
Abstract
By secreting proteases such as cathepsins and matrix metalloproteinases (MMPs), macrophage foam cells may be a major cause of ruptured atherosclerotic plaques. The aims of the present study were to investigate in vitro role of human macrophage foam cells in degrading type I collagen, a major component of extracellular matrix (ECM) in plaques, and to establish whether the pro-inflammatory molecules, tumor necrosis factor (TNF)-alpha, and receptor activator of nuclear factor-κB ligand (RANK-L) increase this degradation. CD14+ monocytes isolated from peripheral blood were differentiated into macrophage foam cells and cultured on a type I collagen matrix in the presence of TNF-alpha and RANK-L. Matrix degradation was measured by the cathepsin K-generated C-terminal cross-linked telopeptide of type I collagen (CTX-I) and the MMP-generated carboxyterminal telopeptide of type I collagen (ICTP) in supernatants showing that macrophage foam cells secrete MMPs and cathepsin K, resulting in release of ICTP and CTX-I. Stimulation with TNF-alpha increased CTX-I and ICTP dose dependently, with ICTP levels increasing by 59% and CTX-I levels increasing by 43%. RANK-L enhanced the release of CTX-I and ICTP by 56% and 72%, respectively. This is, to our knowledge, the first data describing a simple in vitro system in which macrophage foam cells degradation of matrix proteins can be monitored. This degradation can be enhanced by cytokines since TNF-alpha and RANK-L significantly increased the matrix degradation. This in vitro system in part is a model system for the macrophage-mediated proteolytic degradation of the ECM, which is found in many diseases with an inflammatory component.
Introduction
Human atherosclerotic plaques are characterized by a lipid-rich core covered by a fibrous cap composed in part by fibrillar collagens, elastin, and proteoglycans 8,9 of which, collagens are the major component. 10,11 Type I collagen accounts for approximately 60%–70% of total collagen in ECM in the arteries. 12,13 The ECM is critically important for maintaining the physicochemical structure of the artery, providing tensile strength as well as viscoelastic properties. 14 Changes in the composition of the ECM are in part due to elevated levels of proteases produced by macrophages, macrophage foam cells, and other inflammatory cells, which degrade the proteins in the fibrous cap in the plaque. This pathological remodeling may lead to erosion of the fibrous cap and delivery of the highly thrombogenic lipid core to the circulation, resulting in thrombus formation. 15 –17
In atherosclerotic plaques, matrix metalloproteinases (MMPs) have hereto been considered to be the proteases of paramount importance. 18 –21 An array of MMPs is expressed in many stages of the development of the plaque. MMPs are most often secreted by smooth muscle cells, macrophages, and macrophage foam cells bordering the fibrous cap within plaque shoulders. 11,22 The important role of MMPs was also shown by overexpression of the tissue inhibitor of metalloproteinase-1 (TIMP-1) in apolipoprotein E-deficient mice, which was associated with reduced atherosclerotic lesion growth and protection against aneurysm rupture. 23,24 However, recent research has clearly outlined a more complex nature of plaque degradation, in which other proteases such as cathepsins and aggrecanases play equally important roles. 21,25 –28 Cathepsin K is localized in rupture-prone areas such as the fibrous cap, plaque shoulders, and at the actual site of plaque ruptures. 29 –31 It is predominantly secreted by macrophage foam cells and by osteoclasts in the bone. 32 –35 Moreover, disruption of the cathepsin K gene reduces atherosclerosis progression, suggesting the proteolytic activity of cathepsin K to be important for the pathogenesis of atherosclerosis. Mice deficient in both cathepsin K and apolipoprotein E show increased lesion stability, as collagen content in plaque areas and fibrous cap thickness increased. 36 In terms of biochemistry, cathepsin K may be more collagenolytic than MMPs, emphasizing its importance in collagen-rich matrix remodeling. 37
