
Abstract
Abstract
Objective:
The purpose of this neurophysiological study of 8 volunteer subjects was to see if electrical stimulation of the auricle of the ear might result in objective somatic electromyographic reflexive activity in the dorsal cervical region.
Materials and Methods:
Two auricular sites were stimulated in each ear in most of 8 human subjects (5 males and 3 females), and a third site was stimulated in the author. Recordings were made to determine if the reflex activity occurred.
Results:
This study documents the presence of a skeletal muscle electromyographic reflex, named the human auriculo–cervical reflex, in 5 male and 3 female subjects. This subclinical reflex is recorded with surface electrodes over suboccipital muscles at the posterior neck, and is evoked by electrically stimulating apical, conchal, and lobular portions of the auricle of the ear. The reflex may occur bilaterally on unilateral stimulation, and has variable early, intermediate, and late components.
Conclusions:
The identification of this reflex may help advance understanding of the reflexive nature of auriculotherapy and may potentially have neurological diagnostic utility.
Introduction
The primary purpose of the present study was to demonstrate somatic auriculo–cervical reflex (ACR) activity produced by electrical stimulation of the human pinna.
In humans, electromyographic (EMG) trigemino–cervical reflexes have been described.7–11 These are most commonly obtained by stimulating the supraorbital or infraorbital branches of the trigeminal nerve, with EMG surface-electrode or needle recording at the ipsilateral splenius capitis, semispinalis capitis, and sternocleidomastoid muscles. What is also pertinent to the investigation of an ACR reflex is the observation that mental-nerve stimulation on the face may also result in a trigemino–cervical reflex, although this happens less consistently than occurs with facial supraorbital nerve or infraorbital nerve stimulation. Such reflexes are relevant to this discussion, as the trigeminal nerve provides a significant portion of sensory innervation to the auricle of the ear, particularly through its mandibular division.
The ACR reported herein is distinct from somatic reflexes evoked from direct intraoperative stimulation of the greater auricular nerve in the cat, or from direct stimulation of the supraorbital, infraorbital, and mental branches of the trigeminal nerve in the human face. In addition, the human ACR as described in this report is different from Snellen's reflex, sometimes called the “auriculocervical reflex.” Snellen's reflex consists of “unilateral congestion of the ear upon stimulation of the distal end of the divided auriculocervical nerve” 12 (note that the “nervus auricularis cervicalis” is an older name for the greater auricular nerve. 13 ) Snellen's reflex is an autonomic reflex of induced vascular congestion of the ear and is not a somatic EMG reflex.
The auricle of the human ear is richly innervated. The principal sensory innervation of the apex region is provided by the mandibular division of the trigeminal nerve (V3); of the conchal region by the auricular branch of the vagus nerve (X); and of the lobular region by the greater auricular branch of the superficial cervical plexus (SCP). A secondary purpose of this study was to determine if focused, electrical stimulation of these three specific auricular areas of innervation might produce ACRs, and to see if any such evoked somatic reflex activity might vary according to the specific auricular area stimulated in this fashion.
Materials and Methods
This study involved 8 healthy individual volunteers, including 5 males and 3 females. Their ages ranged from 15 to 70. The 15-year-old subject was the son of the author. All other subjects were adults. Subjects were selected to sample a wide age range, involving both genders. The nature of the study was explained, and oral informed consent was obtained from all participants.
Each subject was seated in a chair. Silver cup electrodes were placed vertically over the splenius capitis muscles bilaterally, with the G1 recording electrode above, at the level of the C2 spinous process, and the center of the G2 reference electrode recording 3 cm inferior to the center of G1. The ground electrode was placed on the arm ipsilateral to the ear being stimulated. All stimulating and recording electrodes were secured with tape.
In the majority of subjects, two auricular sites were stimulated in each ear. The first site was at the apex of the ear, with the S1 (cathode) on the lateral (external) surface of the helix and the S2 (anode) on the medial (internal) surface of the helix. The second site was at the middle of the earlobe (lobule), with S1 placed laterally and S2 placed medially. This lateral-to-medial surface stimulation of the pinna was formulated to focus stimulation on relatively discrete areas at the apical helix of the ear and at the earlobe.
In 1 individual, the author, a third site was also stimulated, which was the concha of the auricle. Other subjects were not tested routinely with conchal stimulation, because such stimulation induced facial nerve reactivity and, consequently, a gross clinical facial movement artifact. Conchal stimulation was carried out by placing the S1 electrode in the superior concha and the S2 electrode in the inferior concha. Lateral-to-medial stimulation across the concha would not be feasible in the absence of a deep-needle stimulatory electrode, because of the attachment of the ear to the cranium (although it might possibly have avoided facial-nerve contamination). Only surface stimulation and recording techniques were used for simplicity (and to help recruit volunteers). Figure 1 illustrates the placement of S1 and S2 surface stimulating electrodes at the apical helix, lobular, and conchal sites.
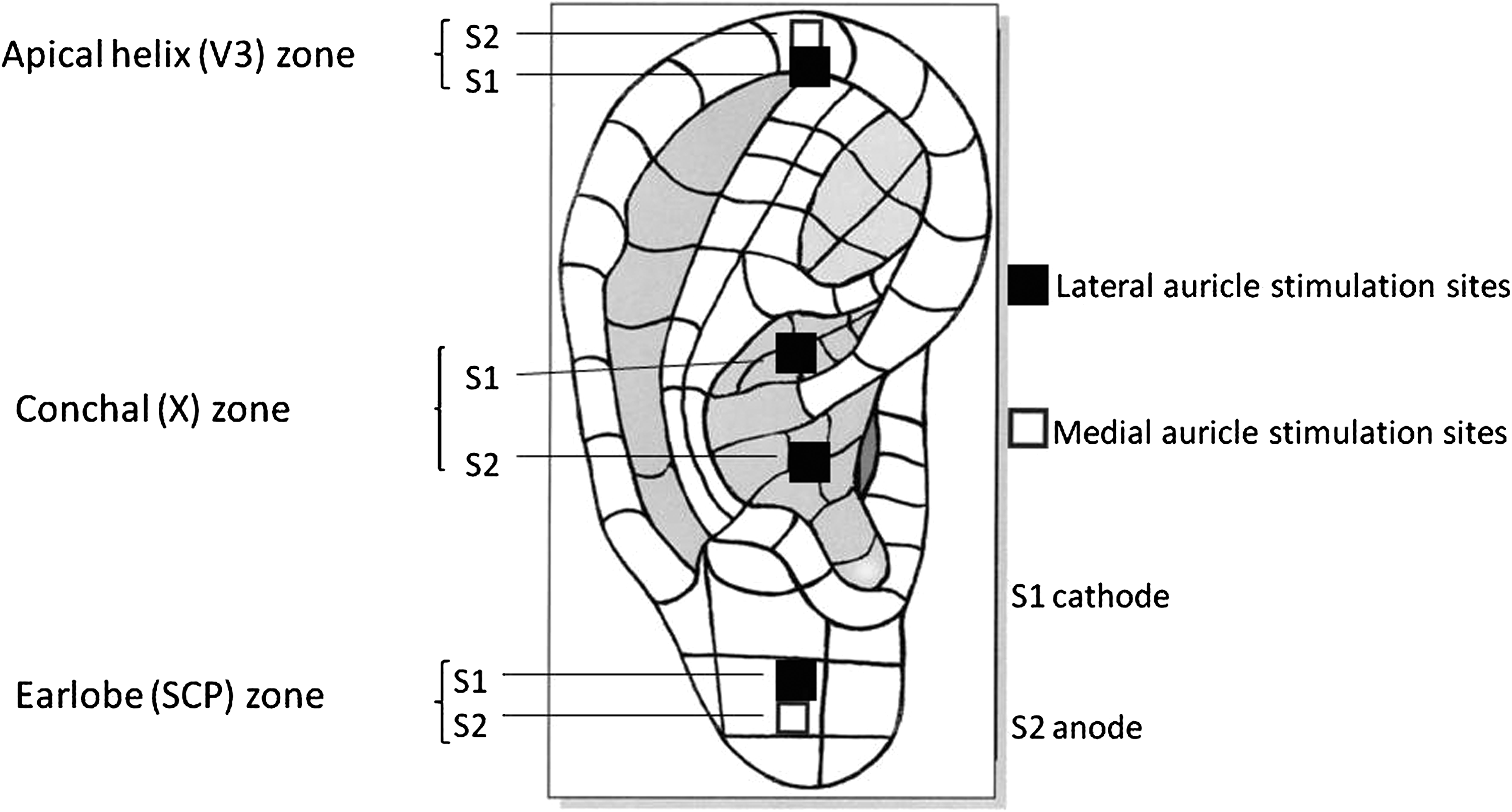
Setup for auricular stimulation. SCP, superficial cervical plexus.
