
Abstract
Background:
Jaques Benveniste suggested that electronic transmission of 4-phorbol-12-b-myristate-13-acetate (PMA) activity, carried out using a simple amplifier configured to function as an oscillator when coupled to two electromagnetic coils, demonstrates the same chemical activity as the active molecule. The results obtained suggested that there are associated signals at the PMA molecules that can be transferred to target neutrophils by artificial physical means in a fashion that mimics the original molecules.
Methods:
Retinoic acid was placed at room temperature on one coil attached to an oscillator (VEGA select 719), while LAN-5 neuroblastoma cells were placed on another coil and incubated under controlled condition. The oscillator was then turned on for 12 hours a day for 5 days, after which cells were counted and morphology studied by contrast microscopy.
Results:
The effect of the differentiating agent added to the cell culture by physical means generates a decrease in cell growth, metabolic activity, and the protrusion of a neuritelike structure typical of the differentiated cells.
Conclusions:
These preliminary results suggest that retinoic acid molecules emit signals that can be transferred to LAN-5 neuroblastoma cells by artificial physical means in a manner that seems related to the chemical structure of the source molecules.
Introduction
In the present study, the specific EMS of retinoic acid (RA) (a potent chemical acting on human cells as a differentiating agent) was electronically captured and transmitted to a biologic system constituted by a human neuroblastoma cell line (LAN-5).
LAN-5 neuroblastoma cells represent one of the most common pediatric solid tumors originating from the sympatho-adrenal lineage of the neural crest. This tumor shows extremely different clinical phenotypes such as spontaneous regression on the one hand and aggressive growth on the other hand. Undifferentiated LAN-5 represents a good model to study neuronal differentiation induced by a variety of stimuli such as RA treatment.
Materials and Methods
Cell cultures
LAN-5 cells were grown in RPMI (Gibco Laboratories, Scotland) supplemented with 10% fetal calf serum (Gibco Laboratories, Scotland) and antibiotics (110 IU/mL of penicillin and 0.1 mg/mL of streptomycin) at 37±0.3°C, and 5% CO2 as carbon source and subcultured twice a week at a 1:5 ratio. For every experiment, control and exposed cells were taken from the same flask.
Transmission apparatus
For transmission experiments to cells, the input coil coupled via a homemade amplifier (gain 0.25 dB from 1 to 100-Hz output voltage [max] 20-V p-p, output current 1 A, maximum power 20-W rms) to wave generator VEGA select 719 was operated at room temperature, while the output coil was placed in a cell incubator. The source tube contained 5 μmol/L RA and the target coil contained LAN-5 cells. The target coil was made of 85 turns of 2-mm copper wire, 17-cm long and 9.5-cm wide and fed at 100 mV. Figure 1 is a diagram representing the assembly of the experimental apparatus. The signal from the RA solution in the coil (1) was driven (2A) in the electronic amplifier (2); then from the electronic amplifier (2B) the signal was transferred (3A) in the wave generator (3). In the wave generator, the electronic signal corresponding to RA was superimposed on a 7-Hz sinusoidal frequency carrier modulated at 3 kHz. From the wave generator, the signal (3A) was driven in the cell incubator (4) where the target coil (5) was placed to irradiate LAN-5 cells. The oscillator was then turned on for 12 hours a day for 5 days. During the experimental procedure, the various parameters such as power, voltage, capacitance, and impedance remained constant.

Experimental apparatus and assembly. 1, source signal coil; 2, electronic amplifier; 2A, input signal in the electronic amplifier; 2B, output signal from the electronic amplifier; 3, wave generator; 3A, input signal in wave generator; 3B, output signal from wave generator; 4, cell incubator; 5, target coil.
Cellular metabolic activity and proliferation by water-soluble tetrazolium salt assay
LAN-5 cells were exposed to the electronically transmitted RA EMS by Vega select 719. For each experiment, LAN-5 cells were plated into 25-mL 4.2×5.2-cm base Corning flasks (2.0×105/mL cells in a total volume of 5 mL). The flasks were kept in the exposure system continuously for up to 5 days with or without RA-EMS. Cells were then counted and metabolism determined by the water-soluble tetrazolium salt (WST-1) method. The experiment was repeated 3 times.