As type I collagen is the main constituent of ECM in the plaque and both collagen-degraders, MMP and cathepsin K, have been implicated in the pathogenesis of atherosclerosis, we investigated this relationship in a cell culture model. Macrophage foam cells were cultured on a matrix of collagen type I and the degradation was measured by the use of two enzyme-linked immunosorbent assays (ELISAs); the C-terminal cross-linked telopeptide of type I collagen (CTX-I) assay is an ELISA specifically recognizing a cathepsin K fragment of type I collagen 38 and the carboxyterminal telopeptide of type I collagen (ICTP) assay is another ELISA measuring an MMP-generated fragment of type I collagen, which has been shown to be degraded by cathepsin K. 37 Neither of these assays react with intact type I collagen, and are specific for a given type of pathological degradation of type I collagen. 38
Inflammation plays an important role in atherosclerosis. Antitumor necrosis factor alpha (anti-TNF-alpha) has been identified as a player in several cardiovascular processes 39 –41 by, for example, being a potent stimulator of MMPs. 42,43 . TNF-alpha has been shown localized in human atheroma and is mainly synthesized by macrophages. 44,45 Large studies with TNF-alpha treatment have shown to reduce systemic and local inflammation in patients with rheumatoid arthritis (RA), reduce disease activity, and lower risk of developing cardiovascular events associated with RA. 46,47 This is compatible with the hypothesis that inflammation contributes to the development of cardiovascular events. This prompted the current investigation into the role of TNF-alpha on macrophage foam cells and, in particular, their ability to release the proteases leading to degradation of ECM components.
Another potent inducer of catabolic enzymes is the receptor activator of nuclear factor-κB ligand (RANK-L), 35 whose signaling is required for osteoclast differentiation, activation, and survival. 48 This cytokine has recently been suggested a role in atherosclerosis, and the underlying mechanism and relationship to osteoporosis are being explored. 49 –51 Investigation of protease secretion and collagen degradation in RANK-L-stimulated human macrophage foam cells is therefore of high interest when studying the role of and mapping molecular mechanisms in atherosclerotic matrix remodeling.
The aims of this study were (1) to develop an in vitro system mimicking the human macrophage foam cell degradation of matrix in atherosclerosis, and (2) to investigate the levels of proteases secreted by human macrophage foam cells, by measuring two specific markers of MMP and cathepsin K-mediated matrix remodeling, ICTP and CTX-I, respectively. In addition, we investigated the effect of TNF-alpha and RANK-L on the remodeling of the ECM.
Experimental Procedures
Cell Culture
Blood from healthy humans was received from the Danish University Hospital in Copenhagen as thrombocyte concentrates (buffy coats). The investigation conforms with the principles outlined in the Declaration of Helsinki. 52 Human monocytes were isolated from the blood by centrifugation on a Ficoll-Paque gradient (Ficoll-Paque™ Plus; GE Healthcare Bio Sciences AB) and magnetically sorted using primary coated CD14+ magnetic beads (Dynabeads M-450; Invitrogen Dynal). The cells were seeded into 150 cm2 flasks containing phenol-red free α-minimum essential media (α-MEM) (MEM ALPHA; Gibco, Invitrogen) added 10% serum (Sigma-Aldrich), 100 units/mL penicillin and 100 μg/mL streptomycin (BioWhittaker), 0.22% thymidin (Applichem), and 25 ng/mL macrophage-colony stimulating factor (M-CSF; R&D Systems) and incubated for 6 days at 37°C, allowing the monocytes to differentiate into macrophages. These macrophages were lifted by trypsin (BioWhittaker) and cultured on either plastic or matrigel, as described in the following.
The macrophages cultured on plastic were cultured in alpha-MEM in a 24-well plate (200,000 cells/well) for 1, 3, 7, or 10 days counting from the time of culture into the wells. Half of the macrophages were incubated in the presence of 50 μg/mL LDL from human plasma (Sigma-Aldrich), allowing the differentiation into macrophage foam cells. Alpha-MEM and LDL were replenished twice a week. After the culture with or without LDL, some cells from each group (macrophages and macrophage foam cells) were fixed in 4% formaldehyde, and stained for lipid droplets by oil red O staining. Some cells from each group were lysed for western blotting. Supernatants were collected for gelatin zymography and for an assay measuring MMP activity.
