
Abstract
Objective:
Complementary and alternative medicine, such as Traditional Chinese Medicine, represents an efficient therapeutic option for obesity control. It was previously reported that acupuncture catgut embedding therapy (ACET) with moxibustion reduces body weight and reverts insulin resistance in obese women. This study aimed to evidence changes in adipokines and gene expression in adipose tissue that could explain the effects of ACET with moxibustion.
Design:
Overweight/obese women were treated with ACET with moxibustion or sham acupuncture as control. Peripheral blood samples and fat biopsies were taken before and after intervention. Circulating adipokines (leptin, adiponectin, tumor necrosis factor alpha, and resistin) were quantified by enzyme-linked immunosorbent assay. Gene expression in adipose tissue was determined by cDNA microarray assays and assessed by quantitative reverse transcription real-time polymerase chain reaction.
Results:
ACET with moxibustion did not modify circulating adipokines levels. However, correlations with anthropometric and biochemical parameters were affected. Interestingly, transcriptional changes in adipose tissue revealed the modulation of genes participating in homeostasis control, lipid metabolism, olfactory transduction, and gamma-aminobutyric acid signaling pathway.
Conclusions:
The effects of ACET with moxibustion on body weight and insulin resistance were associated with the regulation of biochemical events that are altered in obesity.
Introduction
O
The control of obesity requires a multidisciplinary approach, which includes a balanced diet, physical activity, pharmaceutical regimen, and bariatric surgery in selected cases. Complementary and alternative medicine represents an attractive option for weight control. Several reports have described the beneficial effects of Traditional Chinese Medicine, particularly the stimulation of specific body points (acupoints) by acupuncture and related methods, to reduce body weight through the regulation of lipids, lipoproteins, hormones, adipokines, neurotransmitters, glucose metabolism, and inflammatory markers. 5 A recent review evidenced that acupuncture catgut embedding therapy (ACET), which involves embedded surgical chromic catgut sutures into the subcutaneous tissue, has the best clinical efficacy in obesity. 6 It was previously reported by us that ACET with moxibustion was efficient to reduce and body mass index (BMI) and revert insulin resistance, which could help to control the risk of diabetes. 7 To gain insights into the molecular events underlying these effects, this study evaluated changes in circulating adipokines and transcriptional profile in adipose tissue in response to ACET with moxibustion. Results revealed the modulation of genes participating in biochemical pathways that are altered in obesity, which could explain the positive effects of ACET with moxibustion on women with obesity.
Methods
Subjects and treatments
This study included a total of 37 overweight/obese women who were recruited for a six-week weight reduction program by ACET with moxibustion or sham treatment as described. 7 The research was according to the Declaration of Helsinki, and the Ethics Committee at the authors' institution approved the protocol.
Biological samples
At the beginning and end of the protocol, a blood sample was taken from the left cubital vein after overnight fasting, and serum was stored at −20°C. Abdominal subcutaneous adipose tissue (SAT) biopsy was also obtained. After local disinfection and under anesthesia with lidocaine containing 2% epinephrine, a small incision (<1 cm) was performed in the abdomen with a sterile scalpel. A SAT sample was taken with sterile forceps and kept in Allprotect Tissue Reagent (Qiagen) at −70°C.
Adipokines determination
Circulating adipokines were determined using the Human Leptin Enzyme-Linked Immunosorbent Assay (ELISA) Kit, Human Adiponectin ELISA Kit, Human Resistin ELISA Kit (Millipore), and Human TNF-α HD ELISA Kit (Quantikine). Adipokine concentrations were expressed as mean ± standard error of the mean (SEM). Normal values for leptin, adiponectin, and TNF-α were defined as 1–15 ng/mL, 5–20 μg/mL, and <0.7 pg/mL, respectively. 8 A resistin concentration >10 ng/mL was considered as high. 9
Statistical analysis
Initial and final data in each group were compared with a paired Student's t-test. Correlations between adipokines and anthropometric and biochemical parameters previously reported 7 were evaluated by Pearson analysis. Significance was set as p ≤ 0.05.
