
Abstract
A high-throughput chemiluminescence and ELISA-based biochemical assay to identify mTORC1/mTORC2 kinase inhibitors is described. These mTOR complexes were isolated from HeLa whole cell lysate using mTOR antibodies and in-well immunoprecipitation. The integrity and purity of the mTORC1 and mTORC2 immunocomplexes were confirmed by western blotting. Full-length recombinant 4E-BP1 was used as a substrate and the catalytic activity was measured by detection of p4E-BP1 [T37/46] by a chemiluminescence method. The performance of this assay that can be used to identify dual mTORC1 and mTORC2 kinase inhibitors in a high-throughput 384-well format is described.
INTRODUCTION
The mammalian target of rapamycin (mTOR) is a Ser/Thr protein kinase that is involved in cell growth and survival through control of cap-dependent translation of mRNA. mTOR belongs to the PI3K-related kinase (PIKK) family and is activated by growth factors, nutritional signals, and energy supply. It exists in 2 distinct complexes described as mTORC1 (ternary complex of mTOR, raptor, and GβL) and mTORC2 (quaternary complex of mTOR, rictor, GβL, and Sin1).1 mTORC1 is involved in phosphorylation of S6K1 and 4E-BP1, which are linked to protein synthesis. mTORC1 has been shown to phosphorylate S6K1 at T389 and 4E-BP1 at T70 and S65.2 , 3 On the other hand, mTORC2 has been shown to phosphorylate a survival kinase, AKT at S473 in vitro and in vivo.4 In addition, it has been demonstrated that mTORC2 is involved in 4E-BP1 phosphorylation.5
New studies have linked mTOR to several human diseases including cancer, diabetes, obesity, cardiovascular diseases, and neurological diseases, and as a consequence, drugs that target mTOR have therapeutic value.6 The diseases most clearly associated with mTORC1 are tuberous sclerosis complex (TSC) and lymphangioleiomyomatosis (LAM), both of which are caused by mutations in TSC1 or TSC2 tumor suppressors.7 Patients with TSC mutations develop benign tumors. Both mTORC1 and mTORC2 have roles in sporadic cancers.8 Inactivation of several tumor suppressors, particularly, PTEN, p53, VHL, and NF1 has been linked to mTORC1 and/or mTORC2 activation. Rapamycin and its analogs (CCI-779, RAD001, and AP23573) inhibit mTORC1 and have shown modest anticancer activity.9 Because of the existence of negative signal from S6K1 to the insulin/PI3K/AKT pathway, it is important to note that inhibitors of mTORC1 such as rapalogs can activate Akt. If this effect persists with chronic rapamycin treatment, it could provide cancer cells with an increased survival signal that may be clinically undesirable.10
On the other hand, mTORC2 has been shown to activate Akt. Akt is activated in many cancers and it regulates cell survival, cell proliferation, and metabolism by phosphorylating proteins such as BAD, FOXO, NF-κB, p21Cip1, p27Kip1, GSK3β, and others. AKT might also promote cell growth by phosphorylating TSC2. AKT activation probably promotes cellular transformation and resistance to apoptosis by collectively promoting growth, proliferation, and survival while inhibiting apoptotic pathways. Clearly, an mTORC1 and mTORC2 inhibitor might be beneficial for treatment of tumors with elevated AKT phosphorylation. Ideally, such an inhibitor might down-regulate cell growth, cell survival, and cell proliferation and could be used in combination with other chemotherapeutic or MTT agents.
Recently, Toral-Barza et al. have reported on the feasibility of screening for mTOR inhibitors using recombinant mTOR expressed in HEK293 cells.11 Although they have reported significant activity with recombinant full-length and truncated mTOR enzyme, others have been unable to generate recombinant enzyme that is active in vitro. The literature suggests that mTOR exists as a protein complex in vivo and adaptor proteins are necessary for its catalytic activity. Therefore, we have developed a high-throughput immunoprecipitation (IP) kinase assay in a 384-well format for screening mTORC1/mTORC2 inhibitors. This assay is suitable for higher throughput screens similar to the assay described by Toral-Barza et al.,11 but it uses native enzyme complex.
