
Abstract
The role of the β2AR (β2 adrenergic receptor) after stroke is unclear as pharmacological manipulations of the β2AR have produced contradictory results. We previously showed that mice deficient in the β2AR (β2KO) had smaller infarcts compared with WT (wild-type) mice (FVB) after MCAO (middle cerebral artery occlusion), a model of stroke. To elucidate mechanisms of this neuroprotection, we evaluated changes in gene expression using microarrays comparing differences before and after MCAO, and differences between genotypes. Genes associated with inflammation and cell deaths were enriched after MCAO in both genotypes, and we identified several genes not previously shown to increase following ischaemia (Ccl9, Gem and Prg4). In addition to networks that were similar between genotypes, one network with a central core of GPCR (G-protein-coupled receptor) and including biological functions such as carbohydrate metabolism, small molecule biochemistry and inflammation was identified in FVB mice but not in β2KO mice. Analysis of differences between genotypes revealed 11 genes differentially expressed by genotype both before and after ischaemia. We demonstrate greater Glo1 protein levels and lower Pmaip/Noxa mRNA levels in β2KO mice in both sham and MCAO conditions. As both genes are implicated in NF-κB (nuclear factor κB) signalling, we measured p65 activity and TNFα (tumour necrosis factor α) levels 24 h after MCAO. MCAO-induced p65 activation and post-ischaemic TNFα production were both greater in FVB compared with β2KO mice. These results suggest that loss of β2AR signaling results in a neuroprotective phenotype in part due to decreased NF-κB signalling, decreased inflammation and decreased apoptotic signalling in the brain.
INTRODUCTION
Stroke is the fourth leading cause of death and the primary cause of chronic neurological disability in the United States (http://www.strokeassociation.org). The β2AR (β2 adrenergic receptor) is a member of the adrenergic receptor family, a group of GPCR (G-protein-coupled receptors) that respond to endogenous and exogenous catecholamines, primarily noradrenaline (Wallukat, 2002). Previous research examining the role of β2AR activation after stroke using pharmacological agents has produced contradictory results. Although intraperitoneal pre-treatment with clenbuterol, a β2AR agonist, significantly decreased infarct size after permanent MCAO (middle cerebral artery occlusion) (Semkova et al., 1996), intravenous pre-treatment with four individual non-specific βAR blockers resulted in reduced infarct size after transient MCAO (Goyagi et al., 2006). In an effort to better understand how activation of the β2AR affects stroke outcome, our laboratory used transgenic mice lacking a functional β2AR in all cell types throughout the body (Chruscinski et al., 1999), and showed that these mice had smaller ischaemic infarcts and improved locomotor recovery following transient MCAO, a model of stroke (Han et al., 2009).
The β2AR is widely expressed throughout the brain and the rest of the body. In the brain, the β2AR is found on neurons (Vasquez and Lewis, 2003; Ampatzis and Dermon, 2010), astrocytes (Hutchinson et al., 2007; Dong et al., 2012) and microglia (Fujita et al., 1998; Tanaka et al., 2002). β2AR dysfunction has been implicated in multiple sclerosis (De Keyser et al., 1999) and Alzheimer's disease (Wang et al., 2011), suggesting that it may play a direct role in regulation of neuronal death. In the periphery, the β2AR is expressed by Band T-lymphocytes (Kin and Sanders, 2006; Sanders, 2012), in addition to cells in most other organs (Anderson, 2006; Snyder et al., 2008). Treatment with β2AR agonists for asthma and COPD (chronic obstructive pulmonary disease) induces brochodilation, although efficacy is reduced over time (Anderson, 2006). An interaction with glucocorticoids has been noted with possible effects on inflammation (Eickelberg et al., 1999).
As the CNS (central nervous system) and peripheral immune system communicate via the release of the adrenergic agonist noradrenaline by the sympathetic nervous system (Sanders, 2012), deletion of the β2AR would be expected to cause changes in immune response both systemically and in the brain. Predicting likely effects is difficult since, depending on cell type, stimulus and time course, the β2AR can either have pro-inflammatory effects (Christensen et al., 1999; Frost et al., 2004; Yin et al., 2006; Rohrbach et al., 2007; Tan et al., 2007) or anti-inflammatory effects (Severn et al., 1992; Mori et al., 2002). To explore how the absence of β2AR receptors is neuroprotective we conducted microarray analysis of brain gene expression in WT (wild-type) (FVB) and β2AR-deficient (β2KO) mice 24 h after stroke. By examining gene expression differences between genotypes, we show that β2KO mice have increased expression of Glo1 and decreased expression of Pmaip1/Noxa, contributing to decreased NF-κB (nuclear factor κB) signalling, TNFα (tumour necrosis factor a) production and apoptosis following stroke. Together, these data show that absence of β2ARs appears to have anti-inflammatory and anti-apoptotic effects in the brain that could contribute to protection following stroke.