Microarray assay
SAT biopsies of six randomly selected women were homogenized by freezing in liquid nitrogen and manual crushing. Total RNA was extracted using the RNeasy Mini Kit (Qiagen). RNA purity and concentration were determined in a NanoDrop 1000 spectrophotometer (ThermoScientific). RNA integrity was assessed by electrophoresis. dsRNA was obtained from 50 ng RNA using the Complete Whole Transcriptome Amplification Kit (WTA2; Sigma) following the manufacturer's instructions. Products were purified using the QiAquick PCR purification kit (Qiagen), labeled with Cy3 using NimbleGen One-Color DNA Labeling Kit (RocheNimbleGen), and quantified by spectrophotometry. Labeled cDNA samples were individually hybridized with a NimbleGen 12 × 135K microarray for 18 h at 42°C, according to the manufacturer's recommendations. After hybridization, the array was washed using NimbleGen Wash Buffer Kit (Roche) and scanned using NimbleGen MS 200 Microarray Scanner (Roche).
Data extraction and analysis
Data were extracted from scanned images using NimbleScan v2.6 software and aligned with a microarray archive to perform array normalization using the quantile normalization method.
10
The expression value of each gene was obtained using normalized expression values for the corresponding individual probes through the Robust Multichip-Array procedure, and data were analyzed based on the RMA processed expression values.
11
Files were exported to Microsoft Excel and processed using the Analysis of NimbleGen Arrays Interface Suite software (ANAIS;
Bioinformatics analysis
Significantly modulated genes were functionally clustered according to DAVID v6.7 (
Quantitative reverse transcription real-time polymerase chain reaction
Quantitative reverse transcription real-time polymerase chain reaction (qRT-PCR) assay was performed for six genes that were selected based on their constant up- or downregulation in the six patients, their high FC value, their low p-value, and their role in relevant pathways. The beta-actin gene was used as a housekeeping gene. Oligonucleotides for each gene were designed by Prime 3 (
From microarray assay.
qRT-PCR, quantitative reverse transcription real-time polymerase chain reaction; FC, fold change; F, forward primer; R, reverse primer.
Results
ACET with moxibustion affects correlations between circulating adipokines and anthropometric/biochemical parameters
Mean age, body weight, BMI, waist and hip circumferences, and waist/hip ratio were similar in women randomly assigned to ACET with moxibustion or sham groups. 7 As expected, circulating levels of leptin were high, while adiponectin levels were low. Resistin levels appeared high, while TNF-α levels were in the normal range. At the end of the protocol, no significant changes were observed in leptin, TNF-α, or resistin levels in either group; adiponectin was slightly reduced by ACET with moxibustion (Table 2). It was previously reported that ACET with moxibustion significantly reduces weight, BMI, triglycerides, glucose, insulin, and homeostatis model assessment–estimated insulin resistance (HOMA-IR) index in women with obesity. 7 At the beginning of the protocol, it was observed that leptin was positively correlated with BMI (p < 0.05), insulin (p < 0.01), and HOMA-IR (p < 0.05). Adiponectin was negatively associated with triglycerides and resistin (p < 0.05), while TNF-α was positively correlated with triglycerides (p < 0.05). After ACET with moxibustion, leptin was negatively correlated with insulin, HOMA-IR, and adiponectin (p < 0.05). Adiponectin remained negatively associated with triglycerides (p < 0.01) and resistin (p < 0.05), while TNF-α was positively correlated with body weight (p < 0.05) and BMI (p < 0.01). A negative correlation was also observed between leptin and adiponectin (Fig. 1).
