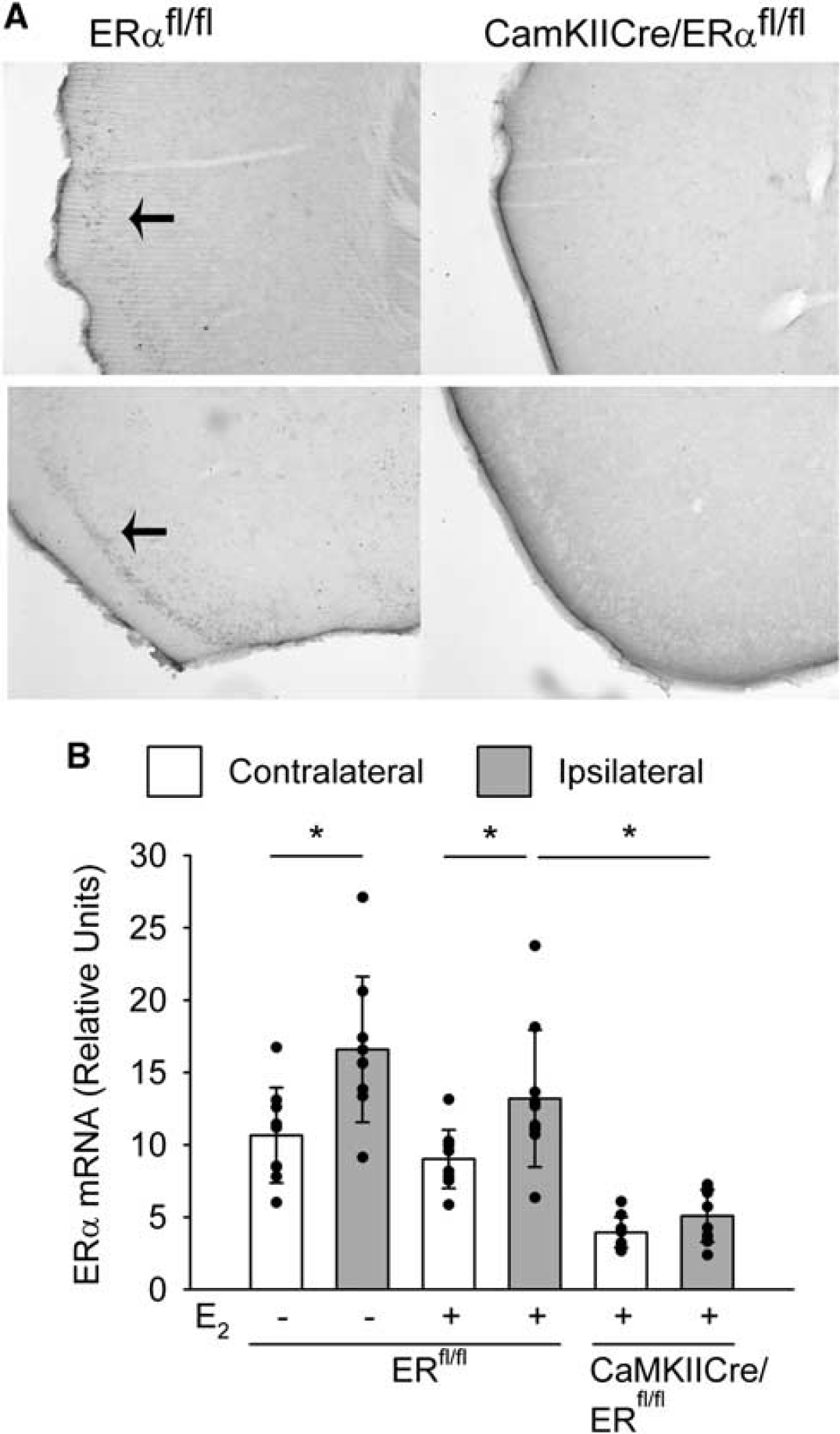
Reduced ERα expression in CaMKIICre/ERfl/fl mice. (
17β-Estradiol-treated female CaMKIICre/ERfl/fl mice showed an increased infarct volume compared with E2-treated control mice 48 h after MCAO (Figures 5A and 5B). Infarct volumes of E2-treated CaMKIICre/ERfl/fl mice were similar to those of the placebo-treated control animals. This finding was reproduced by an independent experiment (infarct volume of ovariectomized ERfl/fl mice without E2 treatment, 21.0 ± 3.6 mm3, n = 7; ovariectomized ERfl/fl mice with E2 treatment, 16.0 ± 5.6 mm3, n = 8; ovariectomized CaMKII/ERfl/fl with E2 treatment, 24.5 ± 6.3 mm3 *, n = 7; ANOVA, F(2/19)=4.833, P = 0.020; *P < 0.05 compared with ERfl/fl mice with E2 treatment, Student–Newman–Keuls method). We conclude that the neuroprotective effect of E2 is absent in female mice lacking ERα specifically and only in neurons.