MATERIALS AND METHODS
Mice
Male β2AR-deficient FVB/N (β2KO) and congenic WT FVB/N (FVB) mice weighing 25–30 g were used. The β2KO mice are those produced by targeted gene disruption (Chruscinski et al., 1999). All experiments were performed according to a protocol approved by the Stanford Institutional Animal Care and Use Committee.
Transient focal ischaemia
Transient focal ischaemia was induced with an intraluminal suture to occlude the middle cerebral artery, as previously described (Han et al., 2009) with some modifications. Briefly, mice were anaesthetized with isofluorane in 70% nitrous oxide balance oxygen with spontaneous respiration. Under the operating microscope, the left common carotid artery, internal carotid artery and external carotid artery were exposed through a midline neck incision. The proximal portions of the left common and external carotid arteries were ligated and a 6-0 silicon-coated nylon suture was introduced into the distal common carotid and advanced approximately 9.0 mm beyond the carotid bifurcation to transiently occlude the middle cerebral artery for 60 min. During all surgical procedures mice were maintained normothermic (37 °C±1.0) using a servo controlled heating blanket (Harvard Apparatus) with rectal temperature monitoring. Animals with no observable deficits immediately after ischaemia, those that died in the first 24 h of reperfusion, and those with subarachnoid haemorrhage at time of killing were excluded from analysis. Sham-treated animals received all surgical procedures but the filament was not inserted and the artery was not opened.
Preparation of samples for microarray analysis
At 24 h after MCAO, sham surgery, or no manipulation (naïve) mice were killed under deep isofluorane anaesthesia, transcardially perfused with cold saline, and cerebral cortices were immediately extracted. RNA was extracted from the ischaemic or ipsilateral sham hemisphere using the RNeasy Kit (Qiagen) according to the manufacturer's instructions. Total RNA samples were processed at the Stanford Protein and Nucleic Acid Biotechnology Facility by one-cycle target preparation, labelling and hybridization to Affymetrix 430_2® (Affymetrix) whole genome mouse arrays, according to the manufacturer's protocol. Naïve, sham and MCAO groups of each genotype were analysed with 7–8, 3 and 7 biological replicates/group respectively. Each array analysed RNA from an individual mouse.
SAM (significance analysis of microarrays) analysis
The SAM method was used (Tusher et al., 2001) to analyse the array data. Raw image files were processed using Affymetrix GCOS software. Initial chip processing and signal calling were done with R and Bioconductor (Gentleman et al., 2004) using the Affymetrix package (Gautier et al., 2004) with RMA background correction, quantile normalization, and the median polish probe set summary method. A fold-change of 2.0 and false positive rate (q-value) of 0.05 were chosen as the cut-offs for selecting significantly differentially expressed genes. A current gene assignment for the Affymetrix probe sets was done using the Gene ID conversion tool on the DAVID bioinformatic resource (http://david.abcc.ncifcrf.gov/) (Dennis et al., 2003). Datasets are posted on the Gene Expression Omnibus (GSE30655).
SAGAT (singular value decomposition augmented gene expression analysis tool) analysis
To analyse the microarray data, we also used the recently developed SAGAT (Daigle et al., 2010). SAGAT extracts transcriptional modules from publicly available microarray data and integrates this information with the dataset of interest. In so doing, it boosts experimental power by increasing the effective sample size of the dataset. SAGAT has been shown to capture additional genes and pathways that may be missed when using methods that ignore data from pre-existing experiments (Daigle et al., 2010).
All available expression data for the Affymetrix Mouse Genome 430 2.0 Array (GPL12610) from the Gene Expression Omnibus were downloaded in March 2009. Probes were mapped to a non-redundant list of Entrez Gene IDs (provided by the Bioconductor R package, mouse4302) and expression values for multiple probes of the same gene were averaged. This resulted in a matrix of 21308 genes by 9199 arrays. We excluded 1168 arrays on the basis of missing data. We then ran the R svd function to generate the weight matrix for SAGAT.
Using the R affy and bias packages, we performed background correction, RMA normalization and bias correction on CEL files from the experiments in this study. We used SAGAT to identify genes that were differentially expressed when comparing MCAO with sham. Genes with a false discovery rate less than or equal to 0.05 were considered statistically significant. A list of the top 500 genes, based on SAGAT scores, a measure of statistical significance, was produced.
