
Abstract
The yeast 2-hybrid (Y2H) system is a powerful method for identifying protein-protein interactions (PPIs), requiring minimal prior information of the putative interactors. Currently available automated versions of the Y2H system are sufficiently developed to allow facile genome-wide PPI screening to compile extensive inter-actome data. A limitation of the Y2H approach, however, is that all primary hits have to be technically verified and biologically evaluated by complementary methods, which is time-consuming, costly, and laborious. Furthermore, the yeast intracellular environment can lead to spurious results for proteins of other organisms, for example because of differences in post-translational modifications or the presence/absence of bridging proteins. Many researchers now confirm PPIs found in the Y2H system by retesting the candidates in the mammalian 2-hybrid system (M2H). However, although such combined Y2H-M2H testing is desirable and perhaps necessary, recloning of Y2H candidates into M2H vectors, especially on a large scale, is time-consuming and costly. To address this shortcoming, we introduce here a new site-specific recombination-capable M2H vector system that is fully compatible with the site-specific Y2H system that we recently described in Biotechniques [2008;45(3):235-244]. The results show that the new vectors are: (a) Gateway®, compatible and suitable for fast cloning; (b) fully functional in the M2H system without influencing the capacity of the selection system or creating autoactivators; and (c) directly compatible with the existing site-specific Y2H vector system, as demonstrated by confirmation of Y2H PPI candidates in the M2H system.
Introduction
Here we introduce a vector system for the M2H system based on site-specific recombinational cloning, 8,9 which is highly efficient, quick, and simple. This new vector system was developed to be used for verification of PPI candidates obtained from Y2H screens, and it combines the advantages of site-specific recombinational cloning and the M2H system. With these vectors, every PPI found in the reading-frame-independent Y2H vector system 2 can be easily verified by retaining the reading frame, without the need to sequence, analyze, redesign, or use restriction enzyme-based cloning.
Materials and Methods
Construction of the Vector System
The 2 M2H vectors pAct and pBind of the CheckMate™ Mammalian 2-Hybrid system (Promega, Madison, WI) were used to incorporate the Gateway® cassettes rfa, rfb, and rfC.1 of the Gateway conversion system (Invitrogen, Carlsbad, CA). For negative selection the cassettes carry the ccdB gene, 10 and resistance to chloramphenicol for positive selection. Blunt-ended linear cassettes were cloned into the EcoRV restriction site of the Multiple Cloning Site of both vectors, which were then used to transform chemically competent ccdB Survival Cells (Invitrogen). Isolated single colonies were cultured and plasmids isolated by using a GenEluteTM HP Plasmid Maxiprep Kit (Sigma, St. Louis, MO). The resulting purified plasmids were sequenced.
p53 and SV40 Large T-antigen Bait and Prey-Plasmid Generation
DNA sequences coding for the mouse p53 and the large T-antigen transcripts, without start and stop codons (to enable C- and N-terminal fusions), were available in the Gateway donor vector pDONR™/Zeo (Invitrogen). 2 Using site-specific LR reactions, 8 the p53 sequence was shuttled into the new pMBD-Gate2 vector and the sequence of the viral oncoprotein large T-antigen of simian virus 40 (SV40) was inserted into the pMAD-Gate2 vector. After transformation into Escherichia coli electrocompetent cells and positive selection against ampicil-lin (100 μg/mL), plasmids from colonies were sequenced. Insert-harboring clones contained cassettes in both the forward and reverse directions.
Mammalian 2-Hybrid Test
To verify the functionality of the new M2H system, p53 in pMBD-Gate2 and SV40 in pMAD-Gate2 were co-transfected with the pG5luc vector of the CheckMateTM M2H system (Promega) into 293 HEK (human embryonic kidney) cells. For this, 1-3 × 105 293 HEK cells were seeded in flasks to reach ∼50°/o-80°/o confluence and electroporated by using a Cell Line Nucleofector® kit V (Amaxa Biosystems, Lonza GmbH, Wuppertal, Germany) according to the manufacturer's instruction with a total of 9 μg plasmid DNA (3 μg each). To detect possible autoactivity of the new vectors, we also transfected both empty pMAD-Gate2 and pMBD-Gate2 vectors together with pG5luc in the same way. In addition, the p53 and SV40 large T-antigen plasmids were each subjected to an autoactivation test by transfecting them with the corresponding empty pMAD-Gate2 or pMBD-Gate2 together with pG5luc. As a positive control, 293 HEK cells were transfected with pAct-MyoD and pBind-Id plasmids (both from Promega) together with pG5luc. The interactions were tested by measuring the activities of Firefly and Renilla luciferase reporters sequentially from each sample by using the Dual-Luciferase Reporter System (Promega): promoter activity values are expressed as arbitrary units after normalization and correction for background reactivity. A PARADIGM Detection Platform (Beckman Coulter Inc., Fullerton, CA) was used for signal detection.