To partially resemble the arterial wall, a matrix was created, consisting of 48.8% Matrigel(IV) (Matrigel™ matrix; BD Biosciences), 41.5% type I collagen 3 mg/mL (pig collagen A-I; Nitta gelatin), 4.9% 10× alpha-MEM (Invitrogen), and 4.9% reconstitutions buffer (NaHCO3, NaOH, and 1 M HEPES). The components were mixed on ice, distributed into 48-well plates with 250 μL/well, and solidified at 37°C. The plates were then incubated at 37°C with alpha-MEM for 3 days, allowing the penetration of medium before some of the trypsin-lifted macrophages were added (100,000 cells/well) in new alpha-MEM. After allowing the macrophages to be incorporated into the matrix (1 day), the cells were incubated in presence of 50 μg/mL LDL transforming the cells into macrophage foam cells and in presence of different modulators for 1, 3, 7, and 10 days, respectively, counting from the time LDL was added to the macrophages—in other words, from the moment the cells become macrophage foam cells. The modulators used were 1, 10, and 100 ng/mL TNF-alpha (R&D Systems), 1, 10, and 100 ng/mL RANK-L (R&D Systems), 20 μM GM6001 (Sigma-Aldrich), and 20 μM E64 (Sigma-Aldrich). The two inhibitors E64 and GM6001 were added together to eliminate the protease activity of MMP and cathepsin K. The supernatants were collected at days 1, 3, 7, and 10 (counting from the day the cells became macrophage foam cells) for assessment of the presence of CTX-I and ICTP and for an MMP activity assay.
Gelatine Zymography
MMP-2 and -9 expression and activity were determined by gelatinase zymography, which is able to detect both pro-enzymes and active enzymes because they migrate differently according to their molecular weight during sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis electrophoresis. Ten microliters of supernatants from cells cultured on plastic at day 1, 3, and 10 was loaded together with a gelatinase zymography ladder for MMP-2 and MMP-9 (Chemicon) onto a gelatin gel (0.5 mg/mL gelatin) as a substrate in 7.6% SDS–polyacrylamide gels and proteins were separated. After electrophoresis the gels were washed with 2.5% Triton X-100 in water and then incubated overnight at 37°C in 0.1% Triton X-100, 5 mM CaCl2, 1 mM ZnCl2, 3 mM NaN3, and 50 mM Tris buffer, pH 7.4, in a closed container. Gels were stained for 30 min with 0.25 Coomasie R-250 (Sigma-Aldrich) in 10% acetic acid and 45% methanol and then stain removed over 30 min with 20% acetic acid, 20% methanol, 17% ethanol, and 0.6% diethyl ether. Gels were dried and scanned for documentation.
MMP Activity Assay
The commercial fluorogenic peptide Bachem substrate M-1895 (Mca-Pro-Leu-Gly-Leu-Dap(Dnp)-Ala-Arg-NH2; trifluoroacetate salt; Bachem) was diluted to 2.66 μM in an incubation buffer consisting of 0.01 M CaCl2, 0.2 M NaCl, 0.05 M Tris-HCl, 50 μM ZnSO4, and 0.05% Triton-X 100 (pH 7.5). It was incubated 3:1 with supernatant, from cells cultured on plastic, in a black 96-well plate at 37°C for 15 min. Cleavage by active MMP at the Gly-Leu bond separates the highly fluorescent Mca group from the efficient 2,4-dinitrophenyl quencher, resulting in an increase in fluorescence intensity. This cleavage was detected by reading the fluorescence at 328 nm excitation and 392 nm emission. As positive control, activated MMP-9 (Abcam) was used diluted with incubation buffer to 0.01 mg/mL. The negative control used was a 50 μM GM6001 solution (Table 1).
Matrix Metalloproteinases Activity Protocol Table
1. Dispense supernatants and controls to a black 96-well plate
2. Add 75 μL fluoregenic peptide
3. Incubate plate
4. Read the fluorescence
MMP, matrix metalloproteinase.