RT–qPCR (reverse transcription–quantitative real-time-PCR) for mRNA quantification
Total RNA was isolated with TRIzol® (Invitrogen) from the ischaemic hemisphere of WT or β2KO mice 24 h following MCAO. RT was performed using the TaqMan MicroRNA Reverse Transcription Kit (Applied Biosystems). Equal amounts of RNA (600 ng) were reverse-transcribed with 1.3 mM dNTPs, 50 units of reverse transcriptase, 10 units of RNase inhibitor and random primers at 25 °C for 10 min, 37 °C for 120 min and 85 °C for 5 min. PCRs were then conducted using the TaqMan® MicroRNA Assay Kit (Applied Biosystems) at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °Cfor 1 min. Each reaction contained 0.75 μl of the reaction product and 5 μl of 2 × Taqman Universal Master Mix (Applied Biosystems) in a total volume of 10 μl using the 7900 HT (Applied Biosystems). Predesigned primer/probes (Applied Biosystems) for mRNAs and GAPDH (glyceraldehyde-3-phosphate dehydrogenase) were also from Applied Biosystems. The expression of mRNAs was normalized using GAPDH as the internal control. Measurements were normalized to GAPDH (ΔC t) and the comparison calculated as the inverse log of ΔΔC t to give a relative fold change value (n=3 per sham group and 4–5 per MCAO group).
Biological function and network analysis
Annotation for biological functions, similar to gene ontology categories, was completed using the IPA (ingenuity pathway analysis) program (Ingenuity Systems, http://www.ingenuity.com). Genes that were differentially expressed, defined as at least a 2-fold change between sham and ischaemic conditions with P<0.05, were used for the analysis. IPA was performed to identify the top biological functions and networks enriched in each gene set. To do this, a dataset containing gene identifiers and corresponding expression values was uploaded into the application. The Biological Function Analysis identified the functions in Ingenuity's Knowledge Base that were most significant in the dataset. Right-tailed Fisher's exact test was used to calculate a P-value determining the probability that each biological function assigned to that dataset is due to chance alone. For the Network Analysis, each differentially expressed gene (identifier) was mapped to its corresponding object in Ingenuity's Knowledge Base. These genes, called Network Eligible molecules, were overlaid on to a global molecular network developed from information contained in Ingenuity's Knowledge Base. Networks of Network Eligible Molecules were then algorithmically generated based on their connectivity. In addition, a comparison analysis between networks enriched after ischaemia in FVB and β2KO mice was conducted to assess the number of shared genes between networks.
Protein preparation for ELISA and immunoblotting
At 24 h following MCAO or sham treatment mice were transcardially perfused with cold saline, and the cerebral cortices were dissected and placed on dry ice. Protein lysis buffer (10 mM Hepes, 42 mM KCl, 5 mM MgCl2, 0.1 mM EDTA, 0.1 mM EGTA and 0.1% Triton X-100) with ProteaseARREST (G Biosciences) was added to tissue at a 1:3 ratio and the mixture was sonicated for 5 s. Following sonication and centrifugation at 15000
Immunoblotting
Samples containing equal amounts of protein (100 μg) were resolved by SDS/4–12% PAGE (NuPage Bis/Tris Gel, Invitrogen). The proteins were transferred on to a PVDF membrane (IPVH00010, Millipore), probed with primary antibodies [rat anti-Glo1 (1:2000, Abcam) and mouse anti-β-actin (1:5000, Sigma) or rabbit anti-β-actin (1:000, Licor Systems)] probed with Licor Fluorescent secondary antibodies (goat anti-rat 680LT and goat anti-mouse 800 or goat anti-rabbit 800, all at 1:15000), and scanned using the Licor Odyssey system. Densitometric analysis of the bands was performed using NIH ImageJ software, with band intensities normalized to β actin.
ELISA
Samples containing equal amounts of protein (100 μg) were analysed with the mouse TNFα ELISA kit (Invitrogen). As naïve brains contain very low levels of pro-inflammatory cytokines (Offner et al., 2006), only post-MCAO tissue was analysed. All samples were run in duplicate, and data were fitted to a standard curve from standards provided by the manufacturer.
p65 activity assay
At 24 h following MCAO or sham treatment mice were transcardially perfused with cold saline, and the cerebral cortices were dissected and placed in liquid nitrogen. Complete lysis buffer AM2 from the TransAM NF-κB p65 Activity Assay (Active Motif) was added to tissue at a 1:5 ratio and the mixture was sonicated for 5 s. In total 20 μl of the mixture was added to each well and measured according to manufacturer's instructions. All samples were run in duplicate. Data are presented as the fold change in absorbance value from sham to MCAO in each genotype.
