
Abstract
Objectives:
To replicate the phantom leaf effect and demonstrate a possible means to directly observe properties of the biological field.
Design:
Thirty percent to 60% of plant leaves were amputated, and the remaining leaf sections were photographed with corona discharge imaging. All leaves were cut before placement on film. A total of 137 leaves were used.
Subjects:
Plant leaves of 14 different species.
Results:
Ninety-six phantom leaf specimens were successfully obtained; 41 specimens did not yield the phantom leaf effect.
Conclusions:
A normally undetected phantom “structure,” possibly evidence of the biological field, can persist in the area of an amputated leaf section, and corona discharge can occur from this invisible structure. This protocol may suggest a testable method to study properties of conductivity and other parameters through direct observation of the complete biological field in plant leaves, with broad implications for biology and physics.
Introduction
T
Corona discharge is induced on an object by placing it adjacent to a high-voltage electrode plate. The ionization of the air molecules caused by the high-voltage electrical arcs induced by the high-voltage field is recorded as a light emission directly onto light-sensitive film. Light emission results from the photoionization of the air. It is a form of direct-contact photography.
The phantom leaf effect was originally described by Adamenko and reported by Tiller 2 and Ostrander and Schroeder. 3 Early experimental replications were published by Hubacher and Moss 4 and Chaudhury et al. 5
Other reports in the literature included that by Wagner, who produced a phantom leaf image 4 that more closely met the need for a clear, unambiguous image of the removed portion of the leaf. Individual samples of cut leaf images were also reported by Hartley, Hillborn, and Silva 6 and Davis, 7 and multiple samples were reported by Detrich. 8
This paper is the first to describe high-definition photographic samples of phantom leaves with large numbers. A total of 137 phantom leaf samples were attempted, and 96 (70%) clear phantoms were obtained. Previous studies had reported only very small numbers or individual samples.
The phantom leaf appears to be a normally undetectable coherent structure that underlies the physical leaf, with the properties of conductivity. The experimental protocol described in this paper may generate a fruitful method for multiple avenues of research.
Materials and Methods
Materials
The study apparatus consisted of a high-voltage–generating circuit that charged an electrode plate assembly (Fig. 1). The three sections of this device are the oscillator, the inverter, and high-voltage generation. The inverter generates a square wave pulse that drives the step-up transformer, and this in turn is full wave rectified into a direct current. This higher voltage is then made available to the anode of the silicon controlled rectifier (SCR). The SCR in turn is oscillated on and off by the pulses applied to the gate of the SCR. The higher voltage from the step-up transformer is then pulsed into the autoignition coil, which in turn creates high voltage of a 10-kV, decaying sine wave, which drives the final electrode assembly.
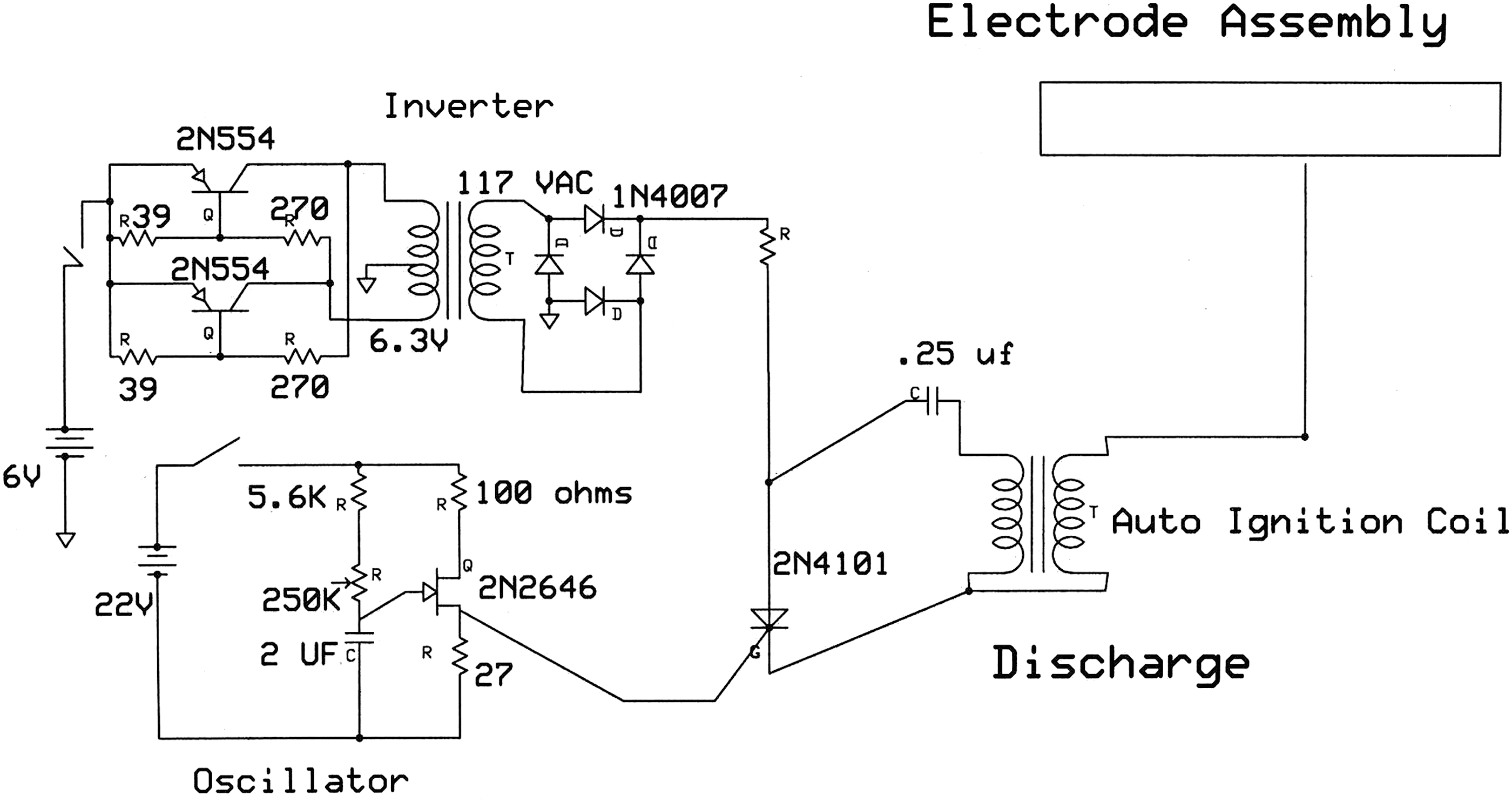
Electronic circuit diagram for high voltage apparatus.
The study used 10 kV as the peak voltage. The class of the waveform was a decaying cosine wave, typical of car-coil ring-down waveforms. A 6-V (early versions were 6-V) car coil was used from a Ford Pinto. A picture of the waveform could not be obtained for this study; such a picture would be valuable and is recommended for future studies to allow replication.
The frequency used was 100 Hz. The electrode assembly was made by using a 0.5-inch by 12×12–inch Lucite plastic sheet as a base plate. Upon this a tin foil sheet, 10×10 inches, was placed. On top of the tin foil was placed a 1/8-inch-thick Lucite plastic sheet, 12×12 inches. The electrode was thus sandwiched between two plastic sheets (Fig. 2). A wire with alligator clip was attached to the tin foil, from the high-voltage car coil.