Western Blotting
The cell lysates were diluted in sample buffer and reducing agent and heated for 10 min at 72°C. A volume of 5 μL of cell lysates and 5 μL marker (Rainbow marker; Amersham) were loaded on 4%–20% pre-cast gels (Bio-Rad) under reducing conditions (migration buffer: Tris 25 mM, Glycine 200 mM, and SDS 3.5 mM, pH 8.3). After transferring the proteins to a nitrocellulose membrane at 30 V in transfer buffer, the membranes were blocked in PBST (1.5 mM KH2PO4, 8 mM Na2HPO4·2H2O, 2.7 mM KCl, and 150 mM NaCl with 0.05% Tween-20) with 5% skimmed milk for 1 hour at room temperature. After blocking, the membranes were incubated with primary antibodies (monoclonal αCathepsin K [Chemicon], polyclonal chicken αLDL receptor [Abcam], monoclonal mouse αMMP-9 [Abcam], polyclonal goat MSR-1 [AbD Serotec], monoclonal mouse CD68 [AbD Serotec], and αTIMP-1 [kindly provided by Department of Veterinary Disease Biology, Faculty of Life Sciences, University of Copenhagen]), respectively, overnight at 4°C. Samples were calibrated using glyceraldehyde 3-phosphate dehydrogenase (GADPH). After washing, the membranes were incubated with peroxidase-labeled goat, rabbit, mouse, or chicken antibody for 1 h at room temperature and the results were observed using the ECL Plus Western blotting detection reagent (GE Healthcare, Amarsham).
Biochemical Analysis
Supernatants from cells cultured on matrigel were analyzed for degradation fragments of the CTX-I using the CrossLaps for culture kit (Nordic Bioscience Diagnostics, IDS Nordic A/S), according to the manufacturer's instructions. The samples were used undiluted to reach the outlined concentration intervals. Briefly, the kit is based on monoclonal antibodies recognizing the isomerized amino acid sequence EKAHD-β-GGR specific for the C-telopeptide of type I collagen and D-β-G representing the isomerized bond between aspartate and glycine. The sandwich ELISA that detects crosslinked peptides (CTX-I) has been reported to have intra-assay and inter-assay coefficient of variation of 4.9% and 7.9%, respectively, and is useful for the quantitative assessment of bone resorption. 53
The release of the ICTP from the cultured cells on matrigel was measured using the UniQ ICTP EIA kit (Orion Diagnostica) according the manufacturer's instructions. These samples were diluted four times to reach the outlined concentration intervals.
Statistics
Data from the cultured cells on matrix were analyzed by one-way analysis of variance with Bonferroni correlation of nine groups, in which the mean of the outcomes in each type of the measurements (MMP activity, ICTP, and CTX-I, respectively) was compared. When comparing the mean outcome from any two groups, a Student's two-sample t-test was used. Differences were considered statistically significant if P<0.05.
Results
Characterization of Phenotypes During Cell Differentiation
Human CD14-positive monocytes cultured on plastic in the presence of M-CSF assumed macrophage appearance as early as 1 day of treatment, counting from the day the macrophages were cultured into the 24-well plate, so at this time the monocytes have been incubated in presence on M-CSF for 7 days in total (Fig. 1). When cultured with M-CSF and LDL, the macrophages transform into foam cell-like appearance as early as 3 days of LDL treatment. During transformation, macrophages showed marked alterations in their morphology, increasing in size and tending to fuse to formation of multinucleated cells. The macrophage foam cells and not the macrophages were oil red O positive, indicating the presence of lipids.

Macrophages differentiate into macrophage foam cells in presence of LDL. CD14+ monocytes were isolated from peripheral blood of healthy humans and differentiated into macrophages in presence of M-CSF for 6 days. These macrophages were cultured on plastic in presence of either M-CSF alone (the four top pictures) or M-CSF and 50 μg/mL LDL (the four lower pictures) for 1, 3, 7, and 10 days. After the culture period the cells were fixed in 4% formaldehyde and stained for lipid droplets by oil red o staining. When incubating the macrophages in presence of M-CSF and LDL, they differentiate into lipid-laden macrophage foam cells with marked alteration in morphology. They greatly increased in size and tended to fuse into multinucleated cells. All the pictures are magnified 20×. M-CSF, macrophage-colony stimulating factor; LDL, low-density lipoprotein.
