
Abstract
Recent years have shown a great deal of interest and research into the understanding of the biological and physiological roles of mechanical forces on cellular behavior. Despite these reports, in vitro screening of new molecular entities for lung ailments is still performed in static cell culture models. Failure to incorporate the effects of mechanical forces during early stages of screening could significantly reduce the success rate of drug candidates in the highly expensive clinical phases of the drug discovery pipeline. The objective of this review is to expand our current understanding of lung mechanotransduction and extend its applicability to cellular physiology and new drug screening paradigms. This review covers early in vivo studies and the importance of mechanical forces in normal lung development, use of different types of bioreactors that simulate in vivo movements in a controlled in vitro cell culture environment, and recent research using dynamic cell culture models. The cells in lungs are subjected to constant stretching (mechanical forces) in regular cycles due to involuntary expansion and contraction during respiration. The effects of stretch on normal and abnormal (disease) lung cells under pathological conditions are discussed. The potential benefits of extending dynamic cell culture models (screening in the presence of forces) and the associated challenges are also discussed in this review. Based on this review, the authors advocate the development of dynamic high throughput screening models that could facilitate the rapid translation of in vitro biology to animal models and clinical efficacy. These concepts are translatable to cardiovascular, digestive, and musculoskeletal tissues and in vitro cell systems employed routinely in drug-screening applications.
Introduction
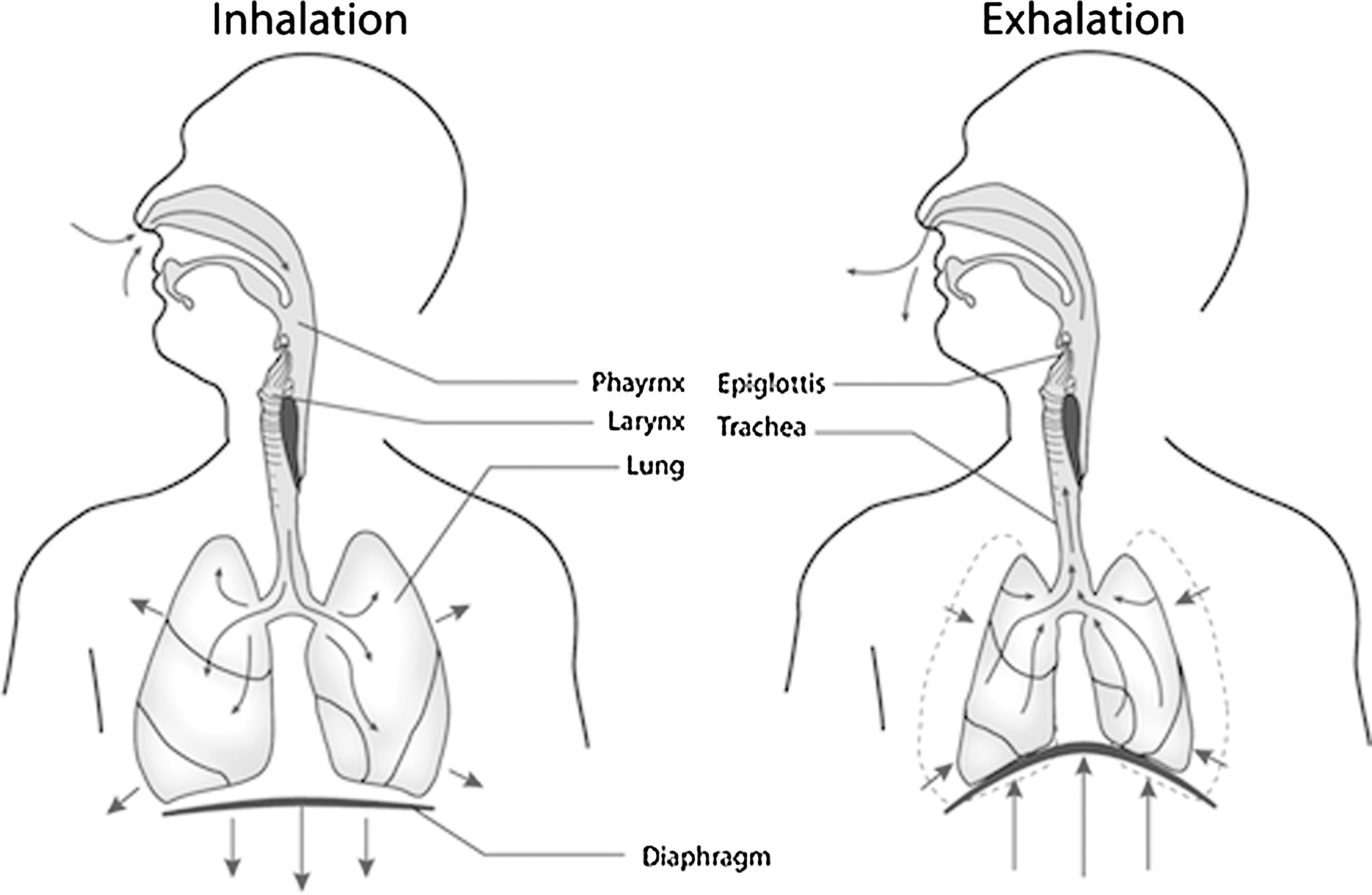
Schematic of lungs during respiration. Arrows show the movement of the lungs during inhalation and exhalation, resulting in a cyclic stretching of pulmonary cells.