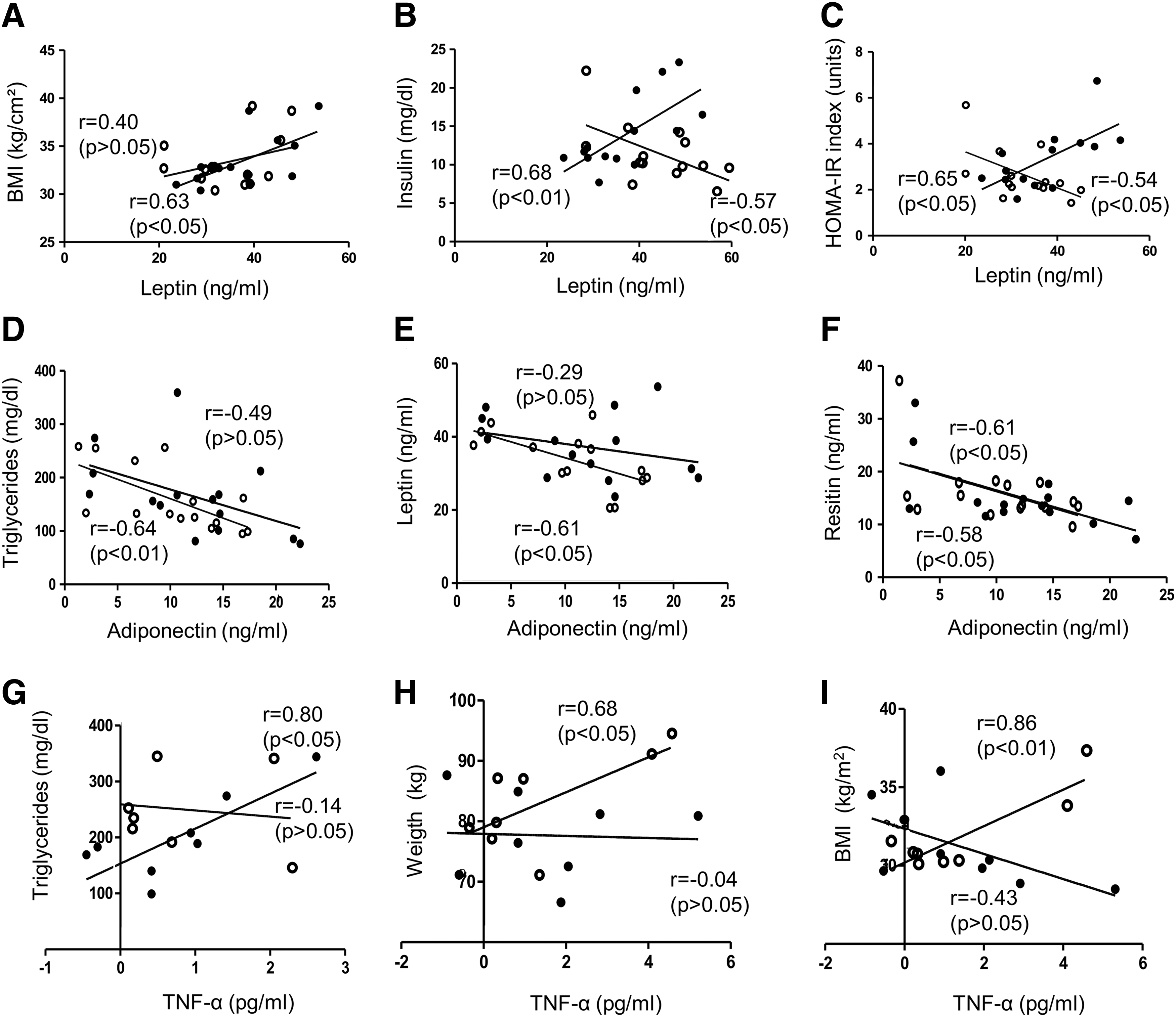
Correlations between adipokines circulating levels and anthropometric/biochemical parameters.
Values are presented as mean ± SEM.
p < 0.05 final vs. initial in each group (paired Student's t-test).
ACET-mox, acupuncture catgut embedding therapy with moxibustion; TNF-α, tumor necrosis factor alpha; SEM, standard error of the mean.