Characterization of the level of protein expression during the macrophage foam cell differentiation using western blotting of lysates of cells exposed to only M-CSF or M-CSF in combination LDL showed that all three scavenger receptors, CD68, RS-AI, and LDL-R, were present from day 1 (Fig. 2). Over time, however, expression of CD68 and SR-A1 decreased and addition of LDL to the cultures did not seem to affect the level of receptors. The proteases, MMP-9, cathepsin K, and TIMP-1, were also present in the cell lysates and were expressed differentially over time. Cathepsin K expression was highly upregulated in the macrophages as a function of time, and both profrom and active form were observed. This upregulation was also observed for macrophage foam cells, but not to the same extent. TIMP-1 was downregulated as a function of time, whereas MMP-9 was not affected by time or exposure to LDL. GADPH was present at all time points.
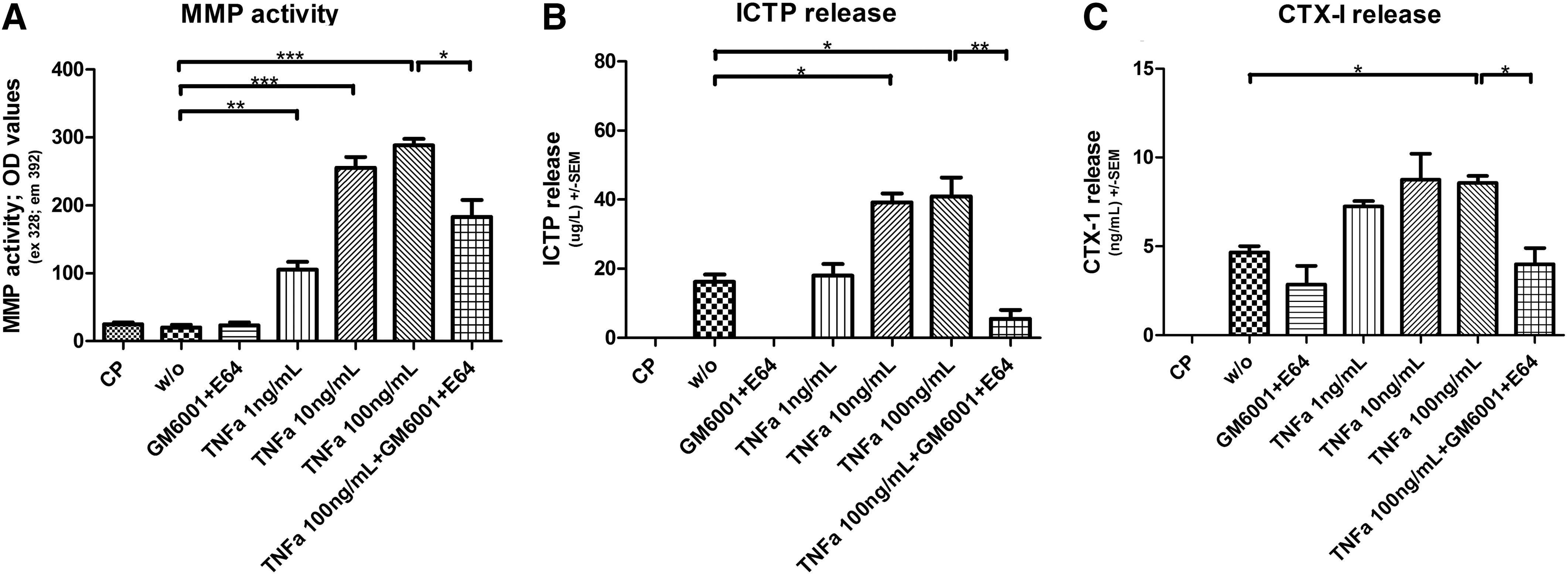
TNF-alpha stimulates MMP expression and matrix degradation. Human macrophages were cultured on a matrigel enriched with collagen type I and in presence of 50 μg/mL LDL. Different concentrations and combinations of TNF-alpha and protease inhibitors were added to the macrophages as indicated on the graphs; w/o indicate no treatment and CP are cells cultured on plastic. MMP content and matrix degradation were obtained by measuring the supernatants collected at day 7. The amount released by matrigel alone has been subtracted and so the results are background corrected.