MATERIALS AND METHODS
Chemicals
All general chemical reagents used for buffers and assays were purchased from the Sigma Chemical Company (St. Louis, MO) unless otherwise specified. The mTOR antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Protein G-coated and Ni-chelate plates were obtained from Pierce (Rockford, IL) or BD Bioscience (Franklin Lakes, NJ). HeLa cell pellets (25 g each) were purchased from Paragon Bioservices (Baltimore, MD). PHAS-I (4E-BP1) was obtained from Stratagene (La Jolla, CA). Phospho-4E-BP1 [T37/43] antibody was purchased from Cell Signaling Technology (Beverly, MA). Anti-rabbit HRP antibodies were from Amersham (Livingston, NJ) and the chemiluminescence detection reagents were from Pierce. LY294002 was obtained from Calbiochem (San Diego, CA).
Preparation of HeLa cell lysate to preserve the integrity of mTOR complexes
To prepare whole cell lysates from HeLa cells, 25 g cell pellet was lysed in 60 mL of ice-cold buffer A [40 mM HEPES (pH 7.5), 120 mM NaCl, 1 mM EDTA, 10 mM sodium pyrophosphate, 10 mM glycerophosphate, 50 mM NaF, 0.5 mM orthovanadate, and EDTA-free protease inhibitors (Roche) containing 0.3% CHAPS] for 30 min on a magnetic stirrer in a cold room. After clearing of the lysates by centrifugation at 13,000g for 10 min, the lysate was frozen at −70°C until use.
Immunoprecipitation of mTOR complexes in 384-well plates and the Chemiluminescent ELISA
Protein G-coated 384-well plates were incubated with 0.25 µg of mTOR (a.k.a. FRAP) antibody in 15 µL of buffer A for 1 h at 4°C. To each well, 40 µg of HeLa cell lysate in 15 µL of buffer A was added and incubated overnight at 4°C to immunoprecipitate mTOR complexes. Plates were washed 3 times with buffer A and twice with immunoprecipitation wash buffer [Buffer B: 50 mM HEPES (pH 7.5) and 150 mM NaCl]. Each compound from our compound library was added at 10 µM concentration to each well and DMSO was added to the control wells. The reaction was started by adding 150 ng of His-tagged 4E-BP1 as a substrate in the presence or absence of 100 µM ATP to each well in 25 µL of freshly prepared kinase buffer [Buffer C: 20 mM HEPES (pH 7.5), 10 mM MgCl2, 4 mM MnCl2, 10 mM β-mercaptoethanol, and 200 µM vanadate] and incubated at room temperature (RT) for 30 min. The reaction was stopped by transferring 25 µL of reaction mixture from each well to corresponding wells of fresh Ni-chelate-coated plates and incubated overnight at 4°C followed by 2 h at 37°C. To detect phosphorylation of 4E-BP1, the plates were washed once with TBST (Tris-buffered saline containing 0.1% Tween-20) containing 5% skim milk powder. To each well, 25 µL of 1:1,000 diluted phospho-4E-BP1 antibodies in TBST containing 5% skim milk were added and incubated for 1 h at RT. The plates were washed once with TBST and then 25 µL of anti-rabbit HRP (diluted 1:10,000) in TBST containing 5% skim milk was added. The plates were incubated for 1 h at RT and washed 5 times with TBST. For detection of phospho-4E-BP1, 25 µL of chemiluminescent reagents A+B was added and chemiluminescence was measured using an Analyst® plate reader.