A TECA Synergy EMG instrument and stimulator were used (Oxford Instruments Medical, Surrey, United Kingdom). The seated subject was asked to gently flex his or her neck 30°–45°, positioning that tended to facilitate the appearance of ACRs. Rectangular electrical shocks with a duration of 0.1–0.2 ms were delivered. The analysis time was set initially at 200 ms, the voltage was set initially at 100 uv/div, and the interstimulus interval was approximately 5 seconds. The filter bandpass was 3 Hz–10 kHz. The intensity of the shocks was gradually increased to achieve a maximum current of 80–100 mA. Most individuals found this mildly painful. For each auricular stimulation site, 20 averaged EMG responses at the splenius capitis muscles in the suboccipital region were recorded bilaterally and simultaneously. This methodology was selected to establish the presence or absence of ACRs, and their characteristics to surface stimulation of specific auricular target sites.
In 1 subject, the author, raster-format stimulation of the ACR was carried out to demonstrate the variability of its morphology from stimulus to stimulus. The same results were also recorded in a superimposed format, for the same purpose (Fig. 2).
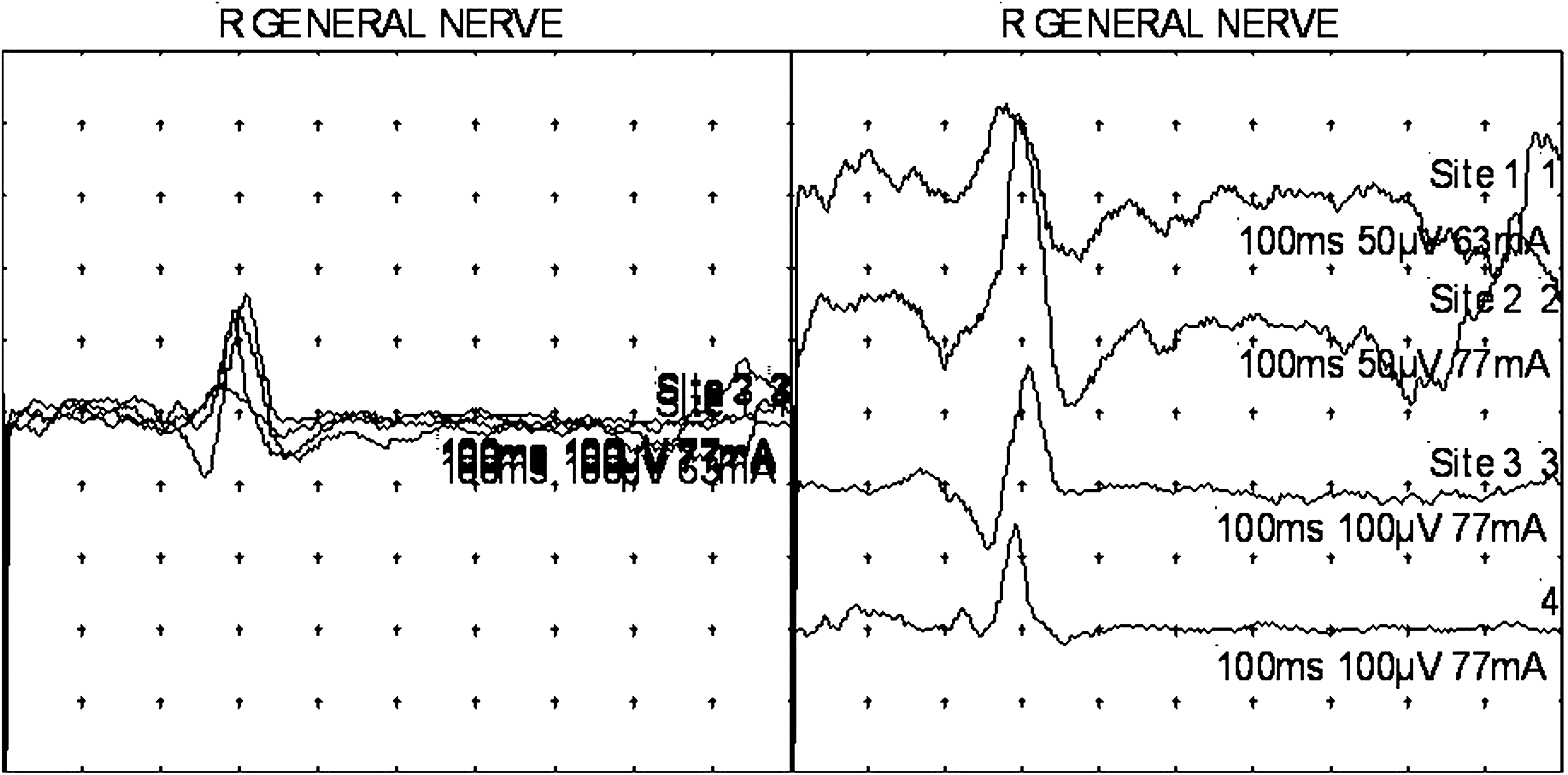
Intermediate latency (T2) auriculo–cervical reflexes obtained via four lobular stimulations, recording over the ipsilateral splenius capitis muscle, in a superimposed mode (left; L), and in a raster mode (right; R). Note that some of the responses have an initial positive (downward) deflection and that some have an initial negative (upward) deflection. Superimposed numbers are the result of these images being computer-generated.
In the author, dual-channel recordings were carried out to assess simultaneous responses ipsilaterally at the postauricular region and at the suboccipital region (Fig. 3), and ipsilaterally at lower posterior cervical, thoracic, and lumbar paraspinal regions. This methodology was chosen to screen for possible auriculo–somatic reflexes below the suboccipital region, in the low cervical, thoracic, or lumbar paraspinal regions.