The quantification of LAN-5 metabolic activity, as an index of cellular proliferation, was performed by a colorimetric assay based on oxidation of tetrazolium salts (Cell Proliferation Reagent WST-1; Roche Diagnostics, Basel, Switzerland). Cells were cultured for up to 5 days in a normal humidified incubator (control) or in the presence of the RA-EMS (exposed), and they were analyzed by means of a formazan dye every 24 hours. WST reagent diluted to 1:10 was added in the wells at 4 hours and at 1, 2, 3, and 6 days after plating, and then incubated for 2 hours in a humidified atmosphere (37°C, 5% CO2). Quantification of the formazan dye produced was performed by absorbance measurement at 450 nm with a scanning multiwell spectrophotometer (Biotrack II; Amersham Biosciences, Little Chalfont, UK).
Immunofluorescence
For immunofluorescence staining, the cells were grown in Labtek chamber slides. The cells were then washed with phosphate-buffered saline (PBS) with Ca/Mg and fixed with absolute ethanol for 5 minutes, then incubated with the specific monoclonal antibodies, anti-200 kDa neurofilaments (Sigma) appropriately diluted for 1 hour at room temperature. Cells were then washed 3 times with PBS and incubated with fluoresceinated anti-mouse immunoglobulin G F(ab')2 fragment (Sigma), (appropriately diluted) for 1 hour at room temperature.
RT-PCR analysis
Total RNA was extracted from cells using TRIzol Reagent (Life Technologies, Merelbeke, Belgium) according to the manufacturers' instructions. Typically 5–10 μg total RNA per 10 cm2 dish of cell culture was obtained. Reverse transcription–polymerase chain reaction (RT-PCR) was used to evaluate relative mRNA levels of neurofilament protein (NF-200) in control and RA-EMR-exposed LAN-5 cells. One microgram (1 μg) of total RNA was used to synthesize first-strand cDNA with random primers using 100 U of ImProm-II™ RT-PCR kit (Promega, Madison, WI) according to the manufacturer. The reaction was also carried out in the absence of reverse transcriptase (RT) to check for genomic DNA amplification. The NF-200 subunit-specific primers used for PCR were 5'-aagtgaacacagatgctatgcg-3' 5'-ctgtcactccttccgtcacc-3'. The 18S were used as internal controls, because these genes are uniformly expressed during development. The subunit-specific primers used for PCR were 5'-tttcggaactgaggccatgattaag-3' 5'-agtttcagctttgcaaccatactcc-3'. An aliquot (2 μL) of RT reaction was PCR-amplified in a final volume of 50 μL, by using 20 pmol of each primer, 200 μmol/L of each dNTP, and 0.5 U of Taq DNA polymerase (T. Aquaticus, Amersham-Pharmacia). PCR was carried out in a Bio-Rad I Cycle instrument. The thermocycling conditions for each pair of primers were as follows: denaturation at 95°C for 3 minutes followed by 30 cycles of denaturation at 95°C for 30 seconds, annealing for 45 seconds at 62°C, elongation at 72°C for 1 minute, and a final polymerization step at 72°C for 5 minutes for NF-200 and 20 cycles for 18S. The amount of template and the number of amplification cycles were preliminarily optimized for each PCR reaction to avoid conditions of saturation. Aliquots (5 μL) of the reaction products were run on 1% agarose gels containing ethidium bromide (0.5 μg/mL) to mark and visualize the PCR products. Gels were then photographed under ultraviolet light with a Versadoc (Bio-Rad) instrument. These experiments were replicated three different times.
Statistical analysis
Statistics were performed with Student's t test with p<0.05 as the minimum level of significance.
Results
Electronically transmitted RA effect on LAN-5 cell metabolism
The cell growth rate was analyzed by the WST-1 both in LAN-5 cells as control (not exposed electronically transmitted RA) or exposed to the field. An inhibition in the cell metabolism in the electronically transmitted RA exposed was statistically (p<0.01) significant after 5 days' exposure (Fig. 2).