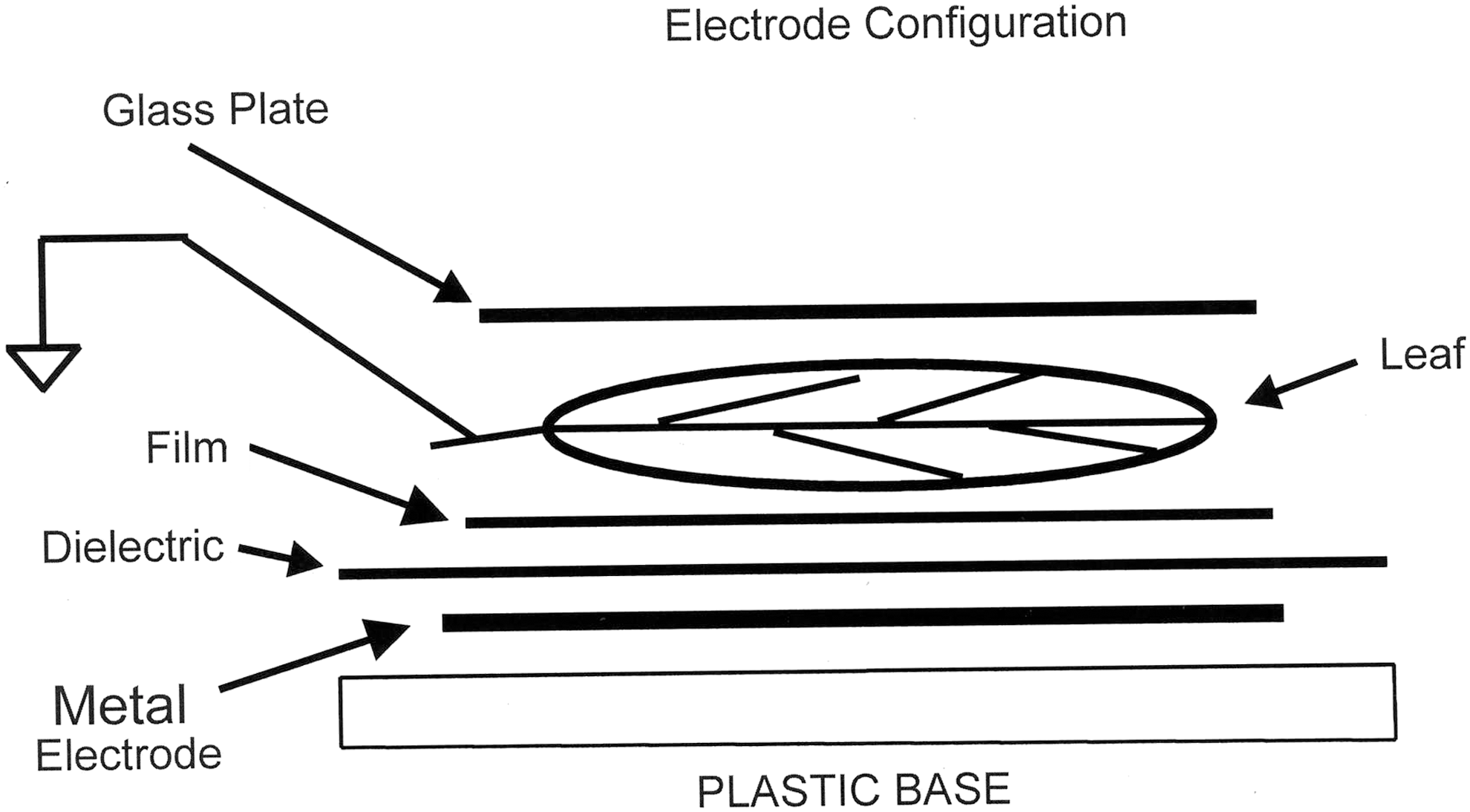
Electrode configuration.
A 4×5–inch film was placed on top of the 1/8-inch dielectric. The cut leaf to be photographed rested on top of the film. A 1/16-” 4×4–inch glass plate was placed on top of the leaf to hold it flat. Finally, a grounding wire was touched to the stem of the leaf. The stem of the leaf protruded from the edge of the glass cover plate.
The ground wire assembly consisted of a pencil-like wooden rod, to which was attached a 1-inch 26-gauge solid copper wire at the tip, held to the wooden rod with tape. Soldered to this end wire was 2 feet of 26-gauge stranded flexible insulated copper wire. The other end of this ground wire was attached to the battery ground of the circuit. This mechanism permitted the photographer to grasp the wooden rod, to which was attached the wire pointer. When the wire pointer touched the leaf stem, the leaf was effectively grounded.
Three types of film were used. Kodak 4×5–inch Tri-X orthochromatic film, which permitted the use of a darkroom red light, was used in pictures 1–115 (Eastman Kodak, Rochester, New York). Kodak Ektacolor, a negative color film, was used in pictures 116–125. For large specimens, 8×10–inch Tri-X orthochromatic film was used. Fifteen pictures were taken with this film.
Black and white film was developed in the authors' laboratory, using Dektol as a film developer. Color films were processed by a commercial laboratory.
Leaf cutting and placement procedures
Healthy leaves were chosen from a garden. Fourteen different species were used. Stems and branches of plants, with leaves intact on the stems and small branches, were place in water-filled vases. Stems remained in the vase for up to 4 hours before leaves were taken for experimentation. Leaves being used for the experiment were removed from the stem or branch, then carried to the apparatus.
There were four groups of leaves in the experiment: 1. Leaves 1–10 were photographed two times. The first photograph was a baseline of the whole intact leaf. The second photograph was of the same leaf, after a large section was cut away. The second photograph is the leaf minus the torn away section. The species of plants in this category were Ficus pumila (creeping fig), Cestrum nocturnum (night-blooming jasmine), Solenostemon hybridus (coleus), and Tibouchina urvilleana (Athens blue princess flower). 2. Leaves 11–116 were samples of leaves that had sections cut away. The original, whole intact leaf was not photographed as a baseline. The species of plant in this category were Sphaeropteris cooperi (Australian tree fern), Asclepias tuberosa (butterfly milkweed), Gardenia jasminoides (gardenia), Abutilon rubrum (red maple), Camellia japonica (camellia), Mandevilla sanderi (Brazilian jasmine), Nephrolepis exaltata (sword fern), F. pumila (creeping fig), C. nocturnum (night-blooming jasmine), S. hybridus (coleus), and T. urvilleana (Athens blue princess flower). 3. Ten cut leaves were photographed in color by using Kodak Ektacolor film, 4×5 inches. The species in this category were F. pumila (creeping fig), C. nocturnum (night-blooming jasmine), and S. hybridus (coleus). 4. Eleven specimens were large leaves (avocado) and leaves that were up to 8 inches long. These were processed on 10×12–inch Tri-X film. The species in this category were Persea americana (avocado), Acer saccharum (sugar maple), Aglaonema commutatum (Chinese evergreen), and one unidentifiable.
This report supplies pictures of two leaves from the first group and three from the second group, as a representative sample.
Placement of whole leaves for baseline photograph
A leaf was placed on the film. Either side of the leaf, dorsal or ventral, was randomly chosen. A new, unused glass cover slip was placed on top of the leaf. The grounding electrode, held by the photographer, touched the leaf stem that protruded from beneath the glass cover slip. A 1-second exposure time was given with the corona discharge apparatus. The film was numbered and set aside to be developed.