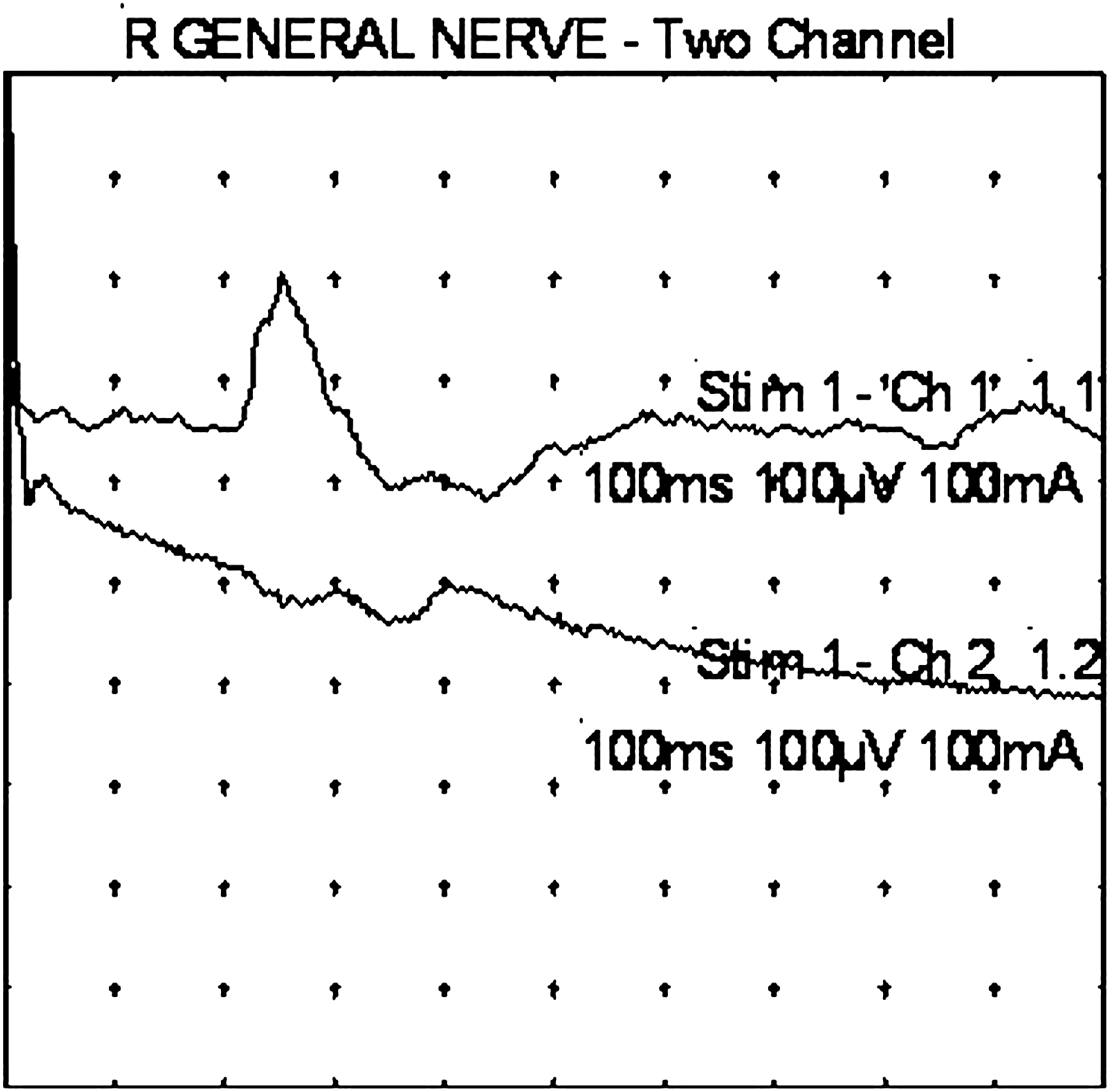
Point of stimulation: center of earlobe (see stimulation setup, in Materials and Methods section of article). Upper trace: Response obtained by recording at the ipsilateral splenius capitis muscle. Initial screening and first identification of an auriculo–cervical reflex. Lower trace: Response obtained simultaneously by recording at the ipsilateral posterior auricular muscle. This might reflect a far field response from the splenius capitis muscle or an inhibitory response at the posterior auricular muscle. R, right.
Results
ACRs were identified on stimulation of the apical helix and lobular regions of the pinnae of the ears in most subjects. These results are shown in Table 1. For each test subject, the presence or absence of ACRs are noted. Short-latency (“T1”), intermediate-latency (“T2”), and relatively long–latency responses (“T3” and “T4”) were variably recorded in these 8 subjects. Their averaged results are depicted in Table 2.
This table notes the results of testing of 8 individual subjects. “T1/V1” refers to latency (in ms)/voltage (in uV) of early onset responses. “T2/V2” similarly refers to intermediate latency responses. “T3/V3” and “T4/V4” similarly refer to late onset responses. The absence of responses is indicated by the dotted lines (-------). “RA” and “LA” refer to the right and left apical helix stimulations. “RL” and “LL” refer to the right and left lobular (earlobe) stimulations. M, male, F, female; Ipsi, ipsilateral; Contra, contralateral.
In this table, the results of individual subject results are averaged. A difference from Table 1 is that each set of latency/voltage data is followed by a fraction, in parentheses, that represents the number of subjects of 8 total in whom these responses were observed. ACR, auriculo–cervical reflex; AVE, average; M, male, F, female; Ipsi, ipsilateral; Contra, contralateral, -------, absence of response.
As raster-mode stimulation revealed that most T1 and T2 ACRs had an initial surface negative deflection, but sometimes had an initial positive deflection (Fig. 2), latencies were measured to the negative peaks, which were relatively more consistent. Where present, the T1 early and T2 intermediate response ACRs tended to have relatively short durations and more uniform surface-negative and surface-positive characteristics, and, so, response amplitudes were measured as voltages from the negative peak to the following positive peak (in displays, negative is always shown in an upward direction). T3 and T4 late responses showed a much more variable organization, typically occurring in a complex that might show a sequence of often relatively long-duration positive–negative–positive–negative waves (+−+ −), or of negative–positive–negative–positive (−+− +) waves, often admixed with higher-frequency waves. For this reason and for the sake of simplicity, latencies were determined by the most prominent long-latency negative peaks, and amplitudes were determined by measuring voltages from the most prominent negative to the most prominent positive peak in each T3 and T4 complex event, whether or not the order of waves was −+− +, or +−+ −. Examples of these responses are given in Figures 4 through 8.

Subject 3 (male, age 59). An example of ipsilateral T1, T2, and T3 responses upon six stimulations of the left apical helix. Note analysis time of 100 ms. L, left.
Subject 4 (male, age 62). Stimulation of left lobular region. Upper trace: Contralateral responses. Lower trace: Ipsilateral responses. Twenty responses averaged. Note that, in this example, T2 is more prominent ipsilaterally, and T1 and T3 are more prominent contralaterally. Note analysis time of 100 ms. L, left.