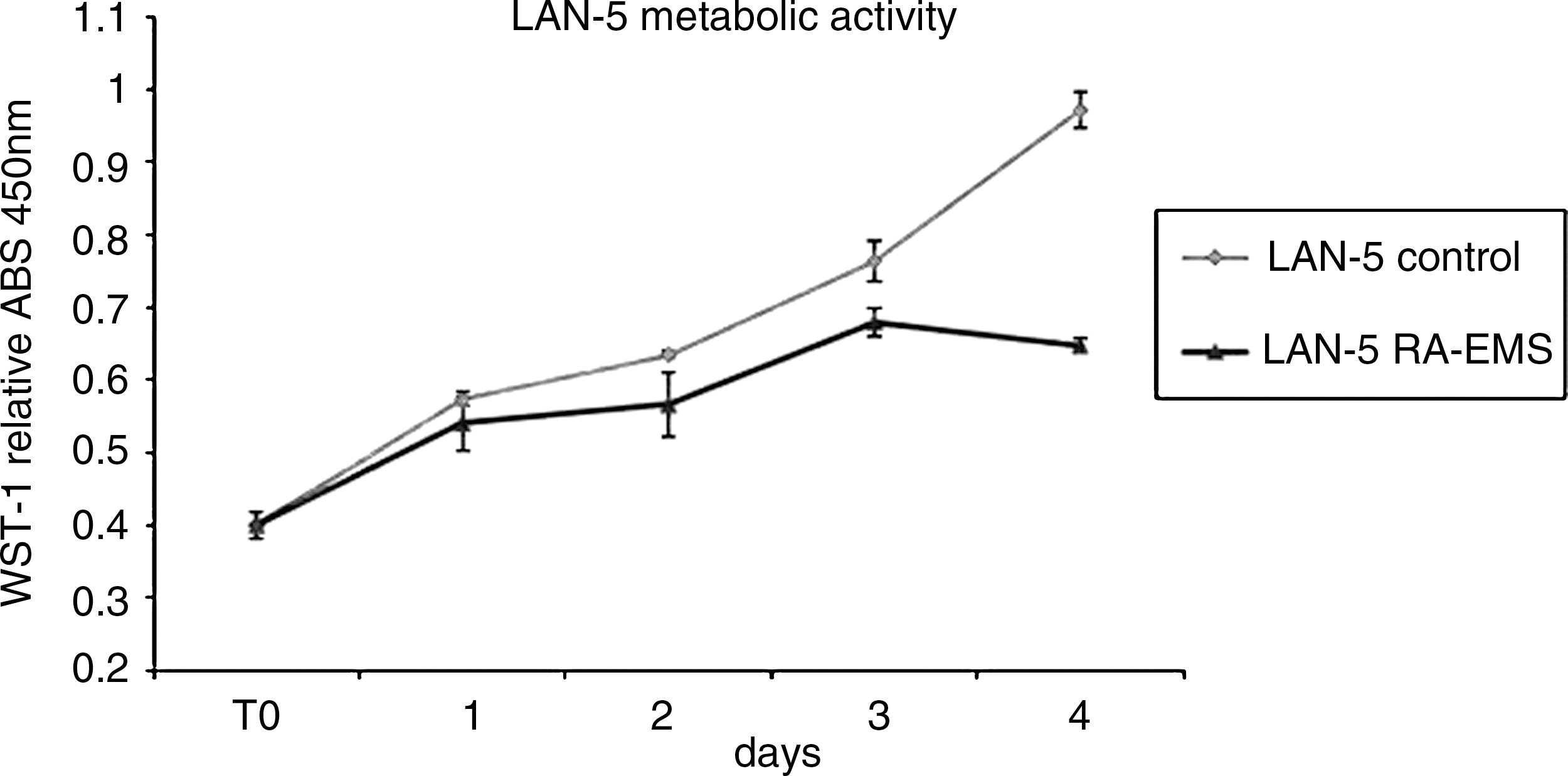
Cellular metabolic activity and proliferation by water-soluble tetrazolium salt (WST) assay. LAN-5 metabolic activity by WST-1 analysis in presence (▵) or absence (▪) of retinoic acid electromagnetic molecular signal (RA-EMS). ABS, absorbance.