Gelatin zymography (Fig. 3) showed that of all the gelatinases, only the MMP-9 pro-enzyme was present in the supernatant from macrophages and macrophage foam cells cultured on plastic (left panel). Levels of the MMP-9 pro-enzyme increased over time and the presence of LDL in the culture did not have a measurable effect on the protease level. Total MMP activity, calculated as the accumulated value of MMP activity at days 1, 3, 7, and 10 in supernatants by a fluorescence assay (panel B), showed, on the other hand, that adding LDL to the culture led to a significant increase in MMP activity.
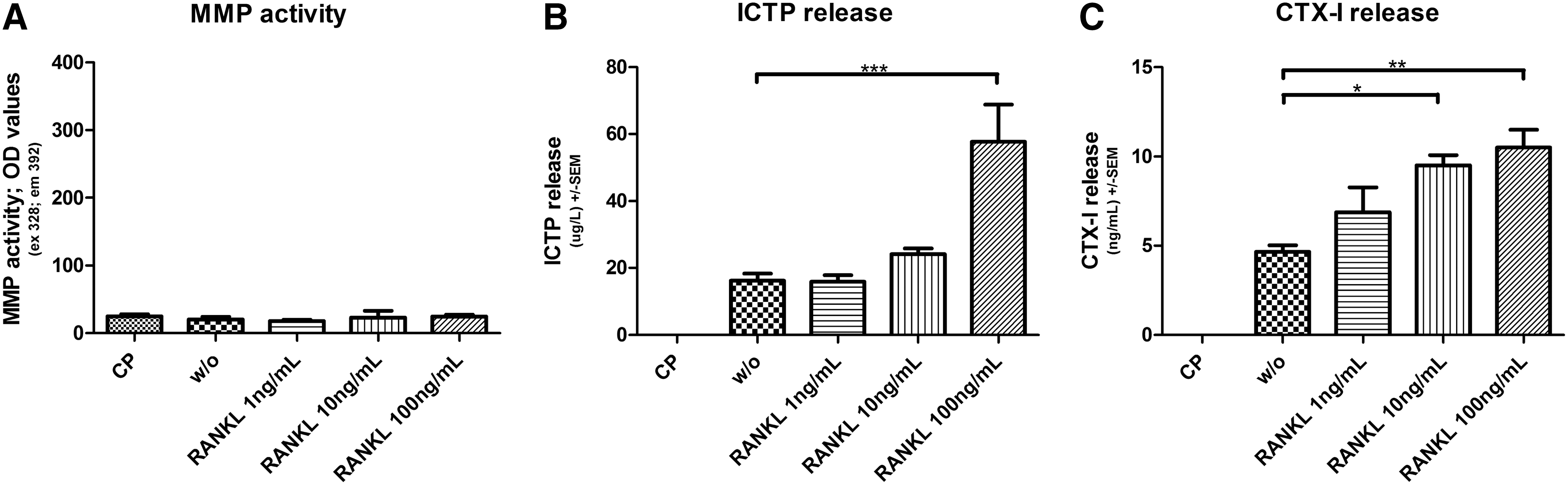
RANK-L induces matrix degradation. Human macrophages were cultured on a matrigel enriched with collagen type I and in presence of 50 μg/mL LDL. Treatments of 1, 10, and 100 ng/mL RANK-L were added to the macrophages as indicated on the graphs; w/o indicate no treatment and CP are cells cultured on plastic. MMP content and matrix degradation were obtained by measuring the supernatants collected at day 7. The amount released by matrigel alone has been subtracted and so the results are background corrected.
RANK-L and TNF-Alpha Induce Macrophage Foam Cell-Mediated Matrix Degradation
Addition of different concentrations of TNF-alpha, protease inhibitors, and RANK-L to human macrophage foam cells cultured on a matrigel enriched with collagen type I was followed by dose-dependent increases in the release of CTX-I and ICTP in supernatants on day 7 (Figs. 2 and 3). Day 7 was chosen for the measurements since the activity of the cells was highest on this day (data not shown).