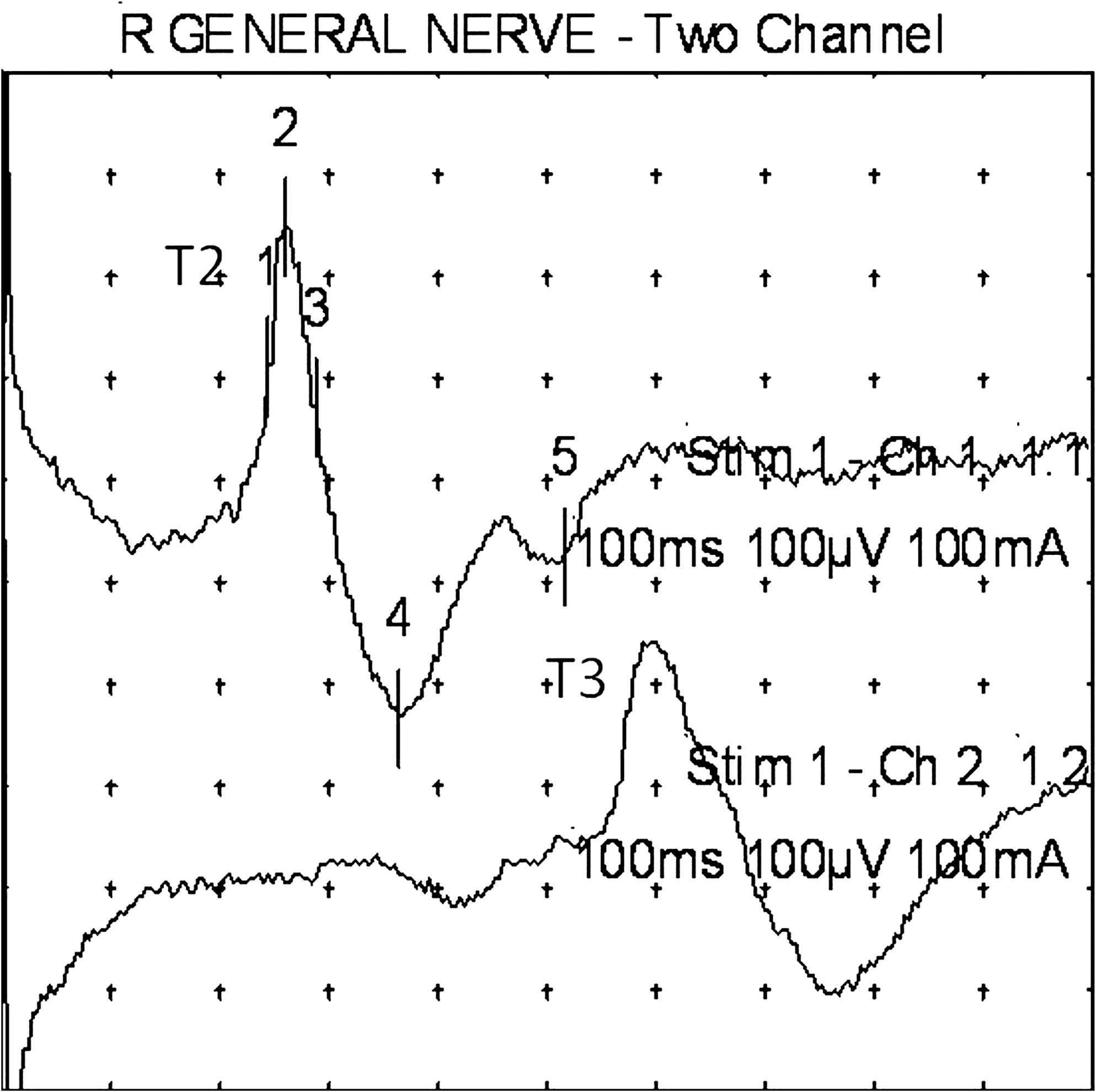
Subject 3 (male, age 59). An example of an ipsilateral T2 response (upper trace), with a contralateral T3 response (lower trace), upon right lobular stimulation. Note analysis time of 100 ms. Note the relatively higher amplitude of these responses. R, right.
Subject 2 (male, age 15). An example of a T3 response. Stimulation of left lobule. Upper trace: Contralateral response. Lower trace: Ipsilateral response. Twenty responses averaged. Note analysis time of 200 ms. The auriculo–cervical reflex here is more prominent contralateral to the ear stimulated. L, left.
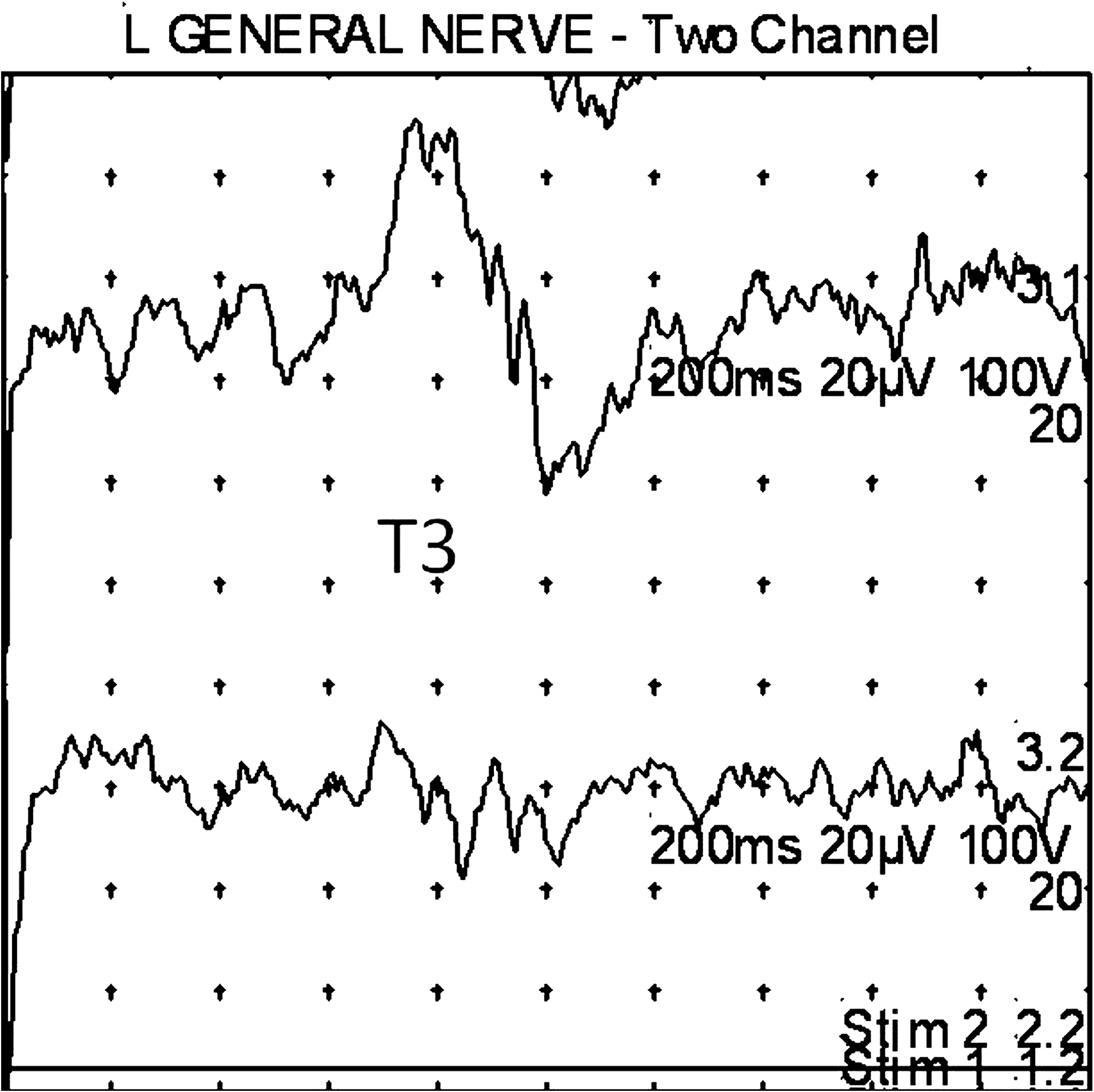
Subject 1 (female, age 70). An example of T3 and T4 complex seen upon stimulation of the right lobular region. Upper trace: Right splenius capitis muscle recording. Lower trace: Left splenius capitis muscle recording. Twenty responses averaged. Note analysis time of 200 ms. R, right.