RESULTS
Integrity, catalytic activity, and specificity of mTOR complexes
The antibodies used to pull down the mTORC1 and mTORC2 complexes from HeLa cells are schematically shown in Fig. 1A. The integrity of the immunocomplexes pulled down by N-terminal (N-17) mTOR antibody is shown in Fig. 1B. The presence of mTOR, rictor, raptor, and GβL in the immunocomplexes pulled by mTOR antibody suggests that it contains both mTORC1 and mTORC2 complexes. We have also confirmed the presence of both rictor and raptor complexes in HeLa cells by isolating mTORC1 and mTORC2 complexes separately (Fig. 1B). The mTORC1/mTORC2 complex isolated using the mTOR antibodies was catalytically active since it phosphorylated His-tagged 4E-BP1 as shown in Fig. 1C. The catalytic activity was present only in the presence of ATP and is significantly inhibited by 2 µM of a known inhibitor of mTOR kinase, LY294002. Subsequently, the catalytic activity of the mTORC1 and mTORC2 complexes was determined using 4E-BP1 and AKT as specific substrates, respectively. As shown in Fig. 1C, OXA-01 (1 µM), a selective mTOR kinase inhibitor, significantly inhibited both mTORC1 and mTORC2 catalytic activities.
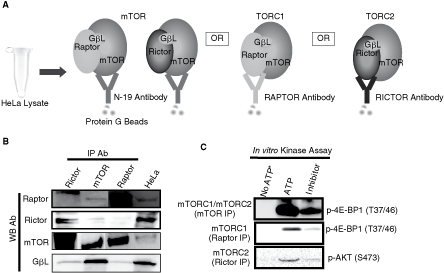
Integrity, catalytic activity, and specificity of mTOR complexes. (
Development of a sensitive and high-throughput chemiluminescence and ELISA-based biochemical assay
We have developed a chemiluminescence-based ELISA assay to measure 4E-BP1 (T37/46) phosphorylation by the mTORC1/mTORC2 complexes in a 384-well plate as illustrated in Fig. 2. In this assay, the mTORC1/mTORC2 complexes were immunocaptured in protein G-coated 384-well plates and used as source of enzyme. As shown in Fig. 3A, phosphorylation of 4E-BP1 (T37/46) was linear with varying amounts of HeLa lysate (ie, enzyme concentration) used for the immunoprecipitation of mTORC1/mTORC2. The phosphorylation of 4E-BP1 (T37/46) was also linear with time as shown in Fig. 3B. The phosphorylation of 4E-BP1 was linear with increasing ATP concentration up to 100 µM. We have determined the K m for ATP for mTORC1/2 complex. The K m for ATP was 40 µM (Fig. 3C). Since this assay was to be used for screening compounds, we also determined the DMSO tolerance of mTORC1/mTORC2 complexes. Up to 3% DMSO was tolerated with a reasonable assay window as shown in Fig. 3D and we have used 2% DMSO in our assays. The assay is very robust and there were no significant issues in optimizing the assay or routine screening.
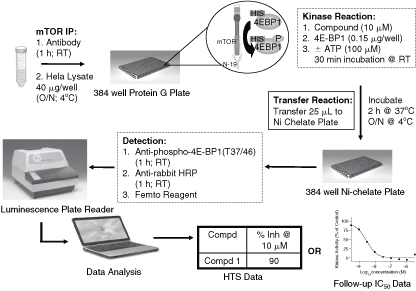
Schematic representation of the 384-well HTS assay protocol.
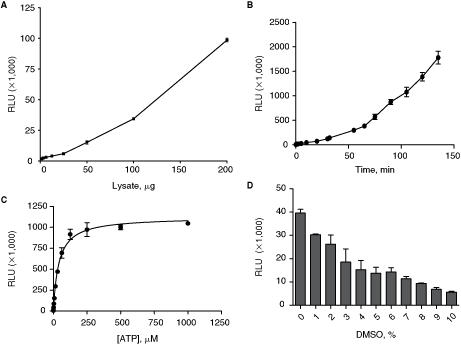
Assay optimization and validation in 384-well plate. Assay optimization with respect to (
Primary screen data
We used the 384-well format assay to screen OSI libraries consisting of thousands of compounds. Primary screen data and follow-up confirmation data for a representative compound are depicted in Table 1. Signal (DMSO control) to background (no ATP) ratio was 50 to 65. Percent inhibition of p4E-BP1 by a representative compound was similar (84% inhibition) in primary and confirmation screens. Surprisingly, the coefficient of variability (CV) was <10% despite multiple steps involved in this assay. The Z/Z′-factor for the assay was 0.18/0.48. Although the Z-factor was considered to be less than ideal, the screen was sufficiently robust permit screening for inhibitors of native mTORC1/mTORC2 kinase.
aDenotes average of 3 values (n = 3).