Statistics
For biochemical experiments data are presented as means±S.E.M. ELISA, immunoblotting, RT–qPCR and p65 activity data were analysed using Student's t tests. Statistical significance was set at P<0.05.
RESULTS
Focal ischaemia alters gene expression in both FVB and β2KO mice, sham versus MCAO
The Affymetrix Mouse 430A 2.0 array assesses 45037 probes for approximately 14000 genes (1–4 probes per gene comprising a probeset). To compare gene expression levels, the median of each probeset was determined to produce a single value for each gene. Genes were considered differentially regulated if the fold change between sham and ischaemia was ≥2 with a P-value of <0.05. Using these criteria, SAM analysis yielded a total of 414 genes differentially regulated after MCAO in the FVB group and 347 genes differentially regulated after MCAO in the β2KO group (Figures 1A and 1B, Supplementary Tables S1 and S2 at http://www.asnneuro.org/an/004/an004e096add.htm). Of these, 260 were shared between genotypes (243 up, 17 down), which was 80% of the FVB genes and 75% of the β2KO genes. In the FVB group, 305 genes were up-regulated after ischaemia (74%) and 109 were down-regulated (26%). In the β2KO group, 324 were up-regulated (93%) and 23 were down-regulated (7%).

(
The SAGAT analysis generates a list of genes ranked by SAGAT score, a value derived both from information from the current arrays and publicly available mouse microarray data (Daigle et al., 2010). To match the number of differentially regulated genes identified by the SAM analysis, the top 414 genes ranked by SAGAT score in the FVB mice and the top 347 genes in the β2KO mice were analysed. This revealed that 234 up-regulated genes and 32 down-regulated genes were shared between genotypes, 67% of FVB and 88% of β2KO up-regulated genes and 48% of FVB and 40% of β2KO down-regulated genes (Figure 1C and Supplementary Tables S3 and S4 at http://www.asnneuro.org/an/004/an004e096add.htm). In the FVB group, 347 genes were up-regulated after ischaemia (84%) and 67 were down-regulated (26%). In the β2KO group, 266 were up-regulated (77%) and 81 were down-regulated (23%). We compared the genes identified by the SAM and SAGAT analyses, and found for FVB mice, 251 up-regulated and 51 down-regulated genes were shared between analyses (Figure 1D). For β2KO mice, 208 up-regulated and 13 down-regulated genes were shared between analyses (Figure 1E). The top 10 genes up-regulated and down-regulated in each genotype from the SAGAT and SAM analyses are shown in Table 1.
The top 10 genes down- and up-regulated after ischaemia in each genotype using SAM and SAGAT analysis
Genes in normal text are in both the SAGAT- and SAM-generated lists. Those in bold do not appear on the other list using the other analysis.

Prg4, Ccl9 and Gem are novel genes up-regulated following MCAO in both genotypes
A list of the top 20 genes up-regulated by MCAO (compared with sham) by SAM analysis is given in Table 2. Genes previously implicated in ischaemia are noted. We identified several genes not previously described as changing after MCAO, two in the top 20 genes (Prg4 and Ccl9), one differentially expressed at a lower fold-change (Gem), and two previously identified by microarray but not previously validated by RT–PCR (Ch25h and Ifi202b). We confirmed the changes in expression of these nine genes by RT–qPCR from sham to MCAO for both genotypes (Table 3).
The top 20 genes up-regulated after ischaemia and their documentation in previous literature
Each of the top 20 genes up-regulated after ischaemia (by SAGAT analysis) is listed by gene symbol and gene name. FC, fold change from sham to ischaemia. Time up, time at which gene or protein is increased after ischaemia. Injury/species, the ischaemia type and animal species. Method of detection, the way in which gene expression change was assessed. MCAO, middle cerebral artery occlusion; HI, hypoxia/ischaemia; RT–PCR, reverse transcription–PCR; IHC, immunohistochemistry; ISH, in situ hybridization. Genes in bold have previously been implicated using protein analysis, genes in normal type have previously been implicated using RNA analysis, and those in italics have not been previously implicated in ischaemia.

Genes increased 24 h after ischaemia by RT–qPCR
Values reflect fold change from sham to ischaemia±S.E.M. (n=6/group in sham and 10–12/group in MCAO). Genes in bold have not previously been implicated in brain ischaemia.

Genes related to inflammatory response and cell death are enriched in both genotypes following MCAO
Using IPA, we assessed the top biological functions enriched after ischaemia using gene lists generated from the SAM analysis, for genes increased ≥2-fold (Figures 2A and 2B). Overall these were similar for the two genotypes, differing by only 1 of the 6 top biological functions.