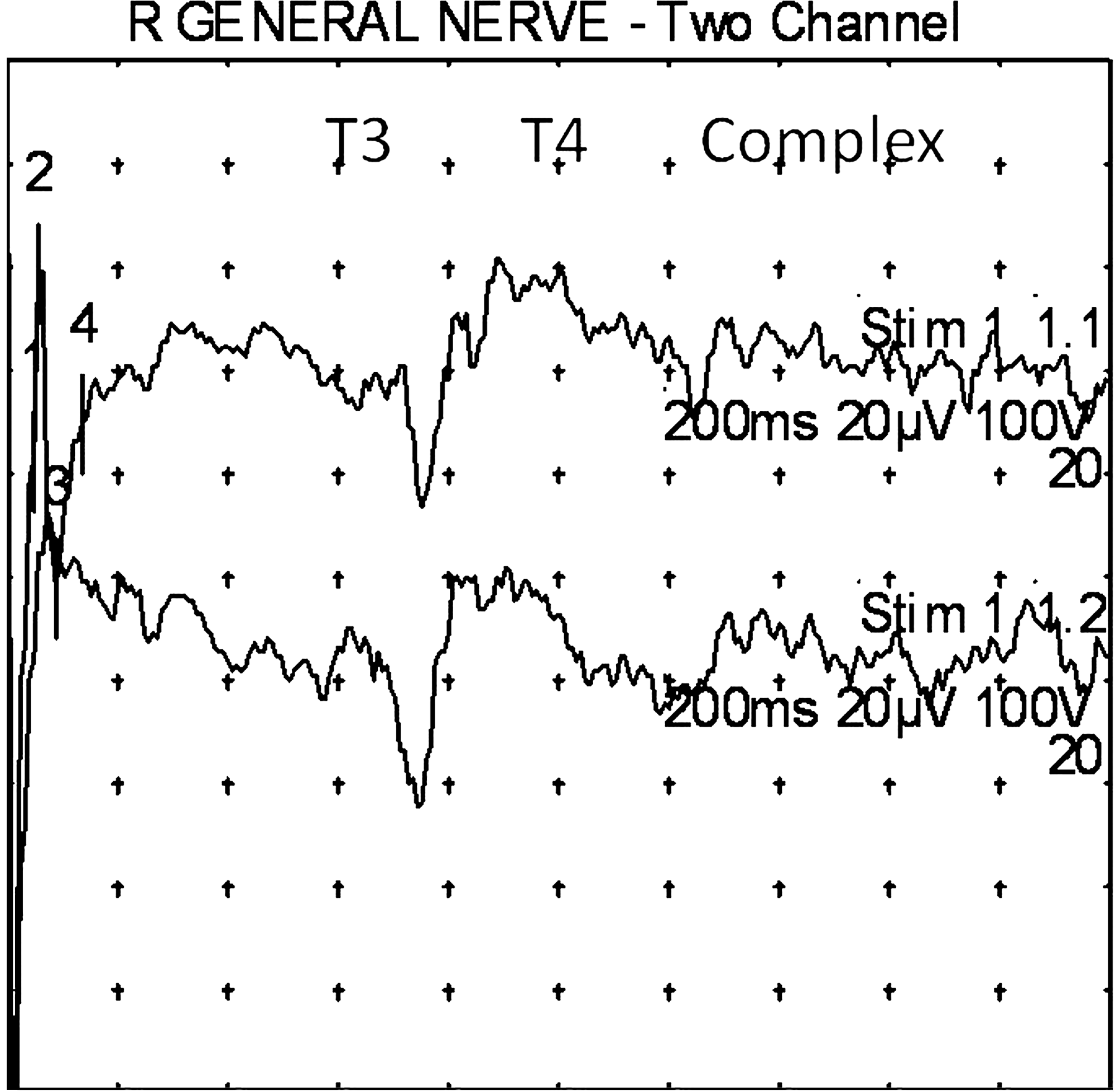
In 1 subject, the author, stimulation was carried out not only at apical and lobular points, but also at conchal points. These results, along with apical helix and lobular stimulation results, are displayed in Figure 9.
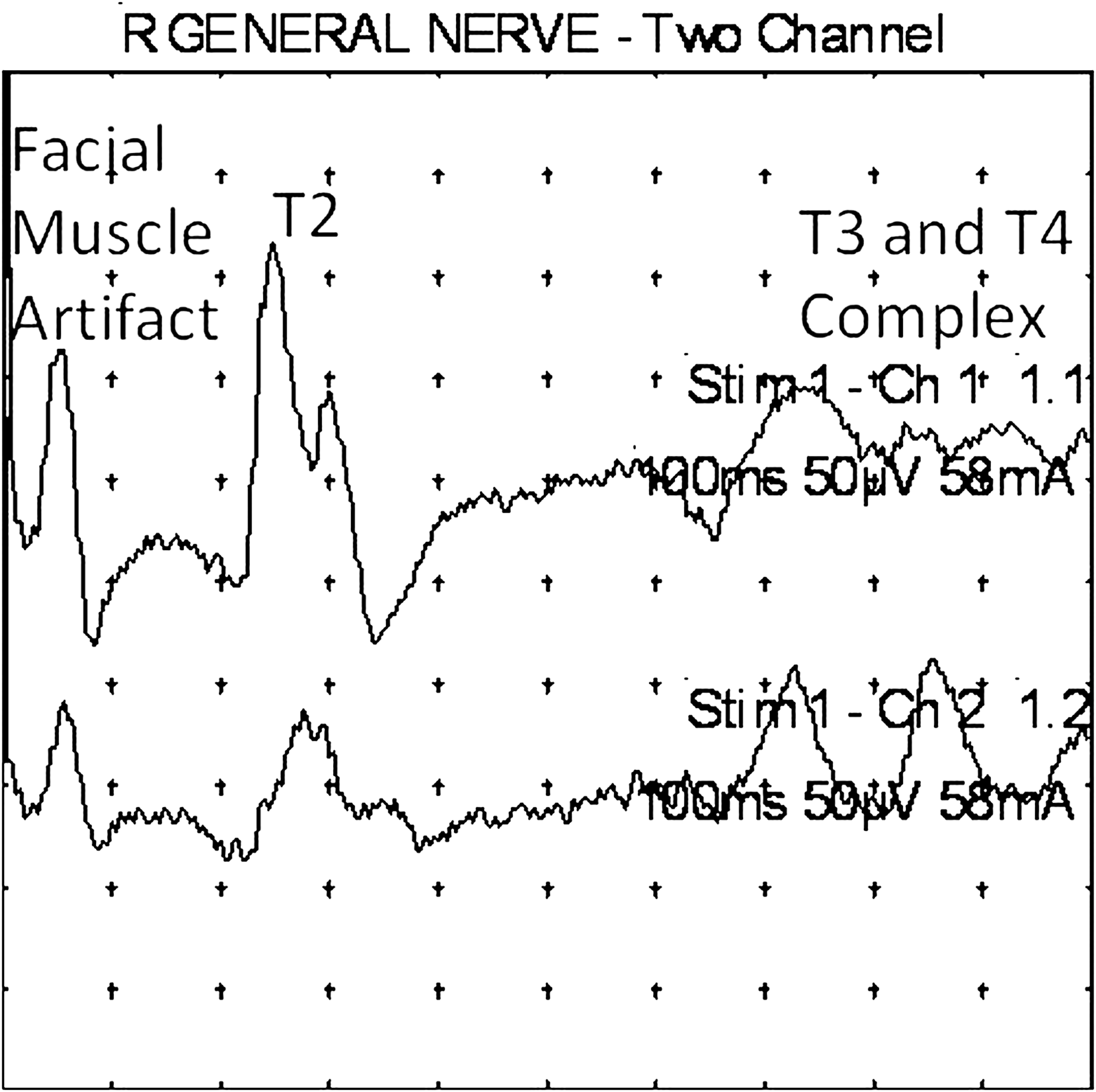
Subject 3 (male, age 59). An example of right conchal stimulation. Note the early facial muscle artifact, which should not be confused with a T1 response. T2 and T3/4 complex responses are evident, but, as the stimulus intensity is increased, a facial muscle artifact appears before the auriculo–cervical reflexes. Upper trace: Ipsilateral response. Lower trace: Contralateral response. Note analysis time of 100 ms. The facial response artifact is seen bilaterally in the suboccipital regions, and is greater ipsilaterally. R, right.
For illustrative purposes, Figure 10 and Figure 11 are included to provide an overall appreciation of right–left differences. In Figure 10, the recordings from 1 subject with particularly robust T1, T2, T3, and T4 responses are shown. In Figure 11, the author's response to apical, conchal, and lobular stimulations are shown. It is worthwhile to note that no two isolated stimulations ever produced identical ipsilateral or contralateral responses. ACRs may be present or absent upon stimulation of any apical, conchal, or lobular site. Sometimes, responses are more prominent ipsilaterally, sometimes contralaterally, sometimes with shorter latency, sometimes with longer latency. This variability of response characteristics is considered further in the Discussion section of this article.