Coupled Y2H-M2H Analysis
A Y2H screen was performed as described by Maier et al. 2 Human full-length Keratin 14 (KRT14) was cloned into pBD-Gate2 and screened against a human 3-frame skin cDNA library. The cDNA library is a Y2H prey library, which was produced by blunt-end cloning of a poly-A primed human skin cDNA into pDONR/Zeo and subsequent LR cloning into a mixture of 3 different Y2H prey-plasmids each having a different reading frame (pAD-Gate1-3). The LR-reaction products were used to transform competent E. coli cells, and 107 independent colonies were obtained, which were subjected to plasmid isolation and library production. A total of 200 μg of the library together with the KRT14 bait was used to transform the Y2H strain AH109 (Clontech, Mountain View, CA) and the cells were plated onto PPI-selective plates (-Leu-Trp-His-Ade synthetic dropout plates). According to the calculated transformation efficiency, we screened 106 bait-prey combinations, which resulted in 3 positive Y2H colonies. We isolated the prey-plasmid from 1 of the 3 colonies and shuttled the candidate insert by 1-tube BP and LR reactions into a mixture of pMAD-Gate1-3 vectors as described in the Gateway cloning instruction manual (Invitrogen). The LR-reaction products were used to transform E. coli cells and propagated as a plasmid mixture. KRT14 was shuttled in the same way into pMBD-Gate2. The candidate clone (plasmid mixture of the candidate clone) was tested directly with KRT14, and additional mock transfections with empty vectors were performed in the Gateway-modified M2H system as described earlier.
Results and Discussion
We created a series of Gateway-compatible M2H vectors specialized for coupled Y2H-M2H analysis. Six different M2H bait and prey vectors were produced ( Fig. 1 ). We termed the modified bait vectors pMBD-Gate1-6 and the prey vectors pMAD-Gate1–6 (the numbers 1 to 6 indicate the different reading frames). All 3 reading frames are inserted in both orientations so that all 6 possible reading frames for correct protein expression are available in the bait and prey M2H vectors. These reading frames are directly compatible with the Y2H vector system. 