ACET with moxibustion modifies transcriptional profile in SAT
To understand the effects of ACET with moxibustion on anthropometric and biochemical parameters, the global transcriptional response in SAT was analyzed by cDNA microarrays. Of the 45,035 genes contained in the chip, 8025 (17.84%) genes were expressed in SAT, and 1791 (3.97%) genes were modulated by the intervention (1548 upregulated; 243 downregulated). A heat map evidenced the hierarchical clustering of selected modulated genes (p ≤ 0.01 and FC ≥1.5) across samples, illustrating the transcriptional response to ACET with moxibustion in SAT (Fig. 2A). qRT-PCR assays confirmed the upregulation of OR6K6, MTERFD3, and PGM2 genes, and the downregulation of PGM2, FSCB, and MMP2 genes in the six patients (Fig. 2B) as well as in the other patients (Fig. 2C).

Transcriptional response to acupuncture catgut embedding therapy (ACET) with moxibustion in subcutaneous adipose tissue.
ACET with moxibustion affects various biological processes
To identify the biological response to ACET with moxibustion in SAT, modulated genes was classified according to Gene Ontology categories. The main biological functions were biological regulation, multicellular organismal process, and regulation of metabolic process. According to cellular component analysis, modulated genes were classified as intrinsic to membrane and integral to membrane and plasma membrane, among others. Molecular functions corresponded mainly to ion binding, DNA binding, and transcription factor activity (Fig. 3). KEGG analysis revealed that 22 genes participate in cytokine–cytokine receptor interaction, 18 genes correspond to calcium signaling, and 15 to jak-stat signaling. Other affected pathways were related to cardiac function, among others (Table 3). Interestingly, functional clustering showed that modulated genes are involved in processes that are altered in obesity, such as homeostasis control (regulation of response