Placement of cut leaves
Following the baseline photograph of the whole leaf, the same leaf had a large section physically removed by cutting. The leaves were cut completely across the width. The procedure for cut leaves was identical, except that the leaf was cut before being placed on the film. At no time was a whole and intact leaf cut while being on the film. Cuts were created with scissors or by manually tearing the leaves apart.
The portions that were cut away were 25%–60% of the original leaf. Most leaves were cut transversely, with the tips or distal portion of the leaves being cut off and completely removed. Two leaves were cut longitudinally, with the entire right side of the leaf being removed (50% of the leaf was removed).
Only the portion of the leaf that was attached to the stem was photographed. New cover slips out of the box were used only once to prevent contamination by moisture residue.
Black and white negatives were scanned at 600 dots per inch, with 8 bits resolution of grayscale. The scanner was the Epson Perfection 4870, using Epson Scan software (Epson, Suwa, Japan). No image alterations were performed on any of the photographs shown.
Results
The total number of attempted phantom leaves was 137. Of this total, 96 phantom leaves were recorded. Almost every phantom was clear, unambiguous, and high quality, closely resembling the original leaves in detail. Most cut portions were greater than 30% of the whole original leaf. The phantom leaf images appeared to clearly represent vein and anatomic structure of the intact original leaf, and the perimeter of the original leaf. Multiple complex and detailed structures were clear.
No parameter was identified at this time to account for 41 of the specimens for which no phantom effect was recorded. Whereas multiple and interacting physiologic and electronic factors combine to allow the production of these images, it is not understood those critical for the phantom effect to occur.
Because of space limitations, selected representative samples of the photographic data are presented.
Description of Photographs
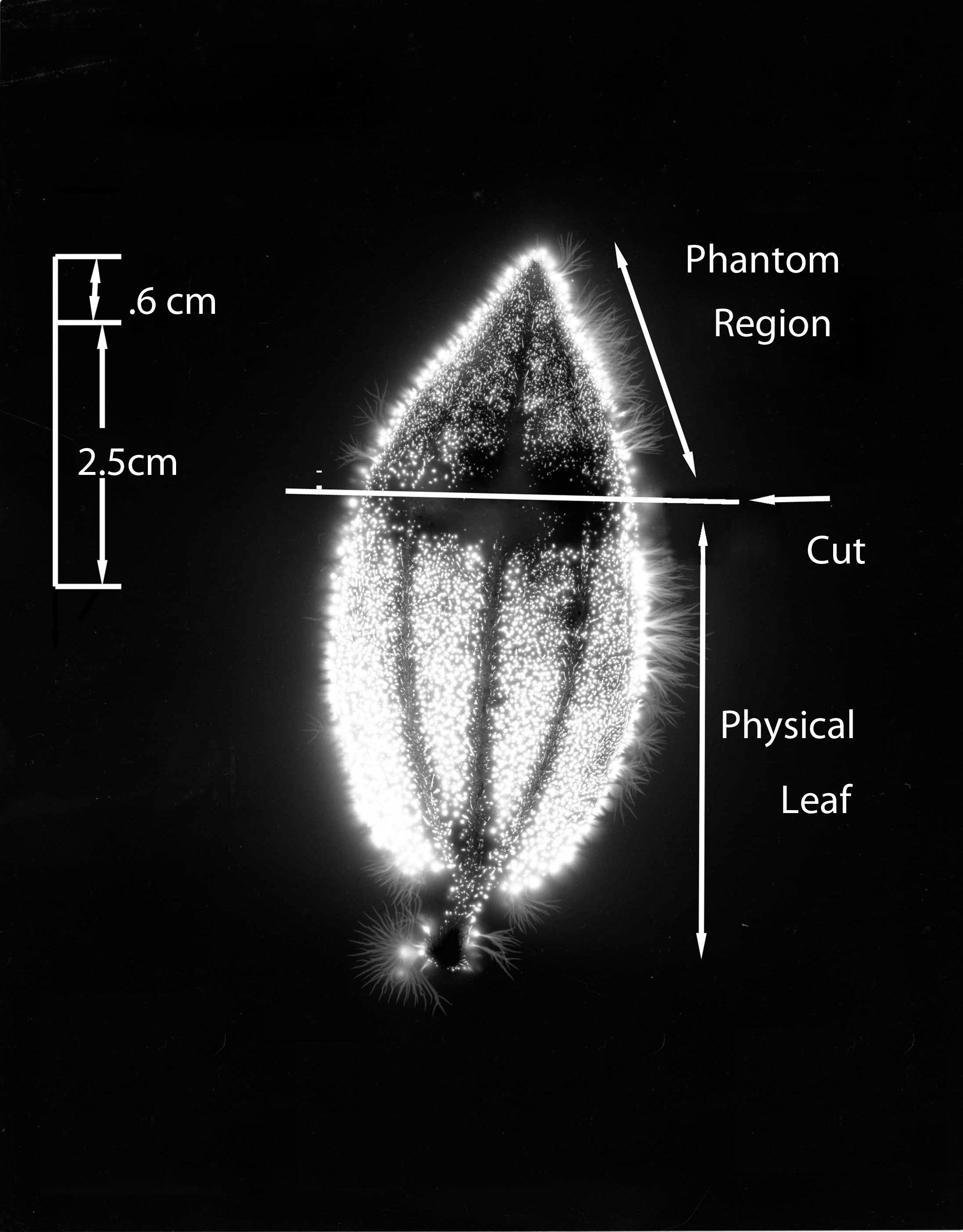
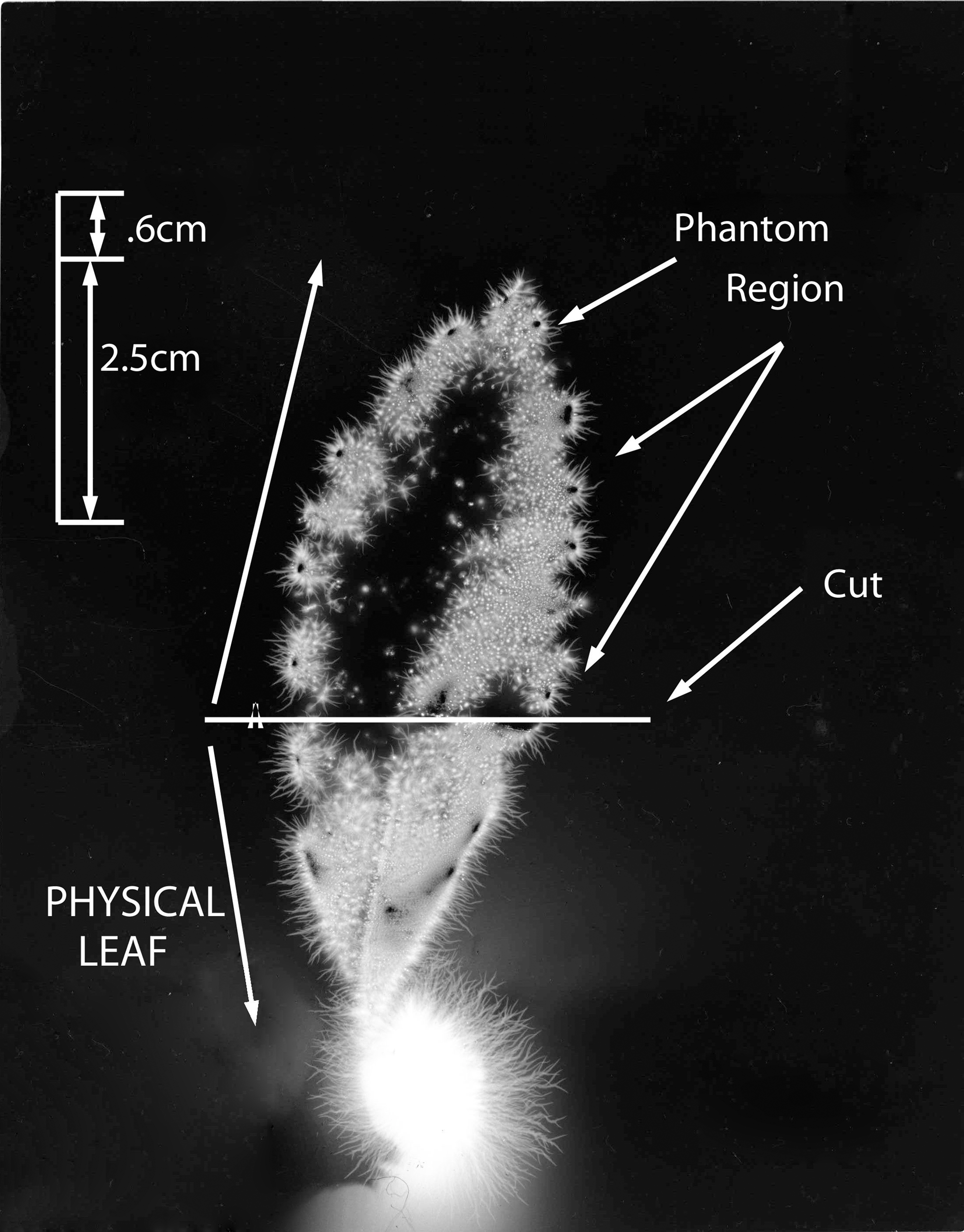
Figure 3 shows the whole #1 leaf at the baseline condition, intact and uncut. The cut form of the #1 leaf appears in Figure 4. The phantom effect, above the cut line, is visible. Figure 5 is the expanded image of the whole #1 leaf at baseline. The figure shows 10 specific anatomic features that are reproduced clearly in the subsequent phantom image after being cut away.