Electronically transmitted RA effect on LAN-5 cell morphology
By phase contrast and scanning electron microscopy, LAN-5 control cells appeared small, polygonal, without neuritelike structures. The exposure to electronically transmitted RA induced morphological changes toward a more neuronal phenotype: The cells were stretched out and rich in neuritelike structures with blebs, mimicking the same effect induced by RA treatment (Fig. 3).
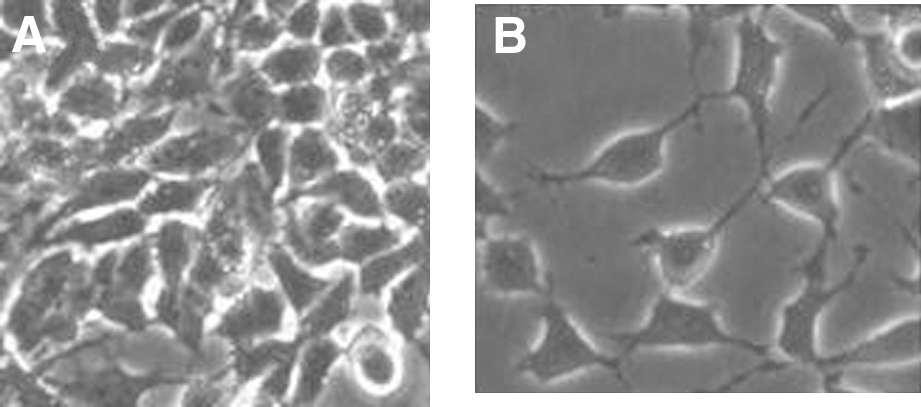
Electronically transmitted retinoic acid (RA) effect on LAN-5 cell morphology by contrast microscopy. Contrast microscopy of LAN-5 cells in absence
EMF effect on neurofilament expression
Figure 4 shows the indirect immunofluorescent analysis of control and exposed LAN-5 cells with anti-NF-200 neurofilaments. While control cells were little or not positive for NF-200 (CTR), the neurofilament protein become more fluorescent after exposure to the RA electromagnetic field (EMF) (exposed). The same results were achieved by RT-PCR analysis for mRNA expression coding for NF-200 (Fig. 5).
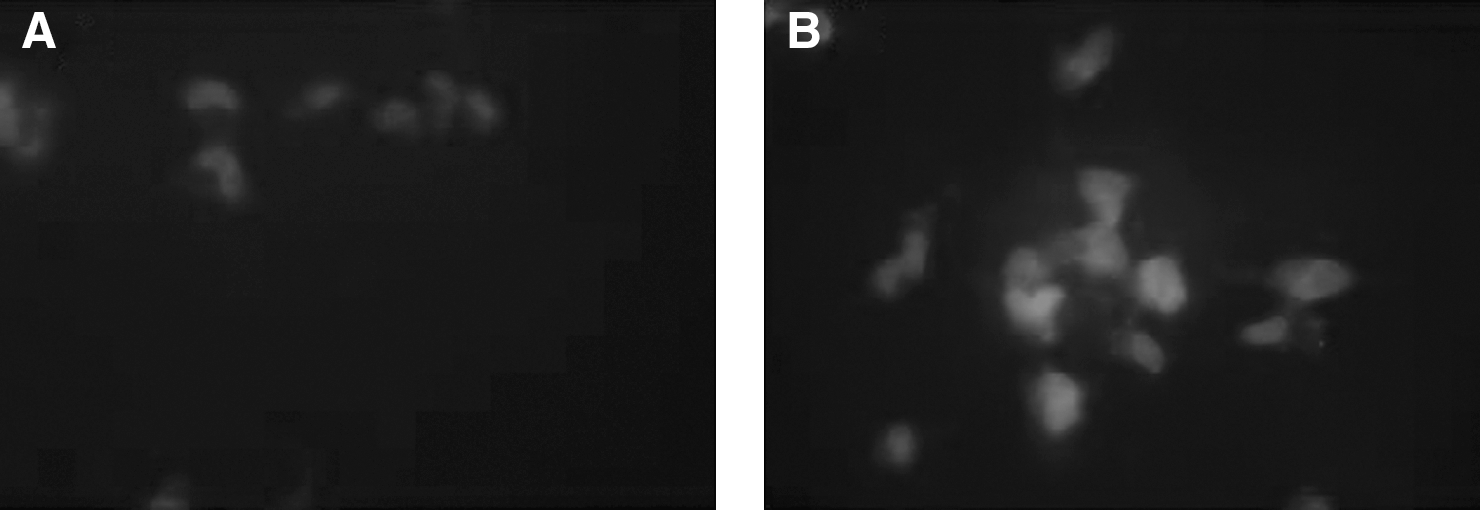
Electronically transmitted retinoic acid (RA) effect on LAN-5 cell by NF-200 indirect immunofluorescence. NF-200 indirect immunofluorescence (red fluorescence shown by white spots) of LAN-5 cells in absence

NF-200 mRNA transcription–polymerase chain reaction (RT-PCR) analysis on electronically transmitted retinoic acid (RA) effect on LAN-5 cell. Left bar
Discussion
Low-frequency electromagnetic fields at 50 or 60 Hz indeed are reported to stimulate nerve regeneration 3 and alter gene transcription, 4 and they may also play a synergistic role in cellular processes that are already activated, such as cell proliferation. 5 Despite an increasing number of publications demonstrating an effect of very low-frequencies EM field on biologic systems, other in vivo and in vitro studies suggest opposite results; in addition, the possible interaction mechanism is not yet completely understood.
A possible mechanism evoked to explain the mechanism of EM field action to biologic systems involves Ca2+ transport across cell membrane, to trigger the signal transduction cascade. 6