to nutrients, homeostatic process, insulin secretion) and lipid metabolism (regulation of fatty acid metabolic process, lipase activity, regulation of lipase activity, diacylglycerol binding, phospholipase C activity, and adipocytokine signaling pathway). Another representative functional theme was olfactory transduction, with the modulation of 44 G protein-coupled receptors (GPCR) rhodopsin-like superfamily genes, including 27 olfactory receptor (OR) genes, as well as genes participating in the regulation of adenylate cyclase activity in protein-G signaling and related to cAMP metabolic process. Other modulated genes were related to the gamma-aminobutyric acid signaling pathway (Table 4).
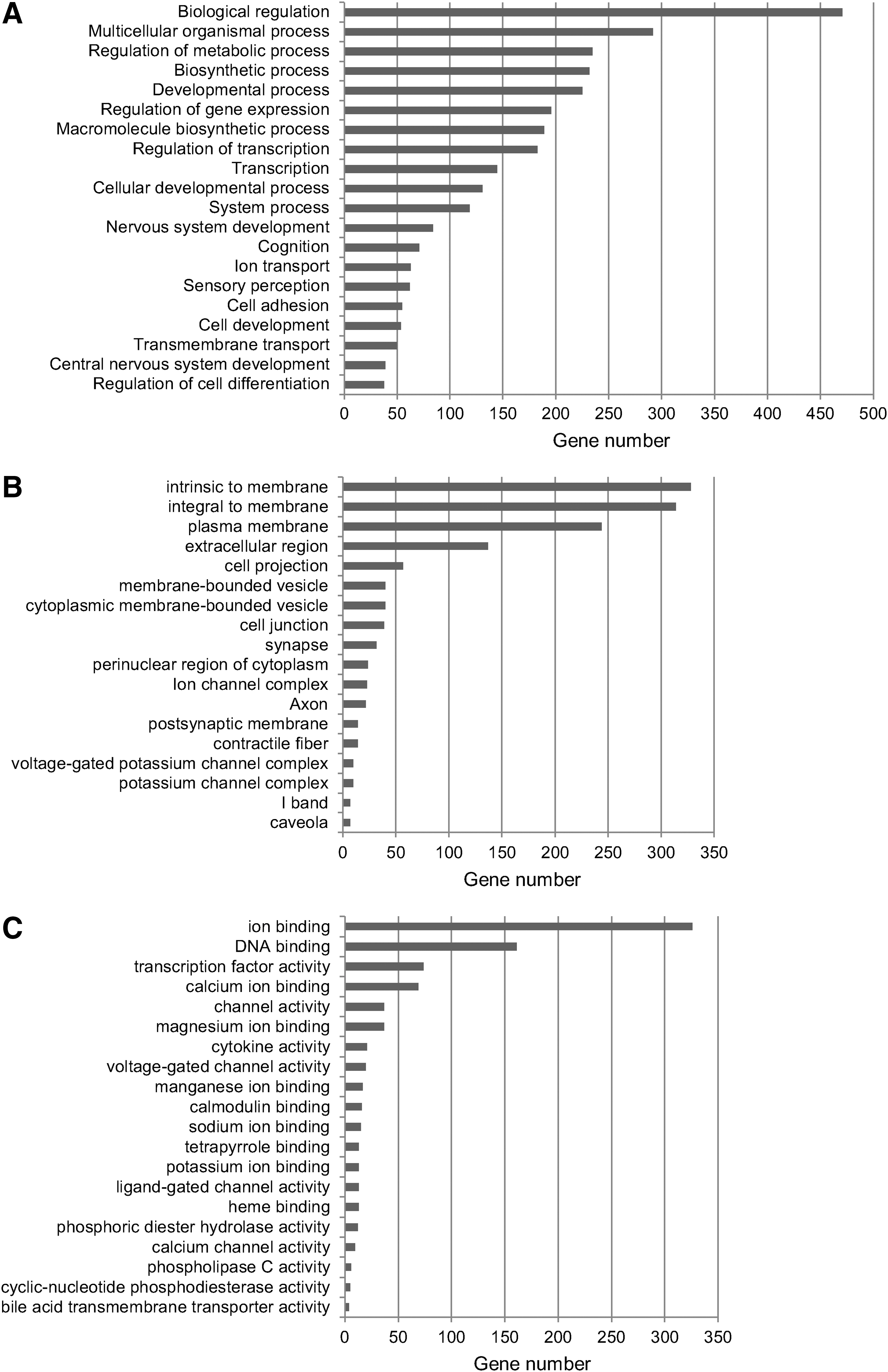
Classification of modulated genes according to Gene Ontology categories by DAVID.
FC, fold change.
Discussion
Obesity is characterized by an excess of adipose tissue and a marked adipocyte dysfunction. To gain insights into the molecular mechanisms underlying the effects of ACET with moxibustion, a traditional Chinese treatment to reduce body weight and insulin resistance, 7 the gene expression profile in SAT of obese women was examined. The results demonstrated for the first time, to the authors' knowledge, that the efficacy of ACET with moxibustion on anthropometric and biochemical parameters is associated with changes in gene expression in SAT. Surprisingly, these effects were not directly related to changes in circulating adipokines, or adipokine gene expression in SAT. Previously, Chen et al. 12 showed that ACET reduces TNF-α levels. This discrepancy could be due to differences in acupoint selection and stimulation conditions. Indeed, changes in correlations were observed between adipokines and anthropometric and biochemical parameters, which suggest that ACET with moxibustion modulated, at least partially, physiological and pathological processes related to energy intake and expenditure in SAT.