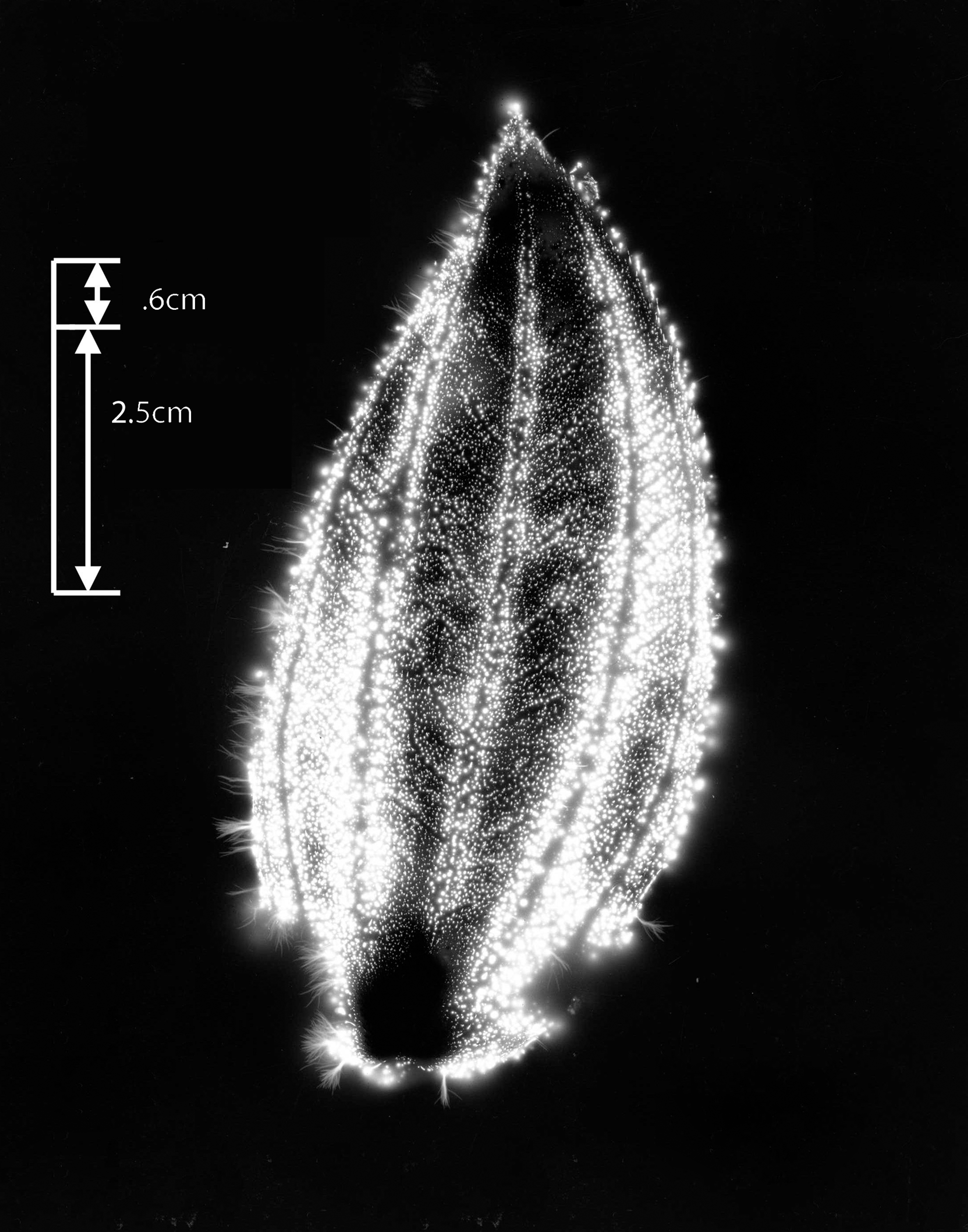
Leaf #1 (Tibouchina urvilleana). Baseline picture, intact and uncut leaf.