Z-score and IC50 curve of LY294002, a known mTOR kinase Inhibitor
LY294002 was tested in a dose–response manner as a positive control mTOR inhibitor in this HTS assay. The IC50 curve is depicted in Fig. 4. IC50 value of LY294002 for mTOR kinase was 2 µM, which is similar to published IC50 of 1–1.5 µM using either recombinant or IP mTOR kinase. The Z/Z′-score for the assay was 0.61/0.87.
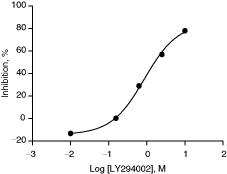
Dose-dependent inhibition of mTOR kinase activity by LY294002. IC50 value of LY294002 was determined in a 384-well format assay as described under Materials and Methods. IC50 value was 2 µM and Z/Z′-score was 0.61/0.87. Z- and Z′-scores were calculated from 352 and 40 wells, respectively. The data presented are average from duplicate wells (n = 2).
Successful screening for mTORC1/mTORC2 inhibitors
We tested several thousands of compounds from our collection in this assay. Compounds were run in a compressed mode, 2 compounds/well, each at a final concentration of 10 µM in 2% DMSO. The pass rate was 72%. More than 50% inhibition at 10 µM compound concentration was considered a hit and the hit rate was reasonable (1.3%). Hits were chosen on a per run basis by either >50% inhibition or 3× St Dev above the mean, whichever was higher. There were 170 confirmed hits and we added additional compounds based on substructure search to determine IC50 values. In the end, there were 213 compounds with IC50 < 10 µM as depicted in Table 2.
aCompounds were run in compressed mode, 2 compounds per well (10 µM each, 2% DMSO).
bFailed compound plates were retested.
cHits were chosen on a per run basis—either ≥50% inhibition, or 3× StDev above the mean, whichever was higher.
dTotal of 337 represents 170 initial confirmed hits + additional compounds that were never tested during confirmation.
Z/Z′-factor is 0.18/0.48. The Z-factor is calculated from 336 compound wells plus 16 background wells for a total of 352 wells. The Z′-factor is calculated from 24 DMSO control wells plus 16 background wells for a total of 40 wells. Staurosporine and DMSO used as a positive and negative controls, respectively.
Reproducible IC50 curves
Raw data for n = 1 and n = 2 IC50 determination for one of the HTS hits is depicted in Fig. 5. There is a clear dose-dependent inhibition of enzyme activity over 6 concentrations tested. Reproducible IC50 of 1.1 µM, Hill coefficient of 1.1–1.3, and r 2 of 1.0 suggest the robustness and reproducibility of the assay.
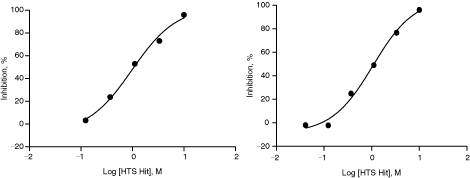
An example of reproducible IC50 determination of a confirmed hit. A confirmed hit was tested for n = 1 (left) and n = 2 (right) in this assay. The IC50 values were same for n = 1 and n = 2 (1.1 µM). The data presented are average from duplicate wells (n = 2).
OXA-01, a selective mTORC1/mTORC2 inhibitor, equipotently inhibits both mTORC1 and mTORC2 complexes
OXA-01, a selective mTORC1/mTORC2 kinase inhibitor, was optimized from the hits identified using this assay. As shown in Fig. 6A, OXA-01 inhibited phosphorylation of 4E-BP1 by mTORC1 in a dose-dependent manner with an IC50 value of 0.029 µM. In addition, OXA-01 inhibited phosphorylation of Akt in a dose-dependent manner with an IC50 value of 0.006 µM as shown in Fig. 6B. Together, OXA-01 is an equipotent mTORC1/mTORC2 kinase inhibitor.