A statistically significant increase in MMP activity was observed when incubating macrophage foam cells in presence of TNF-alpha, compared with no treatment (P<0.001). However, no difference was seen with the addition of RANK-L.
The release of the MMP-generated type I collagen fragment, ICTP, was significantly increased after the addition of TNF-alpha compared with untreated macrophage foam cells and a dose-dependent response was observed. Ten ng/mL TNF-alpha raised the release of ICTP by 59% (P<0.001). RANK-L also showed 72% (P<0.001) increase in ICTP release when 100 ng/mL was added to the cell culture (Fig. 3B). The cathepsin K-generated fragment, CTX-I, was released in significantly higher amounts when macrophage foam cells were incubated with at least 10 ng/mL of RANK-L (P<0.01) and 10 ng/mL TNF-alpha (P<0.01) compared with cells receiving no treatment (Figs. 3C and 2C). The 100 ng/mL RANK-L increased the release by 56% (P<0.01) and 100 ng/mL TNF-alpha increased the release by 43% (P<0.01). A significant decrease in TNF-alpha-induced MMP activity (P<0.05), ICTP release (P<0.001), and CTX-I (P<0.01) was observed when both protease inhibitors GM6001 and E64 were present (Fig. 2).
Discussion
ECM remodeling is receiving increased attention in many pathologies. 54 Until recently, protein expression and protease expression were investigated independently and not the interactions. With the availability of new tools, protease activities on specific proteins of the ECM can be carefully investigated.
We characterized a human macrophage foam cell system based on isolation of CD14+ monocytes in which type I collagen degradation can be investigated. The method of isolation of CD14+ monocytes and differentiation into macrophages has been successful at deriving osteoclasts, 35 for which reason this method was employed. When CD14+ monocytes were exposed to M-CSF a traditional macrophage phenotype was obtained and when exposed the macrophages to LDL, cholesterol was accumulated intercellular and a traditional macrophage foam cell phenotype 55 was obtained characteristic of macrophage foam cells in plaques. In this system native LDL was used and not modified LDL, since studies have showed that cultured macrophages can modify LDL, resulting in a marked increase in cell cholesterol. 56 To further investigate the in vitro phenotypes, human macrophage and macrophage foam cells were demonstrated to express traditional scavenger receptors and (CD68 and SR-A1) as well as LDL-R, which are able to take up the LDL. Interestingly both CD68 and SR-A1 were downregulated as a function of time, whereas in the literature monocyte-to-foam-cell differentiation has been described to upregulate the gene expression of several scavenger receptors. 57 What may be the cause for this difference has not been further investigated in this study.
Of particular relevance for the current investigation is the balance between proteases and their inhibitors, as a net catabolic potential would enable proteases to degrade the ECM, MMPs to generate ICTP, and cathepsin K to generate CTX-I. We observed a decrease in the expression of the MMP-inhibitor, TIMP-1, over time on western blot, whereas the MMP-9 expression was stable. However gelatine zymography of MMP-9 in supernatant showed an increase of proMMP-9 over time. This is in complete alignment with the inhibitor role of TIMP-1; reduced levels of TIMPs result in increased levels of MMPs. 7 Overexpression of TIMP-1 in apolipoprotein E knockout mice has also been shown to reduce lesion development, 23 which is in alignment with the statement that MMPs induce ECM remodeling. 58 Neither the western blot nor the gelatine zymography showed changes in expression of MMP-9 when exposed to LDL, whereas the total MMP activity was observed to increase with exposure of LDL. The reason for this might be that the total MMP activity is the final balance between the MMPs, their activators, and their inhibitors, for example, TIMP-1 reflecting a proteolytic potential of the culture. The non-increase of MMP-9 also indicates that MMP-9 might not be representative of MMPs overall in macrophage foam cells. This is conflicting with what is known for osteoclasts, which among the MMPs mainly are secreting MMP-9. 35,37 The balance between MMPs and ECM remodeling has received recent attention in the literature, 59,60 and may be important in the evaluation of potential intervention strategies and pathways that modulate the cell phenotype. To further investigate whether this altered balance would result in matrix degradation, and could be further augmented by pro-inflammatory molecules, we used human macrophage foam cells cultured on an artificial matrix consisting of type I collagen and a gelatinous protein mixture resembling the complex extracellular environment found in many tissues. A similar approach has been used for other cell types in different pathologies. 34