2 A found Y2H interactor can easily be shuttled into the corresponding M2H bait or prey vector. For convenient handling, the numerical designations of the M2H vectors directly correspond with the numbered Y2H vectors. Thus, for example, a protein cloned in pAD-Gate2 and recovered from a Y2H screen can be shuttled into pMAD-Gate2 while retaining the correct reading frame. We evaluated these vectors by using the strong and well-known PPI between the tumor suppressor p53 protein and the SV40 T-antigen oncoprotein. 11 As shown in Figure 2A, only the p53-SV40 T-antigen transfection as well as the positive control showed a significant increase in reporter values compared with the negative controls.
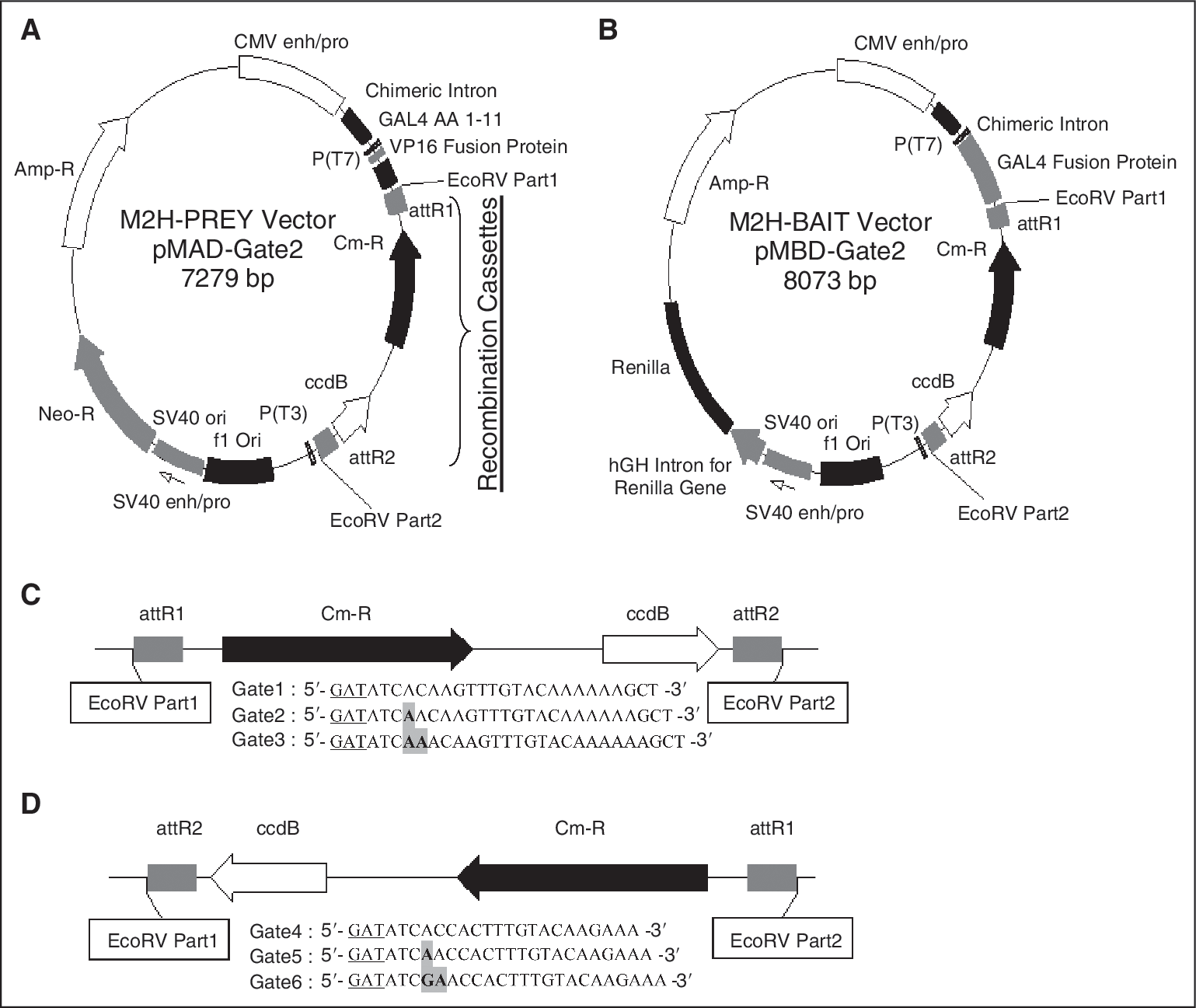
Construction of the new vector system. Illustration and description of the new Gateway®-compatible mammalian 2-hybrid system (M2H) prey vector pMAD-Gate2 (
While there is no requirement for M2H vectors for different reading frames if the genes are specifically directly cloned into an available M2H vector, it can be crucial if clones are isolated in upstream approaches (eg, from a Y2H screen, an isolated/fished partial or out-of-frame cDNA clone). Usually, one has to redesign primers, PCR-amplify (with the possibility of introducing mutations), clone, sequence, and so on before the M2H study can be continued. While this can be done when a few genes are being tested, it will be a rate-limiting and cumbersome task if large sets of genes are analyzed. With the described vectors, the authenticity of the isolated clones from upstream experiments can be saved and shuttled with minimum hands-on steps either to the proper reading-frame M2H vector or, if desired, into all vectors (a pooled cloning) and used in an M2H assay.
Additionally, we evaluated the flexibility and practicability of the new system to verify Y2H PPIs obtained from a genome-wide library screen. For this a Y2H screen was performed. The interaction between the candidate clone and KRT14 was confirmed with the modified M2H system and no autoactivating activity was observed ( Fig. 2B ). Sequencing of the candidate identified a fragment of human Keratin 5 (KRT5), which had already been described as an interactor by forming heterodimers with KRT14, 12 –14 thus reinforcing the reliability of our screen.