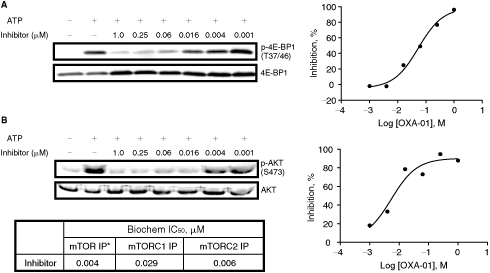
mTORC1 and mTORC2 IC50 values of OXA-01. (
DISCUSSION
The prevalence of PI3K/AKT/mTOR signaling abnormalities in human cancers has suggested the potential use of mTOR kinase inhibitors as novel targeted therapeutic agents. Over the past few years, there has been an interest in developing mTOR kinase inhibitors that inhibit both mTORC1 and mTORC2 activities unlike rapamycin analogs that inhibit only mTORC1 as allosteric inhibitors. Several existing methods for isolation of mTOR complexes employ fresh immunoprecipitation of mTOR from cell lysates. Most assays also require the use of radioactive ATP or western blotting for detection of substrate phosphorylation. These assays are labor intensive and not ideal for screening large compound libraries. Therefore, we have developed the current high-throughput screening assay for identifying dual mTORC1/mTORC2 hits from our compound collection.
This high-throughput screening assay is different from methods reported by others. In this assay, native enzyme complex is used as the source of enzyme rather than using a recombinant or truncated enzyme. It was previously established that mTOR immunocomplex could directly phosphorylate 4E-BP1 at Thr 37/46,2 , 3 , 5 we have confirmed this in our preparation of mTOR complexes by western blotting. Since western blotting is labor intensive and semiquantitative, we have developed this high-throughput screening assay based on ELISA methodology. Assay optimization was achieved by a thorough and appropriate selection of cell line, cell lysis buffer, dose titration of enzyme, substrate, ATP, assay time course, and DMSO tolerance. We observed that HeLa cell lysates were stable when stored in −70°C and thawed no more than once. Enzyme activity was stable after dilution at RT up to 4 h (data not shown). The enzyme reaction was linear up to 200 µg of lysate. We adopted 40 µg of lysate per well in the assay as a suitable compromise between achieving sufficient assay signal and the cost of cell lysate (enzyme). Enzyme activity was linear with time up to 2 h and we have adopted 1 h as an appropriate time point for significant S:B ratio and convenience of use.
We determined the Michaelis constant (K m) values for ATP by incubating mTORC1/mTORC2 complexes with various concentrations of ATP. The apparent K m for ATP was calculated to be around 40 µM. This value for ATP is similar to the value reported by Toral-Barza et al. but significantly lower than a previously reported value of 1 mM ATP.12 Although our result is substantially different from the latter finding, the kinetic parameters of mTORC1/mTORC2 determined in our assays are well within the K m ranges for ATP reported for mTOR and other Ser/Thr kinases.13 Once we had established an optimal chemiluminescence-based high-throughput assay, we assessed the effects of known mTOR inhibitor, LY294002. LY294002 dose dependently inhibited mTORC1/mTORC2 activity with an IC50 value of 2 µM and is similar to IC50 values reported by others.11 More importantly, we have used this method successfully to screen our compound collection and identify mTORC1/mTORC2 inhibitors. OXA-01 is a potent and selective mTORC1/mTORC2 kinase inhibitor that is optimized from these hits and it potently inhibits both mTORC1 and mTORC2 with IC50 values of 29 and 6 nM, respectively. OSI-027, a sister compound of OXA-01, has entered Phase I clinical trial in solid tumors and lymphoma.
Footnotes
ACKNOWLEDGMENTS
We thank Kety Huberman, Chienling Ma, and Anna Chan for technical assistance, and Drs. Lee Arnold and Peter Houghton for helpful discussions.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.