ACET with moxibustion affected the expression of a limited number of genes (3.97%) in SAT. Similarly, 2.5% and 4.09% of genes were modulated in SAT after a 10-week diet 13 and a 6-month exercise intervention, 14 respectively. However, the protocol used in the present study only lasted six weeks. The higher impact of ACET with moxibustion on gene expression may be related to the persistent stimulation produced by the suture at acupoints. The effects of proteolytic enzymes and macrophage action against the absorbable surgical thread cause mild irritation, which may improve and extend acupoint stimulation. 15
A relevant finding of this study was that ACET with moxibustion was associated with the regulation of biochemical pathways that are altered in obesity. Thus, modulated genes participate in homeostasis control, including response to nutrient levels, homeostatic processes, and insulin secretion. NPC1 (Niemann-Pick C1) controls intracellular cholesterol homeostasis by regulating lipoprotein-derived lipids (cholesterol and fatty acids) transport from late endosomes/lysosomes to other cellular compartments. Genetic studies revealed a relation between defects in the NPC1 gene and obesity. In addition, NPC1 mRNA levels are increased in fat in individuals with obesity, while they are reduced during weight loss. 16 Similarly, NPC1 expression was reduced in SAT after body-weight reduction following ACET with moxibustion, confirming the metabolic function of NPC1 in adipose tissue. GHRL (Ghrelin) participates in the hypothalamic regulation of energy homeostasis, stimulating orexigenic NPY and AgRP neuronal activity. It is predominantly synthesized in the stomach, but ghrelin mRNA and peptide have been detected in almost all tissues, including SAT. Circulating and tissue levels of ghrelin drop in diet-induced obesity, 17 which is consistent with the present results. In addition, ghrelin has a role in glucose homeostasis, inhibiting insulin release from the pancreas and promoting hyperglycemia. 18 Accordingly, the reduction of ghrelin in SAT after ACET with moxibustion was associated with a decrease of both insulin and glucose levels. 7 Peroxisome proliferator-activated receptor alpha (PPARA) regulates numerous genes of energy metabolism. A recent work revealed that PPARA mRNA levels were decreased by both genetic and HFD-induced obesity in white adipose tissue, while fasting increased them. PPARA activation promotes adipocyte differentiation and fatty acids oxidation, contributing to the enhancement of whole-body oxygen consumption and suppression of adipocyte hypertrophy. 19 Moreover, PPARA activation prevents inflammation in white adipose tissue and increases adiponectin and adiponectin receptors, which results in the amelioration of obesity-induced insulin resistance. 20 Here, it was found that PPARA was downregulated in SAT, suggesting that it did not contribute to the improvement of insulin sensibility after ACET with moxibustion. In fact, this event could be more related to the overexpression of PPARGC1A (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), CACNA1C (Calcium channel, voltage-dependent, L type, alpha 1C subunit), SLC30A8 (Solute carrier family 30; zinc transporter, member 8), NEUROD1 (Neurogenic differentiation 1), and PDE8B (Phosphodiesterase-8B), together with the downregulation of UCN3 (Urocortin 3). CACNA1 is activated in the first phase of insulin secretion. 21 NEUROD1 controls insulin expression and β-cell survival. 22 SLC30A8 is essential for insulin secretion, the regulation of hepatic insulin clearance, and the delivery of insulin to peripheral target tissues. 23 Diminished PDEB8 potentiates biphasic insulin response to glucose. 24 UCN3 is induced in pancreas under excessive caloric conditions and acts locally to increase insulin production, which may contribute to reduced insulin sensitivity and harmful metabolic consequences. 25 Moreover, PPARGC1A expression is reduced in adipose tissue in insulin resistance. 26 Therefore, the modulation of CACNA1C, SLC30A8, NEUROD1, PDE8B, PPARGC1A, and UCN3 could contribute to the control of insulin resistance by ACET with moxibustion.