(
A network composed largely of GPCR is differentially regulated after MCAO in FVB but not β2KO mice
IPA Network Analysis groups genes based on relationships previously identified in the literature. For example, genes differentially regulated after ischaemia and that were previously shown to associate with each other will be identified as members of a network. If the network members are over represented in our differentially expressed gene list compared with the fraction of the genome represented by all members of the network, then the network is ranked as over-represented. To determine how these networks were different between genotypes, we compared the top 10 networks enriched after ischaemia in each genotype. The analysis examines the number of shared genes between each network and forms a graphical representation linking networks containing shared genes (Figure 3A). The biological functions associated with each ranked network are listed in Table 4. Notably, network 1 of FVB does not share any members with the other identified networks. This network is shown in Figure 3(B). This network included both up- and down-regulated genes with the GPCR family as the core of the network (Figure 3B).
Biological functions associated with networks enriched after ischaemia in FVB and β2KO mice
For each network, the biological functions associated with genes in the network and the number of genes from the microarray that appear in the network are shown.


(
Multiple genes differ between β2KO and FVB mice in naïve, sham and MCAO conditions
To directly analyse genotype effects, SAM analysis was performed to compare differences in gene expression between genotypes by treatment group (naïve, sham and MCAO). Venn diagrams illustrate the number of genes differentially regulated between genotypes in the treatment conditions and how those genes are similar or different (Figures 4A and 4B). With a 2-fold change and P<0.05 cut-off, there were five common genes up-regulated in all conditions in β2KO mice compared with FVB (Figures 4A–4C) and six common genes down-regulated in all conditions in β2KO compared with FVB (Figures 4B and 4C).

(
The genes up-regulated in β2KO mice were: Adrb2, Fxyd2, Glo1, Nedd4L and Vps52. Genes down-regulated in the β2KO mice were: AI506816, Magi2, Pmaip1, Ppcdc, Tcf4 and Zfp398. Glo1 and Pmaip1/Noxa were chosen for verification because both are known to affect cell survival. Glo1 inhibits NF-κB activation (El-Osta et al., 2008) and is protective after renal ischaemia (Kumagai et al., 2009), while Noxa is pro-apoptotic, and has been previously implicated as up-regulated in brain ischaemia and downstream of NF-κB activation (Inta et al., 2006).
Glo1 protein expression is increased in β2KO mice