The in vitro system demonstrated that in macrophage foam cell culture (1) both MMP- and cathepsin-generated neoepitopes of type I collagen were detected, enhancing the previous statement, that macrophage foam cells do secrete MMPs and Cathepsin K, and (2) when stimulated with TNF-alpha and RANK-L, a dose-dependent increase was seen in the levels of these neoepitopes. These are the first data using an in vitro system in combination with neoepitopes to investigate remodeling by human macrophage foam cells. Other investigators have examined cell–matrix interactions, histomorphology, and immunohistochemistry in similar in vitro models. 61
Interestingly, TNF-alpha stimulation resulted in a significant increase in MMP-mediated degradation of type I collagen, as indicated by levels of ICTP, compared with unstimulated macrophage foam cells. In contrast, no significant increase was seen in cathepsin K-mediated degradation, indicated by levels of CTX-I, compared with untreated cells. In alignment with these findings, TNF-α exposure resulted in highly significantly increased levels of total MMP activity in the human macrophages foam cells. The lack of CTX-I release shows that TNF-alpha does not stimulate cathepsin K secretion, which is in keeping with the fact that ICTP values increase and therefore are not degraded by cathepsin K. The MMP-mediated degradation was further inhibited by the protease inhibitors, as expected, which is a positive control for the protease dependence.
Stimulation with RANK-L resulted in a different pattern from that observed with TNF-alpha treatment. This is somewhat expected due to their induction of different pathways. 62,63 Unlike TNF-alpha, RANK-L stimulation did not result in increased MMP activity. However, the highest dose of RANK-L resulted in induction of MMP-mediated type I collagen degradation, which correlates well with a previous study showing that high doses of RANK-L lead to CTX-I by osteoclasts cultured on decalcified bone matrix. 64 In contrast, a dose-dependent and significantly increased release of cathepsin K-mediated type I collagen degradation was observed. This is in alignment with the strong evidence for RANK-L-driven cathepsin K expression and activity in osteoclasts, 35 and also correlates well with the fact that TNF-alpha can induce multinucleation of osteoclasts, yet fails to induce bone resorption measured by CTX-I, as process in which cathepsin K is essential. 65,66 Since RANK-L seems to have a stimulating effect on the macrophage-driven ECM remodeling, it is of high interest to investigate whether the anti-RANK-L intervention for osteoporosis could have additional benefits in atherosclerosis.
The different reactivity pattern between TNF-alpha and RANK-L leads to the question whether there are types of arterial lesions where RANK-L plays a more dominant role than TNF-alpha? This would be interesting to investigate in the future.
These results emphasize that the collagen turnover profile is of pivotal importance when investigating ECM remodeling. One collagen fragment may not be affected, whereas other fragments produced by the activity of different proteases may be affected. Hopefully, this increased understanding of mechanisms leading to matrix destruction, and the measurement of that, may lead to a deeper understanding of the macrophage phenotype that is associated with pathological matrix remodeling and plaque rupture.
In conclusion, the results from these experiments clearly demonstrate that human macrophage foam cells produce cathepsin K and MMPs, which degrade type I collagen, resulting in remodeling of the ECM. These findings support the studies that showed MMPs' involvement in arterial remodeling in animal models. 67 These are, to our knowledge, the first data describing a simple in vitro system in which human macrophage foam cell degradation of the ECM can be monitored and, furthermore, enhanced by cytokines. This system may be useful in testing modulators of matrix remodeling and thus aid in the identification of novel molecular mechanisms and treatments that will attenuate atherosclerotic plaque turnover and subsequently prevent rupture of unstable plaques.
Footnotes
Acknowledgment
The authors acknowledge the funding from the Danish Science Foundation (Den Danske Forsknings fond).
Author Disclosure Statement
M.A.K. owns stock in Nordic Bioscience. All other authors declare no competing financial interests.