Deletion of ERα in neurons abolished the neuroprotective effect of 17β-estradiol (E2) in female mice. (
To investigate whether this mechanism is sex-specific, E2-treated male CaMKIICre/ERfl/fl and control mice were also investigated in the MCAO model. Similar to female mice, E2-treated male CaMKIICre/ERfl/fl mice showed an increased infarct volume compared with E2-treated control mice (Figure 6), suggesting that neuroprotection by E2 depends on neuronal ERα.

Deletion of ERα in neurons abolished the neuroprotective effect of 17β-estradiol (E2) in male mice. The infarct volumes of E2-treated male CaMKIICre/ERfl/fl mice (n = 11) were larger than those of E2-treated male control ERfl/fl (n = 8) mice. Placebo-treated control mice, n = 10. ANOVA, F(2/26) = 5.697, P < 0.01. *P < 0.01 (Student–Newman–Keuls method).
To exclude differences in cardiovascular parameters between control and CaMKII/ERfl/fl mice responsible for the phenotype, we measured blood pressure, pulse, blood gases, and glucose levels before and after MCAO (Table 1). No significant differences were detectable between genotypes.
Preischemic and postischemic physiologic parameters of female ERαfl/fl and CaMKIICre/ERfl/fl mice, both treated with E2

b.p.m. beats per minute; ERα, estrogen receptor-α; MCAO, middle cerebral artery occlusion.
There were no significant differences in physiologic parameters between genotypes detectable (t-test).
Discussion
The neuroprotective effect of estrogens in physiologic doses is well established (Gibson et al, 2006; Strom et al, 2009), but the molecular and cellular mechanisms remain controversial. Although receptor-independent effects of E2 have been reported (Culmsee et al, 1999), there is ample evidence that suggests that ERs mediate most of the neuroprotective effect. In line with this concept, the ER antagonist ICI182780 increased the infarct volume after MCAO (Sawada et al, 2000). Selective ER agonists have been used to define which of the two ERs reduces ischemic brain damage. These studies suggested that in models of global ischemia, ERβ is involved in neuroprotection (Carswell et al, 2004; Miller et al, 2005), but in focal cerebral ischemia only an ERα agonist was neuroprotective (Farr et al, 2007). However, a neuroprotective function of ERα has been challenged by the finding that after transient MCAO, the infarct size did not differ in control and ERα knockout mice (Sampei et al, 2000). In this study, mice were not ovariectomized. As ERα knockout mice have highly elevated serum levels of E2 (Couse et al, 1995), the effect of ERα may have been compensated by an ERα-independent effect of high E2 levels. Indeed, deficiency of ERα but not of ERβ abolished the protective effect of E2 in ovariectomized mice subjected to focal cerebral ischemia (Dubal et al, 2001). Our data in CaMKIICre/ERfl/fl mice support the neuroprotective role of ERα.