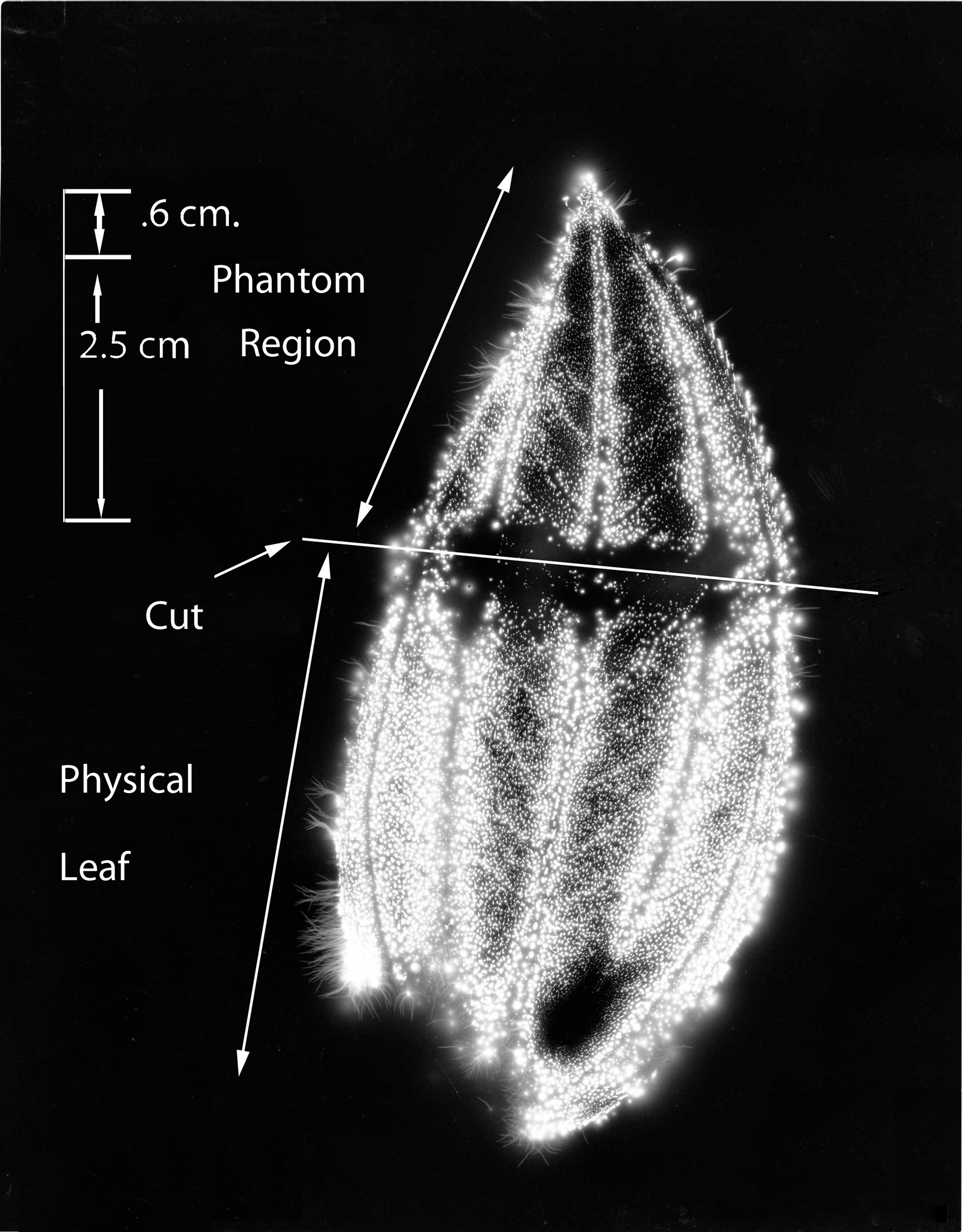
Leaf #1 (Tibouchina urvilleana). Cut leaf showing phantom effect region above the cut.
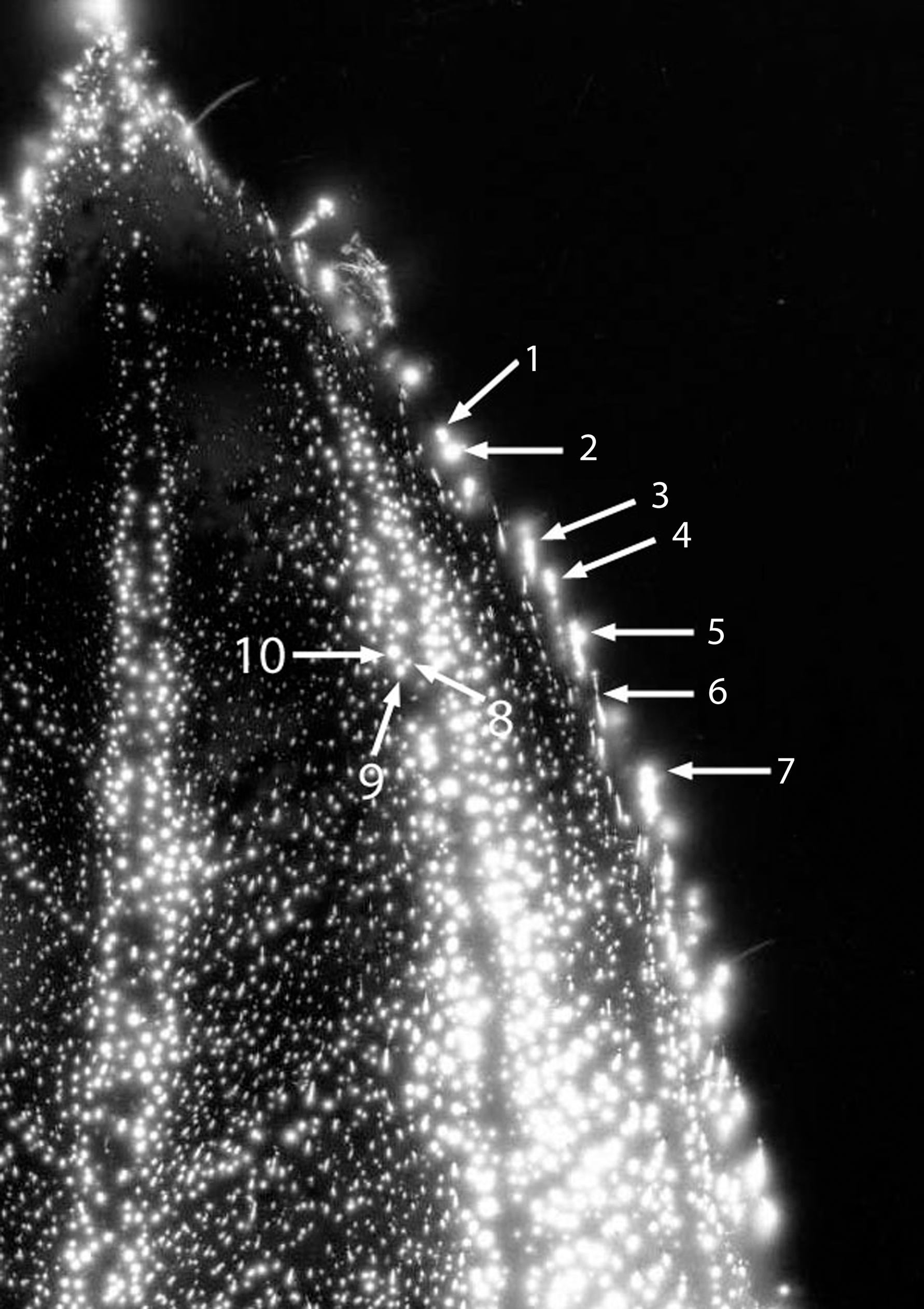
Leaf #1 (Tibouchina urvilleana). Baseline uncut leaf showing magnified view of features (1–10) before cutting away the phantom section.
Figure 6 shows the expanded image of the #1 leaf (cut). The phantom effect can be seen, with labeled anatomic features that are identical in position to the original uncut baseline leaf specimen from Figure 3. The dimensions of the smaller features are calculated to be in the range of 0.025 mm to about 0.100 mm. Thus, small and specific anatomic details are being measured.