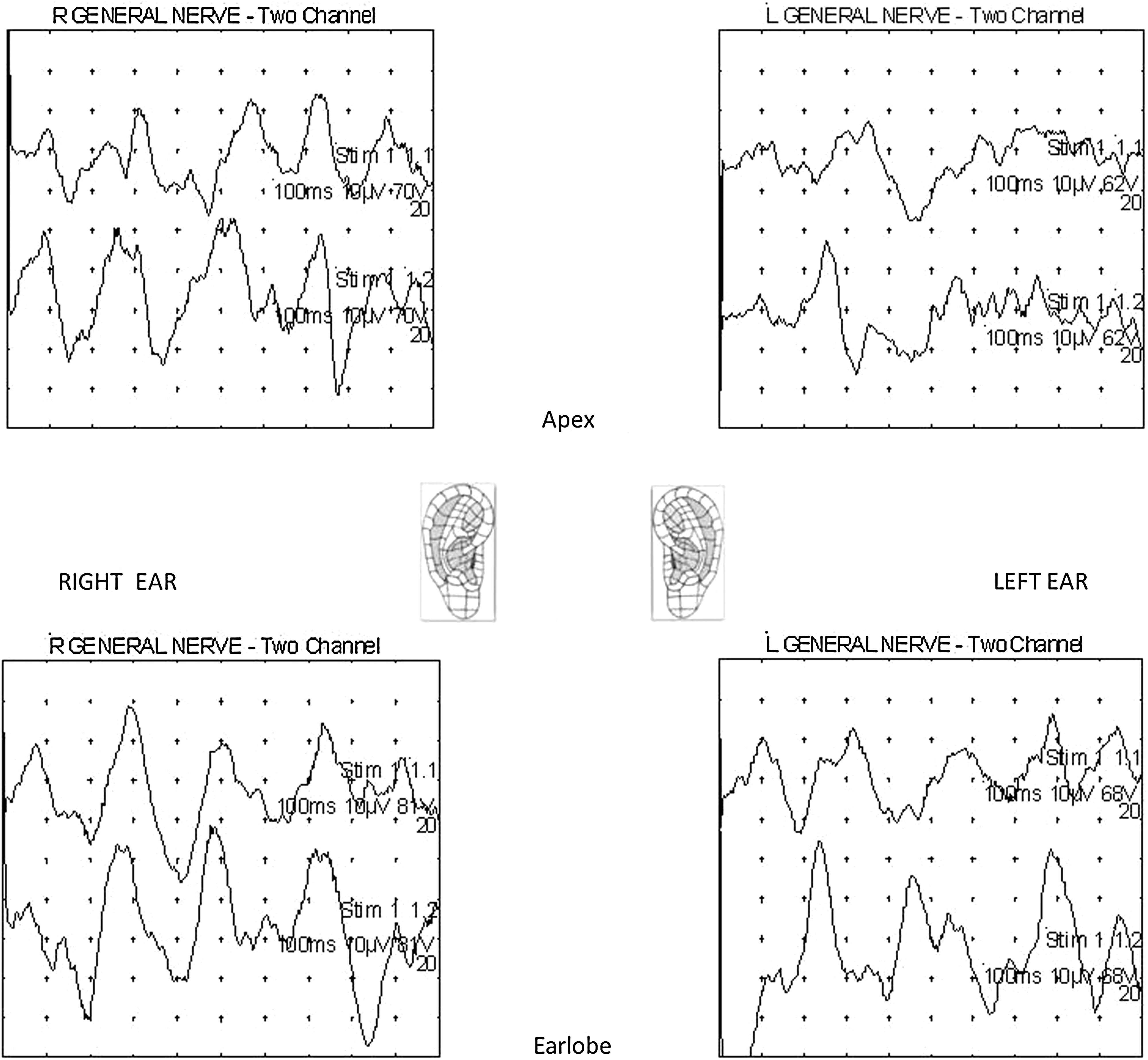
Subject 6 (female, age19). An example of right- and left-sided stimulations of apical helix and lobular points. For right ear stimulations, upper trace responses are ipsilateral, lower trace responses are contralateral. For left ear stimulations, upper trace responses are contralateral, lower trace responses are ipsilateral. Twenty stimulations are averaged in each case. Note analysis time of 100 ms. R, right; L, left.
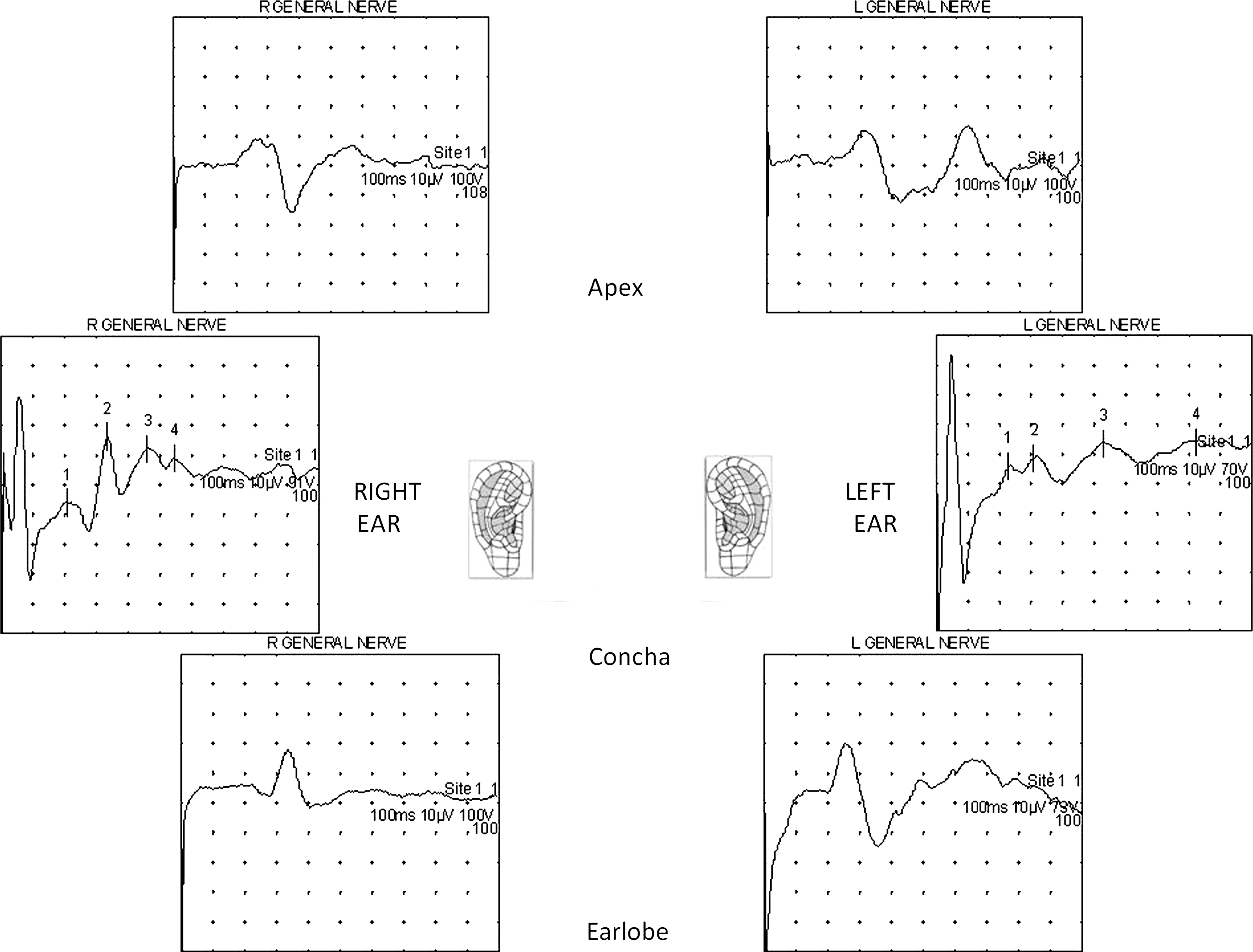
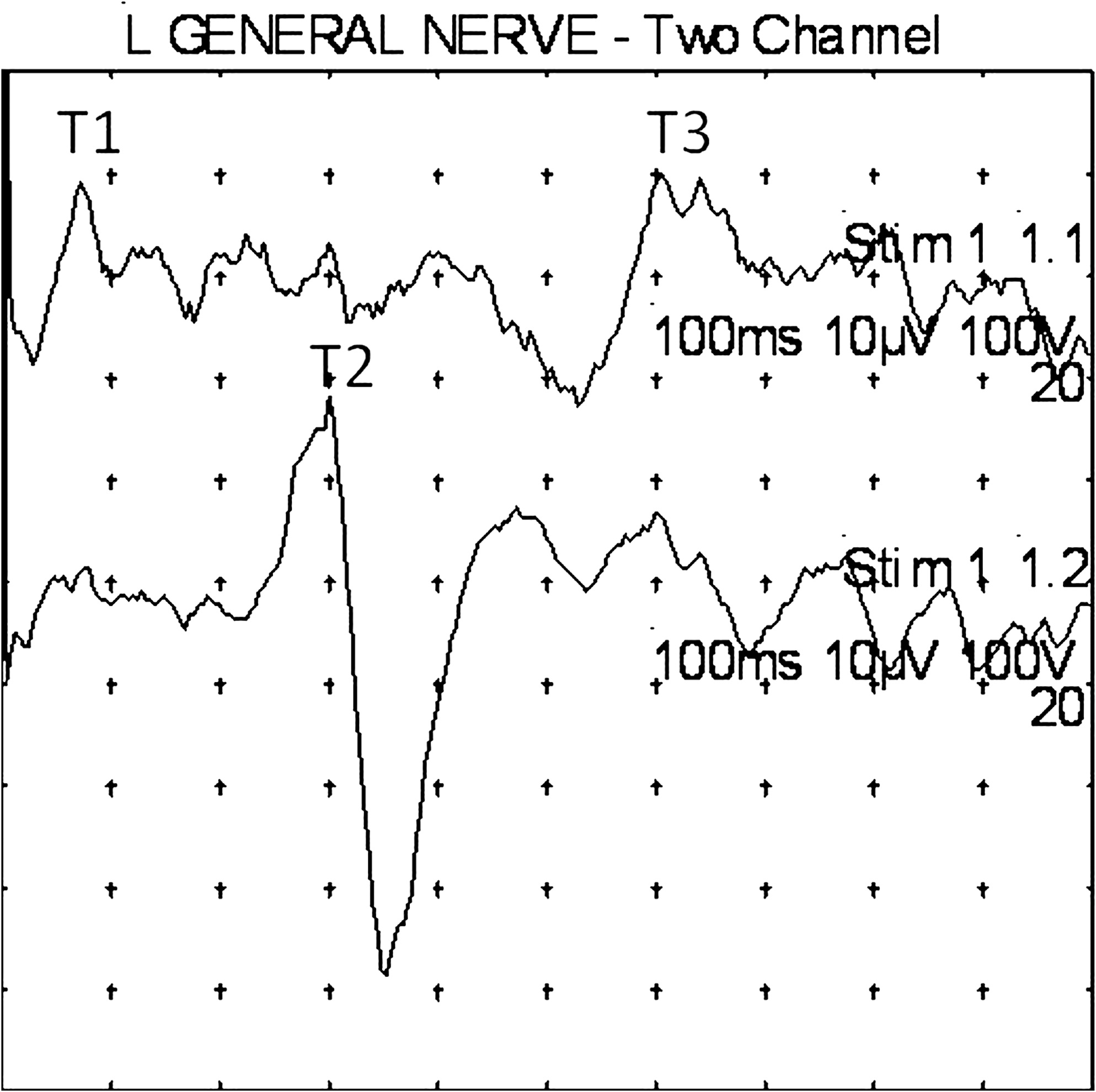
Subject 3 (male, age 59). Another example of of right- and left-sided stimulations. In this example, apical helix (
As shown in Figure 2, stimulation was carried out on the right earlobe of the author, while recording was done simultaneously on the ipsilateral suboccipital and posterior auricular muscles. The averaged suboccipital response was robust and surface-negative; the posterior auricular response was relatively small and surface-positive. Stimulations of the earlobe were also carried out while recording sequentially in the inferior cervical, mid-thoracic, and lumbar paraspinal regions. ACRs were only detected on recording in the suboccipital region, and not more inferiorly.
All ACRs noted were subclinical. That is, no clinical movement of the neck or head was ever seen.
Discussion
This investigation has confirmed the existence of a somatic, EMG human ACR. It is produced by electrostimulation of the skin of the auricle of the ear, and can be recorded over dorsal cervical suboccipital muscles. The ACR described in this report is not an auriculo–visceral or an auriculo–autonomic reflex.
The ACR is not dependent on the auditory function of the ear, but rather on the cutaneous sensory functions of various regions of the auricle of the ear. Mildly painful electrical stimulation of the apical helix and lobular regions of the ears readily induced reflexive subclinical motor responses in the suboccipital region, as recorded over the splenius capitis muscles, but these were not seen while recording over lower cervical, thoracic, or lumbar paraspinal muscles ipsilaterally. The appearance of ACRs was facilitated by loading the splenius capitis muscle, which was effected by asking the