Estrogen receptor-α is expressed in many cells of the brain, including neurons, astrocytes, microglia, and endothelial cells. Therefore, the localization of ERα involved in the protection against ischemic damage is not self-evident. Numerous in vitro studies have approached the cellular basis of E2 neuroprotection presenting evidence for the involvement of neurons (McEwen and Alves, 1999), astrocytes (Dhandapani and Brann, 2007), microglia (Bruce-Keller et al, 2000; Ghisletti et al, 2005), or endothelial cells (Razmara et al, 2008). In vivo E2 inhibits microglia activation through ERα (Vegeto et al, 2003) and induces the expression of bfl-1, an antiapoptotic member of the bcl-2 gene family, in the microglia (Zhang et al, 2004). Whether these effects cause neuroprotection is unclear. To identify the cell type in which ERα exerts its neuroprotective action, we deleted ERα in neurons or microglia and performed MCAO in these mice.
The results show that ERα in the microglia and possibly in other cells of myeloid lineage does not mediate the effect of E2 on infarct volume. We investigated the infarct volume 48 h after MCAO, when the microglia was activated in the MCAO model we used (Muhammad et al, 2008). However, we cannot exclude the fact that at other time points ERα deletion in myeloid cells may affect the infarct size.
In contrast to LysCre/ERfl/fl mice, neuronal ERα knockout animals had larger infarcts than did controls after E2 treatment. In fact, the infarct size of E2-treated CaMKIICre/ERfl/fl mice was identical to that of untreated controls. We cannot exclude that ERα exerts an E2-independent effect on the infarct size. It is also conceivable that androgen levels were altered in male CaMKIICre/ERfl/fl mice, as reported in ERα knockout mice (McDevitt et al, 2007), and that androgens may have affected ischemic brain damage, because male mice were not castrated in our experiments. However, the most likely explanation of our findings is that neuroprotection by E2 is mediated by neuronal ERα. The introduction of the CaMKIICre gene to delete ERα from neurons is a possible confounding factor of our results. As previous experiments found no effect of other Cre genes on ischemic brain damage (Inta et al, 2006; data not shown), it seems very unlikely that the CaMKIICre gene itself could lead to enlarged infarct volumes.
This study suggests neuronal ERα as one of the major targets of E2-mediated neuroprotection in a model of cerebral stroke. To our knowledge, this is the first time that the neuroprotective actions of ERα in stroke were investigated on a cellular level in vivo. It should be noted that cells of the vessel wall, glia, and hematopoietic cells retain ERα in the CaMKIICre/ERfl/fl mouse line, but the E2 action on these targets apparently does not influence the infarct size in this model of acute stroke. Clearly, potential beneficial vascular actions of ERα in stroke prevention and recovery are not addressed in this stroke model.
In view of these in vivo findings, previous studies analyzing the neuroprotective effects of estrogens in cultured neurons gain importance. Intracellular signaling pathways leading to neuroprotection involve PI3K, AKT, and MAP kinases (Morrison et al, 2006). In addition, ERs are known to inhibit the transcription factor nuclear factor-κB (Biswas et al, 2005), an effect that has also been reported in a model of cerebral ischemia in vivo (Wen et al, 2004). Inhibition of nuclear factor-κB in neurons is protective in stroke models (Herrmann et al, 2005; Zhang et al, 2005), suggesting that pathway-selective ER ligands that inhibit nuclear factor-κB (Chadwick et al, 2005) might be an option for stroke treatment. Another way to refine estrogen treatment for stroke therapy would be the development of neuron-specific selective ER modulators (neuro-SERMs) (Zhao et al, 2005), a strategy that would exploit the important role of neuronal ERα shown in this study.
Footnotes
TMW is an employee of Bayer Schering Pharma AG.
Acknowledgements
We are grateful to R Geibig and M Westphal for technical support.