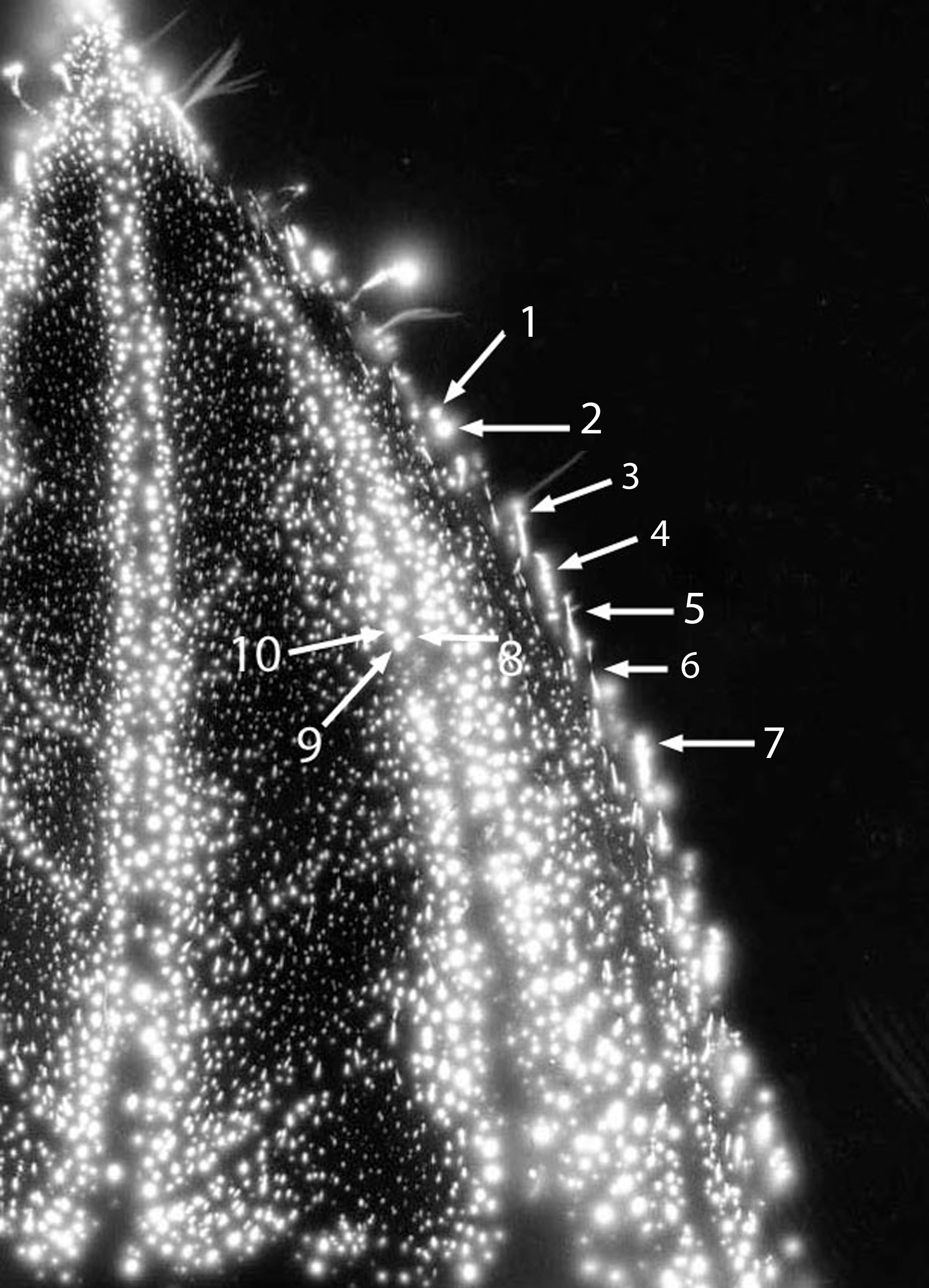
Leaf #1 (Tibouchina urvilleana). Cut leaf: phantom region magnified, with features (1–10) identical to those of Figure 5 labeled.
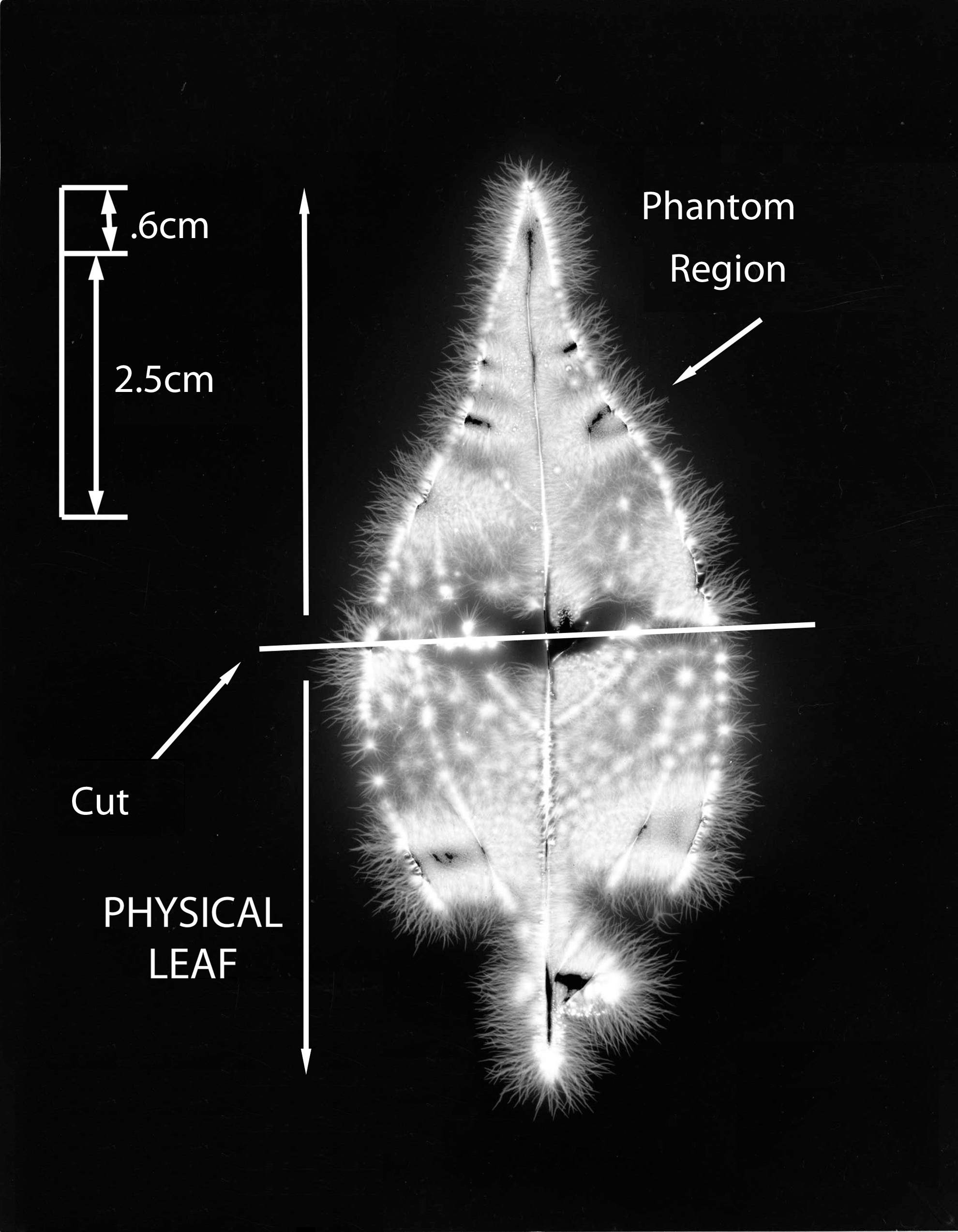
The whole #2 leaf is shown in Figure 7. This is from the baseline condition, intact and uncut. Figure 8 shows the cut #2 leaf, with the phantom effect visible. The phantom region is above the cut line.

Leaf #2 (Tibouchina urvilleana). Baseline picture, intact and uncut leaf.