seated subject to gently flex his or her neck forward. The ACR could also be facilitated by mildly extending the flexed neck against resistance. Stimulation of an ear most often provoked bilaterally synchronous or somewhat asynchronous subclinical motor responses in the suboccipital region. The appearance of these responses tended to be similar, though never identical, regardless of whether the apical helix or lobular regions of the ear were stimulated. In 1 subject, the author, stimulation of the concha of the ear also produced suboccipital responses similar in appearance to those provoked by apical helix or lobular stimulation. The similarity of suboccipital responses suggests that sensory information related to V3, X, or SCP pathways are conveyed to a common central nervous system (CNS) target, which then, through its synaptic relationships, triggers a motor output to the suboccipital muscles (such as the splenius capitis muscle) to effect the subclinical reflex.
ACRs did have some variability with respect to waveform latency, morphology, and amplitude. For example, although the onset of a response tended to be surface-negative, sometimes this would be surface-positive. Mixed excitatory and inhibitory activity within the (presumably brainstem and superior cervical spinal cord) target effector zones would explain the variability of ACR waveform morphology and amplitude. When 20 ACRs were averaged, the typical onset of a response would be surface-negative. Surface-positive activity suggests inhibition and surface-negative activity suggests excitation. Thus the average initial activity of suboccipital motor responses was excitatory. As mentioned above, pre-existent recruitment of splenius capitis muscle activity facilitated the appearance of ACRs. Therefore, these ACR waveforms most often were likely to represent the product of various underlying excitatory and inhibitory interneuronal and anterior horn-cell motor activities triggered by the auricular cutaneous sensory input ultimately into a partially depolarized superior cervical motor pool. It is noteworthy that if test subjects were seated with their heads erect, relatively unloading the splenius capitis muscles, the ACRs were often more difficult to obtain.