Another interesting event that was affected by ACET with moxibustion was lipid metabolism, through the overexpression of PPARGC1A, PRKAA2 (5′-AMP-activated protein kinase catalytic subunit α2), and ACSL3 (Long chain acyl-CoA synthetase-3). A decreased expression of PRKAA2 has been associated with increased body weight and fat mass, as well as impaired insulin sensitivity in high-fat-diet mice. 27 Similarly, a reduced PPARGC1A expression in adipose tissue has been associated with obesity, fat mass accumulation, and insulin resistance. 28 In the liver, the activation of ACSL3 reduces cholesterol and triglycerides. 29 Accordingly with these reports, the upregulation of PRKAA2, PPARGC1A, and ACSL3 after ACET with moxibustion could regulate the perturbed fatty acid metabolism, which is an underlying contributor of obesity. This could also help to control insulin resistance. 7 ACET with moxibustion was also associated with activation of phospholipases c (PLCB4, PLCXD3, PLCZ1, and PLCB1) that hydrolyze phospholipids into fatty acids and other lipophilic compounds, playing an essential role in signal transduction pathways through the release of diacylglycerol (DAG) and the second messenger inositol triphosphate. DAG can also be phosphorylated to form the lipid second messenger phosphatidic acid (PA). Notably, disturbances in DAG/PA balance may impair cellular metabolism, including insulin sensitivity. 30 Although the direct role of diacylglycerol kinase (DGK) isoforms in insulin sensitivity and diabetes is not well known, it seems that DGKd contributes to hyperglycemia-induced peripheral insulin resistance. 31 Therefore, it is possible that the activation of DGK in response to ACET with moxibustion could contribute to insulin resistance control through the inhibition of DAG accumulation. Interestingly, the activation of GPCR by ligand binding stimulates adenylate cyclase and intercellular levels of cAMP rise. Interestingly, alterations in the adipocyte adenylate cyclase regulation inhibit lipolysis, promoting obesity. 32 Congruently, body weight reduction after ACET with moxibustion was associated with the stimulation of non-odorant GPCR, such as CHRM5 and CHRM2 (Muscarinic acetylcholine receptors M5 and M2), HTR7 (5-HT7 receptor), and TSHR (thyrotropin receptor). HTR7 and TSHR are expressed in various tissues, including fat. In the periphery, 5-HTRs are necessary for serotonin functions in glucose and lipid metabolism. On the other hand, a deregulated expression of TSHR in adipose tissue is associated with obesity, as a result of an increased adipogenesis. 33 Therefore, the stimulation of non-odorant GPCR may promote lipolysis, contributing to body weight reduction in response to ACET with moxibustion.
On the other hand, the transcriptional response to ACET with moxibustion evidenced the activation of the olfactory transduction pathway with the overexpression of 27 ORs. Although most components of olfactory-like chemosensory signaling are expressed in the olfactory epithelium, they are also present in non-chemosensory tissues. 34 Recently, the expression of several ORs in SAT and muscles has been related to the progression of obesity. 35 In contrast, it was observed here that OR activation in SAT was associated with body-weight reduction in response to ACET with moxibustion. Further analysis is required to understand the relevance of OR in obesity.
Finally, the results presented here evidenced the activation of the gamma-aminobutyric acid (GABA) signaling pathway mediated by the upregulation of five subunits of GABR (GABA receptor subunit). GABR are expressed in neuronal and non-neuronal tissues, included SAT. 36 A recent study revealed that oral administration of GABA inhibited high-fat diet-induced obesity and improved glucose intolerance and insulin sensitivity, even after the establishment of obesity and diabetes. Furthermore, it reduced adipocyte hypertrophy and fat mass. 37 Therefore, the activation of peripheral GABA receptors by ACET with moxibustion, such as GABA does, could activate signaling pathways to promote body weight and insulin resistance reduction.
Conclusion
The effects of ACET with moxibustion on body weight and insulin resistance did not modify adipokine expression in women with obesity. Importantly, transcriptome profiling of SAT revealed the modulation of genes involved in biochemical events that are altered in obesity, leading to the improvement of anthropometric and biochemical parameters. Further experiments are required to elucidate the exact molecular mechanisms of ACET with moxibustion for obesity control.
Footnotes
Acknowledgments
We thank the Mexican patients who participated in the study. We also thank F. Lozano-Rodriguez, MD, PhD, and E. Abrego-Mena, MD, for patients' recruitment and treatment application, as well as M. C. Garcia-Cardona, MD, MsC, for obtaining SAT biopsies. This work was supported by grants from CONACyT (113148) and SIP-IPN (20131116 and 20143948), Mexico. J.M.G.-V. received a CONACyT fellowship. C.G.-H. and L.A.M. were supported by COFAA and SIP-IPN, Mexico.
Author Disclosure Statement
No competing financial interests exist.