Using immunoblotting, the sham or MCAO cerebral hemispheres of FVB and β2KO mice 24 h after surgery were probed with a specific antibody against Glo1 protein. Glo1 protein levels in both sham-operated and MCAO-treated β2KO mice were significantly higher (approximately twice) than in FVB brains (Figure 5).
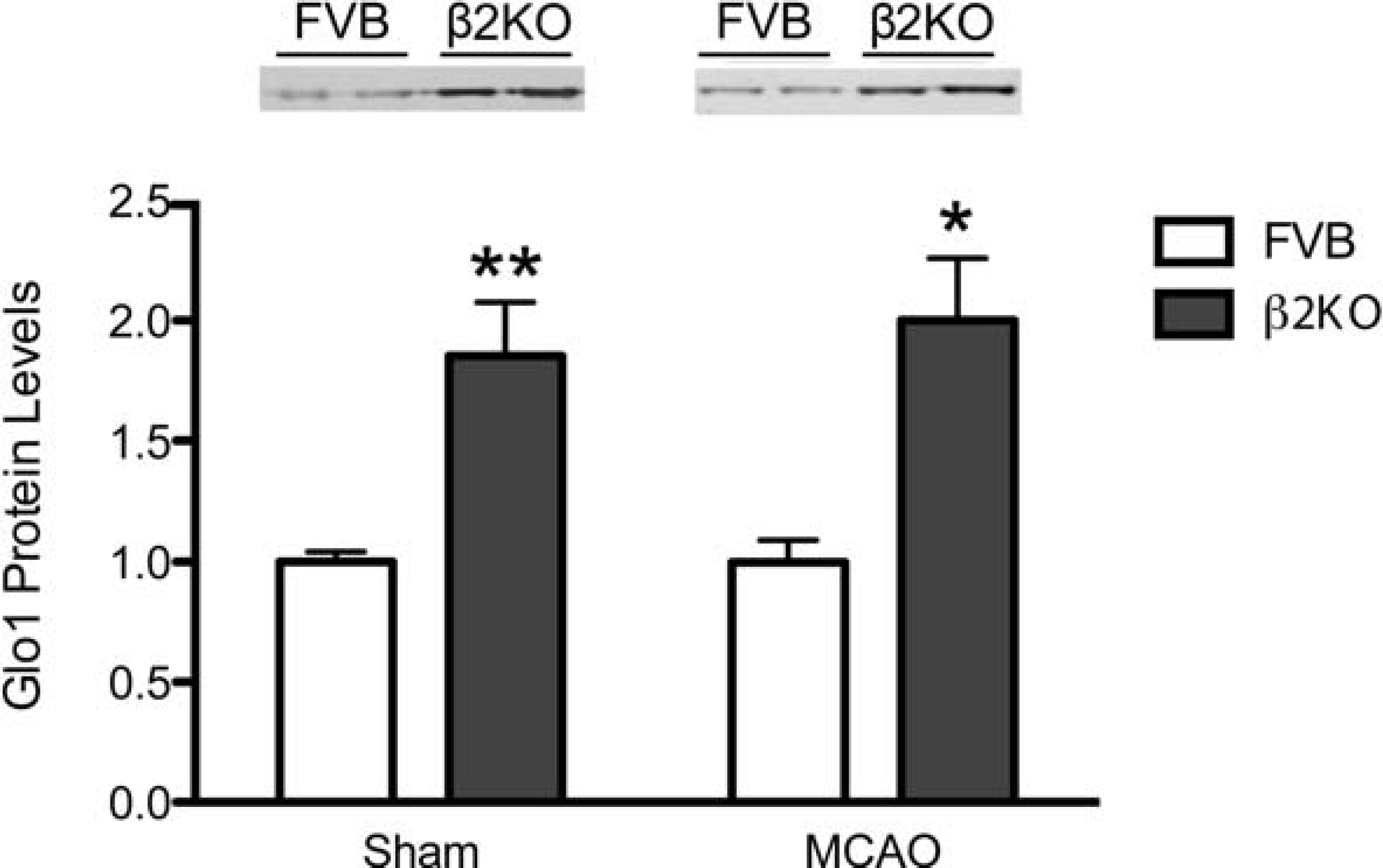
Immunoblots of sham and MCAO tissue probed for an antibody against Glo1. β-Actin protein levels were consistent across samples (data not shown). Graph indicates quantification of blots in sham and MCAO conditions, normalized to β-actin levels (n=3–4/sham group, 4/MCAO group). *P<0.05, **P<0.01.
Noxa mRNA is decreased in β2KO mice
Noxa is decreased 75% in sham and 60% in MCAO conditions in β2KO mice compared with FVB (Figure 6) by RT–qPCR. Previous work found that Noxa was transcriptionally up-regulated by NF-κB activation in the setting of ischaemia (Inta et al., 2006).

Quantification of RT–qPCR showing a significant decrease in Noxa mRNA in sham and MCAO conditions in β2KO mice (n=6/sham group and 10–12/MCAO group). ***P<0.001.
NF-κB activity and TNFα production are attenuated in β2KO mice compared with FVB after MCAO
NF-κB plays a complex role in cerebral ischaemia as a regulator of both apoptotic cell death and inflammation (Ridder and Schwaninger, 2009). In our microarray data, there were no significant differences between genotypes in p65, p50 or Iκbα (inhibitory κB α) and levels of p65 and p50 were not changed by MCAO. Conversely, IκBα was significantly increased from sham to MCAO in both genotypes (3.1-fold in FVB and 2.9-fold in β2KO mice). As NF-κB transcription factor activity is regulated post-translationally, we measured p65 activity in tissue harvested 24 h after sham or MCAO surgery. β2KO mice had a non-significant trend to increased p65 activity in sham compared with FVB (P=0.17, data not shown), while p65 activity increased significantly (20%) more in FVB than in β2KO mice from sham to MCAO (Figure 7A).

(
TNFα is a pro-inflammatory cytokine produced early after ischaemia which induces NF-κB activation, and is one of the inflammatory cytokines downstream of NF-κB activation. In the microarray data, TNFα was 1.4-fold higher in sham β2KO compared with sham FVB mice (P=0.03), with no differences between genotypes after MCAO. Other genes downstream of NF-κB such as IL-1 β (interleukin-1 β) (1.4-fold, P=0.05), IL-6 (1.3-fold, P=0.07) and MMP9 (matrix metalloproteinase 9) (1.4-fold, P=0.05) were all increased in sham β2KO mice compared with sham FVB mice, with no differences between genotypes after MCAO. Increased baseline TNFα, IL-1 β, IL-6 and MMP9 levels in β2KO mice might act as a preconditioning stimulus, limiting NF-κB activation following ischaemia. To assess this we measured TNFα protein levels in the ischaemic brain tissue of the FVB and β2KO mice and found that TNFα protein levels were significantly lower (50%) in β2KO mice compared with FVB mice (Figure 7B).