Leaf #2 (Tibouchina urvilleana). Cut leaf showing phantom effect region above the cut.
Figure 9 demonstrates the #3 cut leaf, with phantom region showing and 50% of the leaf amputated. In Figure 10, the #4 cut leaf is shown, with phantom region visible and 60% of the leaf amputated. Figure 11 shows the #5 cut leaf, with longitudinal axis cut down the spine of leaf and phantom region visible. The phantom region is 50% of the leaf to the right of the cut line.
Leaf #3 (Solenostemon hybridus). Phantom leaf effect with 50% of leaf cut away.
Leaf #4 (Cestrum nocturnum). Phantom leaf effect with 60% of leaf cut away.
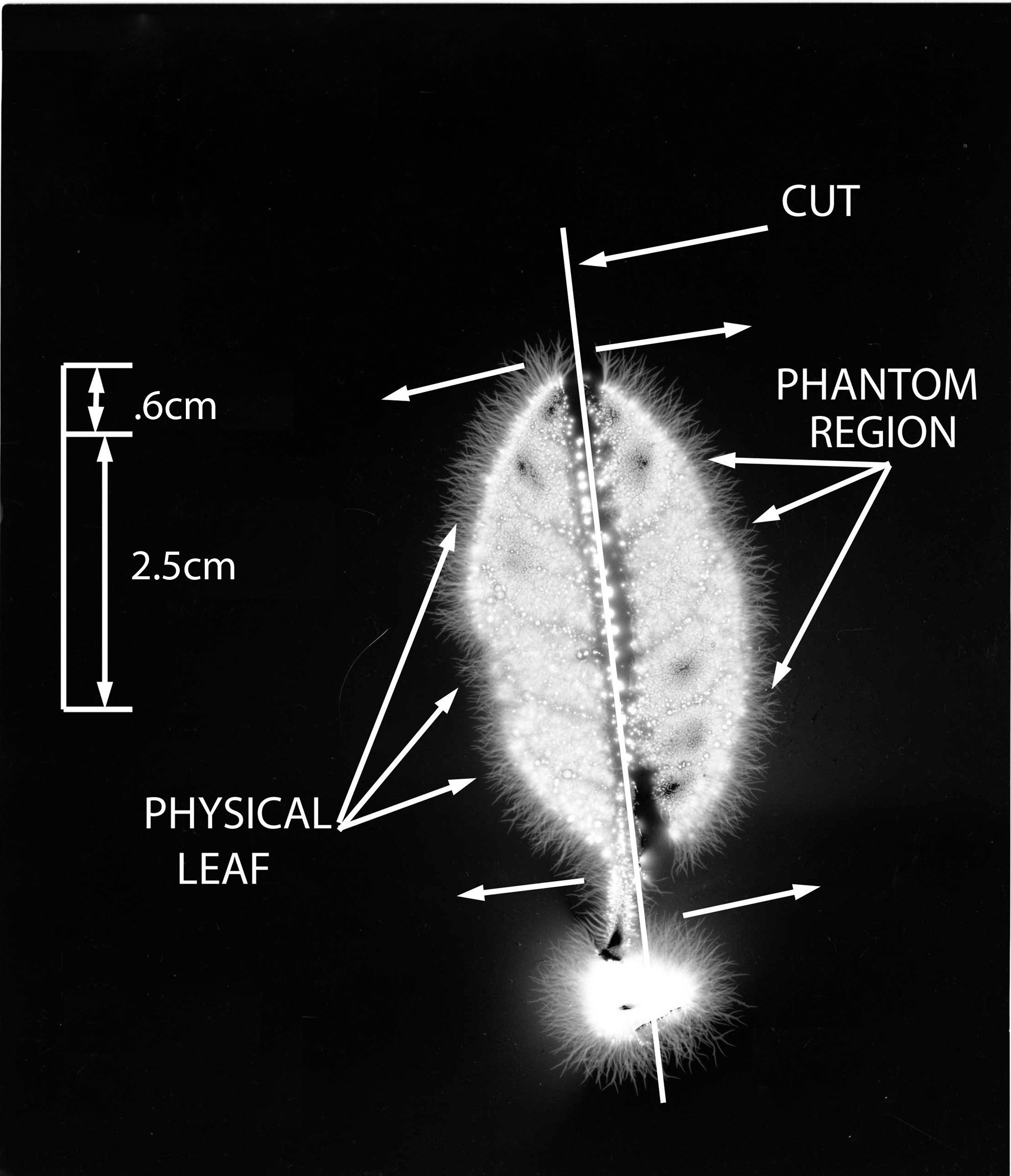
Leaf #5 (Ficus pumila). Phantom leaf effect with 50% of leaf cut away in longitudinal cut.
These samples of phantom leaf images closely or almost exactly resemble the original leaf structure, as would have appeared in a corona discharge photograph of the whole leaf, even though the original physical leaf section is no longer present or intact. The images demonstrate leaf vein structure, and the perimeter of the phantom image closely corresponds to the original leaf perimeter. Comparisons of the image of the original intact (baseline) leaf and the phantom, taken in the experimental photograph, reveal a very close correspondence between identifiable features of the two pictures (see blow up magnifications in Figs. 5 and 6).
The light intensity of the images in the original photographs is approximately the same as in the phantoms. Likewise, the streamer lengths are similar in the original and phantom photographs.
These studies show that substantial portions of the leaf can be cut away and the phantom region will appear. With horizontal cutting, up to 60% of a leaf was removed, and with longitudinal cutting 50% was removed; in both scenarios the effect was present. Compared with findings in other studies, the appearance of these phantom images are relatively reproducible.
Discussion
The phantom leaves presented in this study are complex in structure. The dimensions of phantom sections in the study ranged from approximately 2.5 cm to almost 10 cm in length, with a surface area of approximately 6 cm2 to greater than 30 cm2. From 25% to about 60% of some leaves were cut away and produced phantom images.
Each phantom leaf in this report was obtained by using procedures that eliminated currently known procedural error and artifact. All leaves in this report were cut before placement on film.
In explanation of this effect, Tiller 9 had contributed a model for the “edge effect” phantom. This conjectures that the phantom leaf effect might result from moisture emission from the cut portion driven into the space from which the cut section had been removed by the power of the field emission process.
With our current data, the high-grade images, consistent anatomic replication of the original leaf, large size, complex internal and perimeter structure, and large numbers of specimens described in this report most likely do not support edge emission artifacts as the explanation for the phantom leaf images. On the other hand, as-yet undiscovered, unsuspected artifacts other than the edge emission model may cause the phantom leaf.
A method to investigate the influence of edge effects would be to place a nonconductive sheet of Plexiglas perpendicular to the plane of the leaf and in front of the cut region. The artifactual edge effect would be blocked by the Plexiglas in this model. This setup was reported by Robert Wagner. 4 He describes the phantom as traveling through the Plexiglas sheet, unimpeded by this dielectric barrier. This is an important observation that requires additional research and replication. The edge effect would be cancelled in this model, and a dielectric barrier penetration by the phantom would be demonstrated.
The parameters necessary to reproduce this effect 100% on demand are not known. The current methods, if followed, may be more reproducible than methods used in previous studies. Clearly, some parameter or group of parameters is probably needed beyond what is understood in order to reliably reproduce these results. These include such variables as frequency, waveform, dielectric spacing, pulse widths, and types of grounding. Other variables can include film types, gases in the electrode mechanism, humidity, power sources, times of year, plant species, chemically influenced specimens (e.g., perfusion with chloroform before photography; Silva et al. 6 used chloroform-soaked cotton mingled with leaves within plastic bag before amputation to produce phantom leaves).