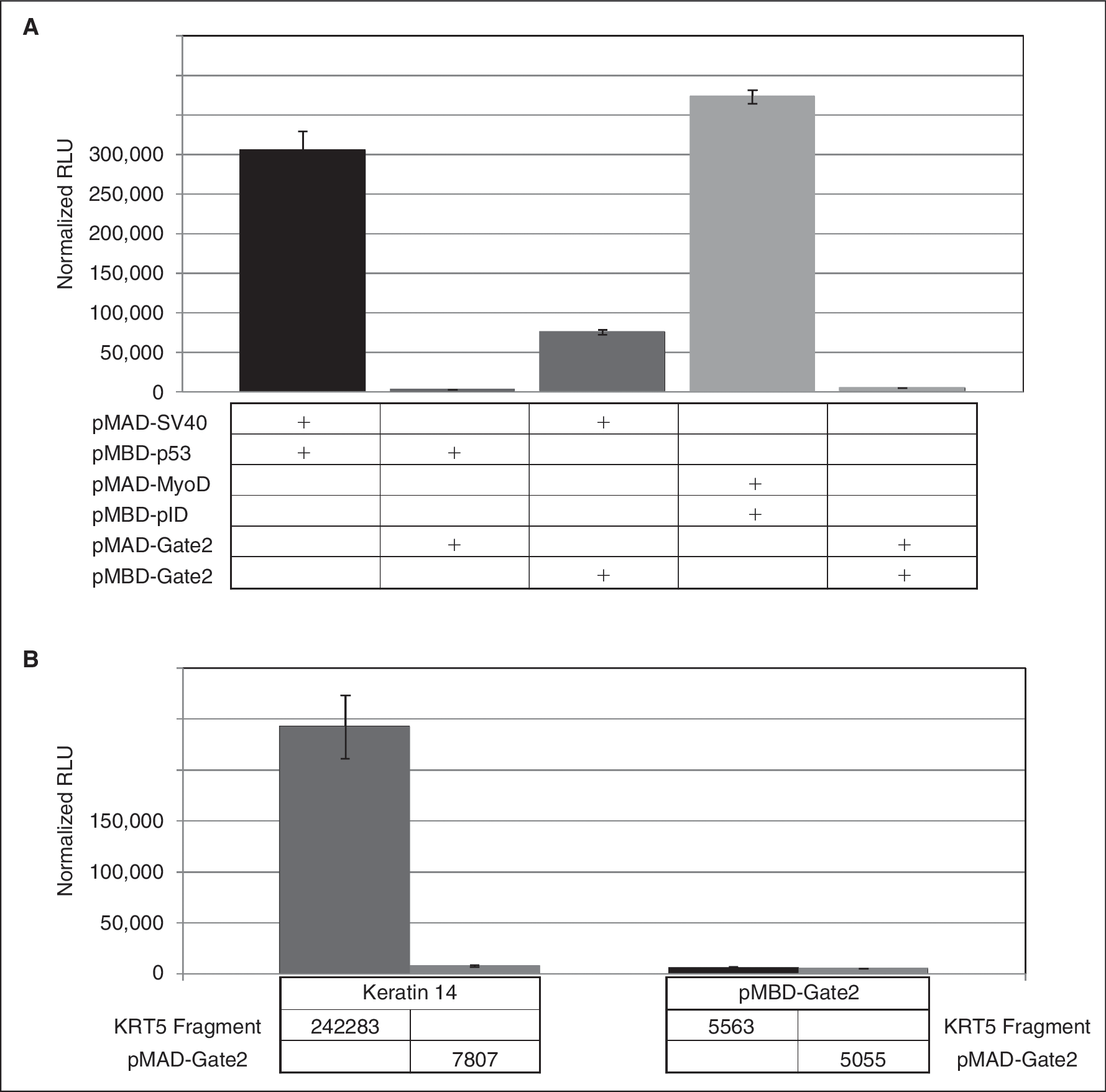
Test of functionality of the modified vectors and evaluation ofa coupled yeast 2-hybrid (Y2H)-M2H approach. (
In summary, we have constructed a Gateway reading-frame-independent vector system for M2H studies for use as an analytic extension to the Y2H assay, for coupled Y2H-M2H PPI hunts, or for stand-alone PPI analysis. The results show that the new vectors are: (a) Gateway-compatible and amenable to site-specific, rapid cloning; (b) fully functional in the M2H system without influencing the capacity of the selection system or creating autoactivators; and (c) directly compatible with the Y2H vector system published by Maier et al. 2 for verification of newly discovered PPIs, as successfully demonstrated by a Y2H-M2H-coupled genome-wide interactor hunt with KRT14. In addition, the use of site-specific recombination-compatible vectors in the M2H system would be useful for interaction studies in view of the rising number of Gateway-compatible resources in entry vectors. Finally, to permit scientific progress, we want to enable any scientist to have unrestricted and easy access to the new vector system. Therefore, the vectors will be deposited in a non-profit plasmid repository (Addgene, Inc., Cambridge, MA).
Footnotes
Author Disclosure Statement
No competing financial interests exist.