DISCUSSION
Using microarray analysis, we assessed gene expression changes 24 h after MCAO in FVB and β2KO mice. We used both SAM, a well-established analysis tool, and SAGAT, an alternate strategy that uses pre-existing mouse microarray data to reduce measurement uncertainty. Both methods identified that the top-most up-regulated genes after ischaemia were similar in both groups. This is not surprising, as both genotypes suffered significant injury following MCAO. The SAGAT method is most useful in increasing effective group size for n<4 (Daigle et al., 2010). As we had n=3 in the sham groups but n=7 in our MCAO groups, identification of similar groups of genes by the two methods is expected. The majority of the top up-regulated genes identified in our microarrays have been previously implicated in stroke (Table 2). However, Prg4, Ccl9 and Gem, all of which we confirmed using RT–qPCR, had not been previously shown to increase after MCAO.
Prg4, a proteoglycan found in cartilage, lung, bone and heart (Ikegawa et al., 2000), has never been documented in brain. However, tendon Prg4 mRNA is up-regulated in response to transforming growth factor β (Rees et al., 2002), a factor that increases in the brain following stroke (Dhandapani and Brann, 2003). Although Ccl9 has not been implicated in stroke, brain levels increase in response to infections including pneumococcol meningitis (Klein et al., 2006) and herpes simplex virus (Aravalli et al., 2005). Gem is a guanidine triphosphate binding protein that is induced in response to mitogens (Maguire et al., 1994), suggesting that it may play a role in post-ischaemic cell proliferation. Additional studies will be necessary to determine the roles and potential therapeutic applications of these novel genes. A limitation of our study is that we only performed array analysis and biochemical analyses at 24 h after MCAO, whereas the process of ischaemic brain injury has a much longer course that still needs to be characterized.
To assess differences in gene expression by genotype, we analysed genes up- or down-regulated in β2KO mice compared with FVB in naïve, sham and MCAO conditions. Gene expression differences between genotypes revealed multiple genes altered between β2KO and FVB mice in naïve, sham and MCAO conditions, with 11 genes differentially expressed in all conditions, indicative of changes in response to the chronic absence of the β2 receptor. Perhaps surprisingly, Adrb2, the gene for the β2AR, was significantly increased on the arrays in the β2KO mice compared with FVB. The probeset for Adrb2 targets nucleotides 1608–1835, outside the fourth transmembrane segment sequence targeted and replaced in the KO (Chruscinski et al., 1999). It suggests that the β2KO mice may produce elevated levels of this aberrant RNA perhaps as an attempted compensatory mechanism.
Of the genes differentially expressed between genotypes, two were related to NF-κB activity: Glo1 and Pmaip1/Noxa. This suggested that activation of NF-κB by MCAO might differ between the genotypes, which we confirmed by a p65 activity assay. Furthermore, Glo1 protein levels are increased in β2KO compared with FVB mice. Glo1 detoxifies methylglyoxal, a toxic by-product of glycolysis known as an AGE (advanced glycation end-product) (Thornalley, 1996). AGEs are produced not only by hyperglycaemia but are also produced in the preinfarct area following cerebral ischaemia (Qiu et al., 2008; Harada et al., 2009). Hyperglycaemia-induced NF-κB activation is attenuated by Glo1 overexpression (El-Osta et al., 2008), and Glo1 overexpression is protective in renal ischaemic injury (Kumagai et al., 2009). Thus Glo1 overexpression in the β2KO mice may be protective by attenuating NF-κB signalling and/or decreasing toxic AGEs.
Noxa mRNA levels are decreased in β2KO compared with FVB mice. Noxa, a member of the BH3 (Bcl2 homology domain 3) family of pro-apoptotic proteins, targets the anti-apoptotic Bcl2 molecule Mcl1 for proteasomal degradation, resulting in increased cytochrome c release and caspase activation (Ploner et al., 2008). Noxa antisense significantly decreased infarct size after rat MCAO (Kim et al., 2004), and Noxa was implicated in the neurotoxicity of cerebral ischaemia, where it was shown to be transcriptionally up-regulated by NF-κB activation (Inta et al., 2006; Ridder and Schwaninger, 2009). Together, these results suggest that Noxa directly affects ischaemia-induced cell death, and levels of Noxa are at least partially dependent upon NF-κB activation. This is consistent with our current observation that NF-κB activity and levels of TNFα are both lower in β2KO mice compared with FVB after MCAO, and supports the idea that reduced NF-κB activation leading to lower levels of downstream genes Noxa and TNFα may contribute to protection in the absence of β2AR.