The bottom electrode plate, consisting of a Plexiglas cover over copper, was not cleaned between pictures. Whereas the leaves touched only the film and never the Plexiglas substrate, it is assumed there was no deposit of moisture on the bottom Plexiglas and hence no artifact problem with this procedure. However, it is recommended that this be controlled rigorously in future experiments. The Plexiglas substrate beneath the film can be cleaned and wiped after each photographic sample or changed with new Plexiglas.
The effect, tried on 14 species of plant, did not seem to be species specific. In addition, this study could not determine the relationship regarding plant leaf age (i.e., young or old).
Published reports of phantom leaves include at least eight from different laboratories and individuals. 2 –8 Various high-voltage parameters and electrode configurations have been used. Thus, the phantom leaf can be observed with multiple waveforms and frequencies and with electrode and film characteristics of the alternative devices and setups. The parameters conducive to producing the effect may therefore be broad rather than very narrow.
What are the physical properties of this phantom “structure,” or possible biological field, and what invisible “factor” would allow the conductivity of electrons in the amputated section of the leaf? Whereas this phantom effect might resemble the concept of a biological field at first glance, confirmation of this effect as representing a true field, classical or quantum, is reserved for future research. The phantom leaf appears as the corona discharge pattern of a normally invisible structure, possibly naturally underlying and permeating a normal physical leaf. As the leaf section is cut away, the action of the charged electrode causes the invisible and probably pre-existing phantom structure to conduct electrons, as corona discharge from the invisible phantom region can be observed. It thus appears that the phantom structure has at least the property of conductivity, like so many small organized wires, in a leaf pattern. How this invisible phantom structure can be organized in the absence of the physical leaf and how it remains organized as a pattern, held in space, is unknown.
The phantom leaf data suggest the existence of an intact and integral system that is conductive and permeates the whole original leaf. The system is conducting electrons in these pictures. It is distributed throughout the entire leaf; it also seems to exist independent and autonomous of the leaf in the phantom region and independent of the normal conductive mechanisms of the physical leaf. Obviously the question arises as to whether the normal known mechanisms of conductivity apply within the phantom structure. Following from this, it may be questioned whether this phantom structure functions normally as a true physiologic system, as authors have suggested would be expected of the purported biological field. This system has been postulated to deliver both energy and information systemically throughout an organism, using electromagnetic signals and forces. 10
Exogenously originated currents are being conducted in these phantom leaves. If this structure is normally present in leaves and living things, yet invisible, it is here being observed to interact with external exogenous fields. Therefore, this phantom structure may be a normal physiologically relevant system that facilitates the biological effects of externally originating electromagnetic fields.
In addition, new understandings of the mechanisms of biological conductance and impedance are suggested because these conductance processes in the phantom leaf may not be understood according to conventional explanation. This experiment may be modeling what naturally occurs: The phantom structure exists within leaves, underlying the normal physical leaf, and it electromagnetically interacts with the environment.
If the phantom structure conducts exogenous electrical currents, it probably can also conduct endogenous currents that originate in the living leaf. That is, the phantom structure might exist within the living leaf, as a normal physiological entity, in all living leaves. Following our observations, this system may have conductive capabilities for both endogenous and exogenous fields and currents. It could interface between exogenous electromagnetic influences and endogenous physiologic processes and fields, of various types. Thus, this structure would likely be assumed to play roles in the leaf's bioelectrical system, as a distributor of energy and signals within the organism, and as a distributor of energy and signals that originate outside the organism. Perhaps, this invisible structure or system is also present in animals and humans.
The medical implications for humans and animals may include eventual documentation or direct visualization of phantom limbs in amputees or documentation of an invisible electrically based physiologic system similar to the purported biofields with far-reaching consequences for disease diagnosis and treatment. As an example, if phantom limb pain is actually related to an “invisible structure” in the phantom limb region of an amputee, not only might we develop effective therapies, but we could study this “system” in humans and apply that knowledge to medicine where it is relevant.
The actual composition of the phantom structure, as well as the physical principles and relation to physics and biophysics, may be capable of being determined, as far as the corona method provides a measureable procedure for direct observations.
A possible mechanism of the phantom leaf effect may be suggested in the research described by Gariaev et al. 11 with laser spectroscopy to observe phantom DNA structures. These authors observed and recorded DNA signatures with a laser photon correlation spectrometer. The signatures that indicated perturbations in the spectroscopic signals were present for long periods after the physical DNA was removed from the measuring system. This was described as evidence for phantom DNA. The correlations between these experiments could provide a means to determine how purported phantom effects of both varieties couple with diverse electromagnetic fields. This work is possibly complementary and may show an example of another property of these phantom structures: They may be scale invariant, existing on both macroscopic and microscopic levels. The phantom structures may be operational on both size scales physiologically, with direct possible implications for quantum biology. The macroscopic phantom leaf might be composed of corresponding microscopic phantom structures, as the total phantom DNA combines to form larger assemblies of whole organisms. This hypothesis, using either of the research methods and others, should eventually be testable and potentially verifiable. The implications could extend to considering whether atomic or subatomic structures have corresponding attributes. Also, lasers might be used to enhance phantom leaf reproducibility. It is not without precedent to suggest that corona discharge might enhance phantom DNA observations: Field emission microscopy was used to originally image atoms. This technique used corona discharge procedures greatly magnified to show atomic structures. 12 Perhaps a version of field emission microscopy could be applied to observe atomic level phantom structures as a measurement technique.
The scientific antecedents for the phantom leaf are likely to include some literature on biological fields. For instance, are these photos directly observing the electromagnetic organizing field of Burr; 13 the coherent organizing field of Frohlich; 14 the bioplasma of Inyushin, 15 Zon, 16 Adamski, 17 Sedlak, 18 or Wolkowski; 19 the morphogenetic field structures of Gurwitsch; 20 or Tiller's R level 21 ? A discussion of the relevance of each possible scientific precedent is beyond this paper's scope, but the context of these experimental data may intersect with some aspects of biological field theory 22 –25 and experiment. It may also extend those theories in new directions.
Demonstrated to occur in leaves, this invisible conductive structure may also occur in animals and other living things. These findings would have implications for such disciplines as biology, medicine, consciousness studies, and physics, as researchers of the biological field have consistently postulated and discussed. 25,26
Conclusions
A normally invisible structure has been recorded in the amputated sections of leaves. This formation seems capable of photographic registration during corona discharge imaging. This structure conducts electrical charges. Up to 60% of a leaf was removed by amputation; a structure that persisted in space was recorded as light emitted during corona discharge.
This report introduced a method to more reproducibly obtain a phantom leaf image. The images presented here also support aspects of the biological field hypothesis and indicate its electrical and optical properties. Because this method contributes to reproducibility, it supports corona discharge imaging as a means for scientific scrutiny of the phantom leaf effect. This may also represent a means to directly observe, identify, and assess the properties of an invisible biological structure, partially independent of the physical organism, with the scientific benefits of a complete visual image.
Footnotes
Acknowledgments
The author acknowledges Alan Detrich for his special assistance in photographing the leaf specimens in this study and Glen Rein for consultation and advice.
Author Disclosure Statement
No competing financial relationships exist. The author in the past has manufactured Kirlian photography apparatus for research and sale through the electro medical company Pantheon Research Inc.