Not only did the latency vary somewhat for averaged ACR waveforms, but, in addition, groups of ACR waveforms were identified at relatively short, intermediate, and long latencies. The short-latency waveforms (T1) had a peak latency typically of ∼9 ms, associated with an average amplitude of ∼21 uV. Such waveforms occurred in 13% of subjects tested. The intermediate waveforms (T2) had a peak latency typically of ∼27 ms, associated with an average amplitude of ∼23 uV. Such waveforms occurred in 63% of subjects tested. The long-latency waveforms typically occurred in a complex and were divided into two groups (T3 and T4). The average peak latency of the relatively earlier long-latency waveform group (T3) was ∼79 ms, with an average amplitude of ∼33 uV, occurring in 88% of subjects tested. The average peak latency of the relatively later long-latency waveform group (T4) was ∼134 ms, with an average amplitude of ∼23 uV, occurring in 43% of subjects tested.
In general, the most impressive responses seen involved the T2 intermediate-latency waveforms, but the most commonly evoked waveforms, and, in fact, those of generally greater amplitude, were of longer T3 and T4 latencies that tended to occur as a complex. The least commonly evoked waveforms, also of generally lowest amplitude, were the short-latency T1 responses.
Inspection of these waveforms confirms the existence of a somatic ACR, despite the small number of subjects tested. Obviously, the same small number of subjects greatly limits more substantial analysis of these results, and so an analysis of their statistical significance was not carried out.
Of note, facial-nerve contamination, routinely seen with conchal stimulation, and infrequently with lobular stimulation, was seen at latencies typically of ∼5 ms. Facial-nerve contamination resulted in clinical ipsilateral facial twitching. Otherwise, all ACR responses in the neck were subclinical, producing no neck or head movement. In the case of the recording of a posterior auricular response (Fig. 3), its relatively low amplitude and largely surface-positive nature suggests that it may have been a far field response manifestation of the synchronous, robust and primarily surface-negative ACR.
Despite the small number of subjects tested, certain speculative trends of the results are suggested, but are as yet unconfirmed. These and the overall analysis results are summarized in Table 3. The relatively small number of T1 responses precludes further analysis. T1 waveforms are probably of disynaptic origin. With respect to the intermediate T2 responses, which is likely to be of oligosynaptic origin, there is a suggestion that latencies obtained by stimulating the earlobe are less than those obtained by stimulating the apical helix and that the evoked amplitudes are greater upon lobular stimulation than with apical helix stimulation. In addition, there is a suggestion that evoked amplitudes tend to be greater ipsilaterally than contralaterally upon stimulation of apical helix or lobular sites. The T3 and T4 late complex responses are probably of polysynaptic origin. There is a suggestion that, in both cases, the latencies obtained are less with right-sided stimulation. There is also a suggestion that response latencies are less for T4 responses contralaterally as opposed to ipsilaterally upon apical or lobular stimulation. Such asymmetries are worth considering for neurologists and auriculotherapists, given the somatotopic organization of the CNS, and its lateralized neurophysiological functions.
Occurrence refers to the % of subjects in which these responses were seen.
Average latency refers to all ipsilateral (Ipsi) and contralateral (Contra) responses.
Average amplitude refers to all Ipsi and Contra responses.
Apical v. Lobular refers to possible trends of latency (Lat) and amplitude (Amp) differences in apical helix (A) versus earlobe (L) responses.
Right v. Left” refers to possible lateralized trends toward Lat differences; no clear trends toward amplitude differences were noted.
Ipsi v. Contra refers to possible trends of differences in Amp and Lat of Ipsi versus Contra responses upon stimulation of specific apical helix or lobular points. v, versus; Stim, stimulation, -------, absence of response.
It seems likely that certain afferent fibers from V3, X, and SCP converge in the trigeminal nucleus. The demonstration of short-, intermediate-, and long-latency ACRs in this study then suggests the possibility that such reflexes are effected through the spinal nucleus of V via disynaptic, oligosynaptic, and polysynaptic pathways, respectively. The output of such reflexes, ultimately, is likely to involve synapses with anterior-horn cells of the superior cervical spinal cord, which then provides motor innervation to the splenius capitis muscles of the suboccipital region.
There is reason to believe that ACRs may be produced independently by stimulation of specific V3 (apical helix), SCP (lobular), and X (conchal) sensory receptor targets, based on this study.
The ACR appears to be distinct from the trigemino–cervical reflex. Although the ACR is produced by V3 stimulation at the ear apex, it is also produced by X and SCP auricular stimulation at the conchal and lobular regions, respectively. Perhaps, in a broad sense, one may say that the ACR is a type of a trigemino–cervical reflex, but the ACR is also a form of a vago–cervical and of a superficial cervical plexus–cervical reflex. Even to say that the ACR is a type of a trigemino–cervical reflex is somewhat doubtful, as the method used to evoke the ACR is different from that used to evoke the trigemino–cervical reflex (as described in relevant published trigemino–cervical neurophysiological literature, referenced above). Altogether, these observations suggest that the ACR is not the same as the trigemino–cervical reflex.
The ACR is also clearly distinct from the cat trial's greater auricular nerve–cervical reflex. 6 That reflex was evoked by direct stimulation of the greater auricular nerve intraoperatively, whereas the ACR is obtained by electrostimulation of the auricle of the ear in healthy, awake human subjects. The ACR is a subclinical EMG reflex, which is distinct from behavioral locomotive 14 or hindlimb scratching 15 reflexes seen in certain cat trials on mechanical auricular stimulation. The subclinical ACR is a somatic reflex—not an autonomic reflex such as those reported in anesthesized laboratory rats, 5 in human subjects,16,17 or regarding Snellen's reflex. 12
The clinical significance of the ACR is uncertain. The small number of subjects in this study precludes firm conclusions regarding the absence, presence, symmetry, asymmetry, morphology, or latency characteristics of the ACR in normal subjects.
Using the trigemino–cervical reflex as an analogy, one might postulate that the ACR participates in withdrawal of the head and neck to noxious stimulation of the ear. The trigemino–cervical reflex has been suspected as a contributor to the withdrawal of the face from noxious trigeminal stimulation. Considering speculation by Di Lazzaro et al. regarding the trigemino–cervical reflex, one might say that it is uncertain that ACR “responses are functionally useful. They produce no visible movement of the head in the present experiments. It may therefore be that under normal circumstances they interact with later events to produce a purposeful withdrawal movement.” 7
Similarly, ACRs produced no visible movement of the head in the trials reported in this article, and possibly, during normal behavior, interact with later events to produce a purposeful withdrawal movement.
Beyond its theoretical interest to the field of auriculotherapy as a reflexotherapy, the demonstration of the human ACR may have clinical neurological implications. This is because abnormalities of the trigemino–cervical reflex have been reported in various clinical settings.18–20 Perhaps unilateral absence of the ACR may represent underlying disease of the central or peripheral nervous systems. Such a conclusion will require ACR studies in greater numbers of normal subjects and patients.
Conclusions
This study has documented the existence of a subclinical, electromyographic, somatic human ACR. This dorsal cervical skeletal muscle reflex is distinct from auriculo–visceral or auriculo–autonomic reflexes, from acoustic and non-acoustic clinical behavioral pinna reflexes, and from Snellen's auriculocervical reflex. It is similar to, but different from, the trigemino–cervical reflex. The existence of the human ACR provides neurophysiological evidence that electrical sensory stimulation of the auricle of the ear produces somatic reflexes at a site removed from the ear, lending further support to reflexive theories that aim to explain the mechanism of action of auriculotherapy.
Footnotes
Acknowledgments
The author wishes to thank David Alimi, MD, of the Faculty of Medicine of the University of Paris XIII (France), for his inspiration and instruction, and for his review of this research project. This project was originally submitted to the University of Paris XIII as a Mémoire toward the Diplôme Inter-Universitaire (DIU) d'Acupuncture Auriculaire. The originial manuscript, dated December 20, 2010 was edited for publication.
Disclosure Statement
No competing financial interests exist.