Although NF-κB signalling can protect against oxidative stress-induced neuronal death (Kratsovnik et al., 2005), the majority of the literature suggests that neuronal NF-κB activation (Zheng et al., 2008; Ridder and Schwaninger, 2009) and TNFα are detrimental following stroke. Inhibition of NF-κB activation is protective after rat MCAO (Xu et al., 2002), and transgenic mice specifically overexpressing a dominant non-inactivable IκBα in neurons have significantly smaller infarcts following permanent MCAO (Zhang et al., 2005). Rats overexpressing TNFα have greater injury after stroke (Pettigrew et al., 2008), while TNFα-neutralizing antibodies significantly reduce damage (Yang et al., 1998). Furthermore, activation of NF-κB signalling can lead to further production of harmful pro-inflammatory cytokines (Tak and Firestein, 2001) and chemokines (Thompson and Van Eldik, 2009), contributing to further neuronal death following the initial injury. NF-κB activation has a complicated and variable dynamic timeline, which we have modelled previously (Sheppard et al., 2011). The kinetics of activation is known to influence which downstream targets will be induced (Fine et al., 1999). As this study only assessed NF-κB 24 h following MCAO, additional studies are needed to determine the full kinetics of these processes after stroke.
The reasons for altered NF-κB signalling, cell death and inflammatory response in the β2KO mice could be due to direct, indirect or combined effects of the chronic absence of β2AR signalling. The role of the β2AR in modulating inflammation and NF-κB activation is controversial. Multiple studies have shown that treating cells with selective or non-selective βAR agonists has an anti-inflammatory effect, decreasing TNFα (Severn et al., 1992; Mori et al., 2002), IL-8 (Farmer and Pugin, 2000) and IL-6 (Mohamed-Ali et al., 2000) in response to LPS (lipopolysaccharide). But treatment with agonists alone was also shown to increase IL-6 via NF-κB signalling (Christensen et al., 1999; Frost et al., 2004; Yin et al., 2006; Rohrbach et al., 2007; Tan et al., 2007), and the β2AR-specific agonist terbutaline increased microglial production of harmful ROS (reactive oxygen species) (Qian et al., 2009). Furthermore, treatment with the agonist isoprenaline increased p65 and p50 activity in rat salivary gland 1 h after administration (Yeh et al., 2012).
A previous study (Rough et al., 2009) has begun to shed light on this controversy, demonstrating that pre-treatment with adrenaline for 18 h prior to LPS stimulation significantly increases TNFα production, an effect that is completely abolished by treatment with a β2AR-specific inhibitor. Thus acute and chronic blockade of the β2AR may have different and even opposite effects on NF-κB signalling and inflammation. This is consistent with the known unique switch in coupling of β2AR from Gs to Gi with chronic exposure to agonist (Daaka et al., 1997; Xiao et al., 2006). In our study, chronically absent β2AR signalling contributes to damping production of TNFα and thereby likely attenuating injury. This would be consistent with the idea that chronic absence of β2AR produces a kind of preconditioning. Earlier observations demonstrated that preconditioning is associated with NF-κB activation which then limits NF-κB activation by the more severe ischaemia (Ridder and Schwaninger, 2009).
Preconditioning with low levels of systemic LPS significantly decreases brain inflammation and infarct size following MCAO (Vartanian et al., 2011), while higher doses given before and/or after MCAO significantly exacerbate damage (McColl et al., 2007; Langdon et al., 2010). Thus, chronically mildly elevated levels of systemic inflammation may lead to protection by a form of preconditioning, while high levels of inflammation prior to stroke are likely to be detrimental. Interestingly, our arrays identified a modest increase in TNFα, IL-1β, IL-6 and MMP9 in the sham β2KO compared with FVB mice, suggesting that higher basal brain inflammation might be present in the β2KO mice.
The effects of abolishing β2AR signalling in the periphery as well as in the CNS must be considered, as inflammation in the periphery can affect the CNS and vice versa (Denes et al., 2010). Earlier studies in the β2KO mice found that the contact sensitivity response and T-cell-dependent antibody response in β2KO compared with FVB mice were similar, despite defects demonstrable in isolated immune cells, but the mechanism of compensation was not identified (Sanders et al., 2003). Thus it is unclear whether changes in peripheral inflammation might play a role in the difference between the genotypes.
CONCLUSIONS
Microarray analysis revealed that β2KO mice have a unique genetic profile both before and after ischaemia. The current study demonstrates reduced cell death signalling, reduced inflammation and reduced NF-κB activation in β2KO mice compared with FVB mice. Altered levels of Glo1 and Noxa may contribute to the neuroprotection observed.
Footnotes
Acknowledgements
We thank Rani Agrawal for assistance with RNA isolation and Dr Ludmila Voloboueva, Dr Kurt Lucin and Dr Frances Davies for review of the paper before submission.
Funding
This work was supported, in part, by the