Electromagnetic therapeutic potential can be seen in the proven efficacy of low-energy pulsed magnetic fields in nonunion bone fracture healing, confirming that under certain conditions non-ionizing electromagnetic energy can influence physiologic processes in organisms. Physiologic paradigms for nonionizing radiation effects are required. Clues may be found in the mechanisms by which the EM field interacts with cultured cells under controlled laboratory conditions and by correlating in vivo evidence with in vitro data. 7 Brain maturation depends on a sequence of postnatal events. 8 Brushart et al. 9 found that electrical stimulation at 20 Hz promotes motoneuron regeneration, confirming a previous finding of the use of an electric field for the orientation and growth of neuritis. 10,11
Exposure to an RA-EMS field 12 hours a day for 5 days has significant effects on cell proliferation, leading to a 30% decrease of cell metabolism (Fig. 2). The decrease in cell metabolism is typical of cells, while they are blocked in G1 and in G0 phase, during the differentiation process. In addition, the decrease in cell metabolism during RA-EMS electromagnetic field exposure generated a morphological change, as reported by the contrast microscopy study shown in Figure 3.
In particular, contrast microscopy analysis end indirect immunofluorescence (Fig. 4) and RT-NF-200 PCR (Fig. 5) showed a more neuronal morphology characterized by the development of neuriticlike processes in the exposed cells compared to control. Immunofluorescence using monoclonal antibodies for the major neurofilament proteins NF-200 unequivocally demonstrated an increase in synthesis and accumulation of these neuronal proteins in the RA-EMS exposed cells. Taken together, all these data support an evident effect of the electronically transmitted retinoic acid (RA-EMS) electromagnetic field of driving neuroblastoma cells toward a neuronal differentiation, which resembles the effect determined by morphogens, such as RA in its chemical form. The possibility to induce differentiation elicited by this system through an extremely low-frequency electromagnetic field represents an effective, minimally manipulating, and safe biotechnological tool to improve neurogenic differentiation in neurodegenerative diseases.
Footnotes
Acknowledgments
This work has been partially supported by a grant from NAMED.
Disclosure Statement
No competing interests exist.
Alberto Foletti and Mario Ledda contributed equally to this article.