The transduction of a mechanical stimulus into a cellular response in lung cells has been the subject of several excellent reviews over the past few years. 4 –6 Despite these reports, new molecular entities (NMEs) for pharmacological activity against lung diseases are routinely screened in static cell culture models that neglect the mechanically active in vivo environment. The purpose of this review is to engage drug discovery scientists on the effects of mechanotransduction in the lung and the use of bioreactor technologies, and to elaborate on the significance these technologies have in preclinical drug testing. Emphasis is given to the effects of cyclic stretch on cellular processes that are important during lung development and function such as DNA synthesis and proliferation, gene expression, protein expression, cellular differentiation, and surfactant secretion. Cellular pathways responsible for these processes are also discussed. Mechanical forces also play a critical role in the lung tissue under pathological conditions including lung injuries. Cells in lungs are often exposed to nonphysiological stretching as a result of disease or injury. The role of nonphysiological tensile forces representing those seen during disease and ventilator-induced injury are reviewed as well.
Early In Vivo Studies
Much of our understanding of mechanotransduction began with animal studies, and many in vivo animal studies of the lung have focused on studies of fetal development. Fetal breathing movements are important for the normal development of the lung, and our current understanding of the importance of fetal breathing movements is heavily dependent on previous work done in animal models. A variety of in vivo approaches have been taken toward understanding lung development, and each of these has led to a deeper understanding of the importance of mechanical forces in the developing lung.
The lungs are filled with fluid during development, and small changes in the fluid volume and pressure allow for a controlled lung expansion and contraction that are important for formation of a healthy lung. Early studies of changes in diaphragmatic electrical activity and tracheal fluid pressure in fetal sheep indicate that in utero breathing movements are correlated to changes in lung fluid flow. 7 The role of lung fluid volume in lung development has been demonstrated by manipulating the fluid volume in fetal animals through tracheotomy or tracheal obstruction. Fetal lambs undergoing tracheotomy showed decreased lung growth and distendibility, as well as decreases in DNA content. 8 Conversely, tracheal obstruction leading to decreased fluid drainage, and therefore increased and prolonged extension, has reportedly resulted in increased lung weight and increased alveolar structure, but decreased numbers of alveolar type II cells. 2 Tracheal ligation studies in fetal sheep by Moessinger et al. 9 have shown increases in lung DNA content as well as lung hyperplasticity. These studies indicate that, while controlled expansion of the lungs is important during development for proper lung growth, it is equally important to counteract any expansion with a similarly controlled contraction to keep lung cell proliferation under control.
The abolishment of fetal breathing movements through other mechanisms has also been employed to demonstrate similar findings. The creation of a hernia in the diaphragm as a means of decreasing lung expansion leads to decreased alveolar development and increased alveolar type II cell proliferation 10,11 as observed in lung tracheotomy studies. 8 Cessation of breathing movements has also been achieved by separating the lung from nervous system control. Phrenic nerve sectioning leads to the abolishment of fetal breathing movements through decreased diaphragm activity 12,13 and a decrease in airway fluid volume, lung weight, and total DNA content without altering the production of tracheal fluid. 13 Sectioning of the cervical spinal cord above the phrenic nerve in fetal rabbits also leads to cessation of fetal breathing movements without altering muscle tone of the diaphragm and has been shown to result in similarly hampered lung development. 3 Taken together, these studies indicate the importance of not only a functional diaphragm, but also of intact neuronal control of breathing and their importance in lung development.
While in vivo studies have significantly contributed to the fundamental understanding of the lung mechanotransduction at the organ-level, they do have limitations. Aside from ethical considerations that come into play in in vivo animal models, whole organism studies present greater difficulties in interpreting the results. Deconvolution of the mechanisms behind the treatment and the observed effect can be challenging on larger organ-level studies. Understanding mechanotransduction at the cellular level necessitates the precision and more tightly controlled conditions of in vitro models, and these factors contribute to the decreased popularity of animal models in mechanotransduction studies. Although no in vitro models perfectly mimic in vivo mechanical forces, bioreactors are currently being used to approximate breathing or ventilator-induced injuries and to study their effects in vitro. These dynamic in vitro models are also important from the drug discovery perspective because they have the potential to improve to the predictability of new therapeutic agents in preclinical translation.
Bioreactors: Bringing Forces into the Lab
Bioreactors are often used to simulate various types of in vivo forces such as tensile, compressive, and shear forces and perform in vitro studies to understand the effects of applied forces on cellular behavior. 4 To mimic the motion of the lung during respiration, lung cells are often subjected to cyclic tensile forces. With this approach cells are typically cultured on a flexible elastomeric membrane, such as polydimethyl siloxane (PDMS), that has been coated with a basement membrane component. A cyclic tensile force is then applied in one or more directions, allowing for physical manipulation of the cellular substratum in order to simulate the desired mechanical effect, which cells experience through interaction with the substratum as well as with neighboring cells (Fig. 2).
Selection of the appropriate mechanical force profile is important when attempting to mimic the dynamic nature of the in vivo conditions in an in vitro model. Although the precise change in surface area experienced by cells in the lung in vivo remains unclear, injury to cells exposed to a mechanical stretch in vitro is dependent on the force profile employed. Cellular injury in vitro has been shown to occur when changes in cell surface area exceed 37%, 15,16 a magnitude corresponding to inhalation greater than total lung capacity. The frequency of changes to cellular surface area in cyclic stretch profiles leads to cell damage primarily within the first few minutes of exposure to stretch, 16 and cell damage increases with increased amplitude and magnitude. 16 Changes in cellular surface area up to 30%, however, do not injure cultured cells 16,17 and are representative of the magnitude of stretch that cells experience in vivo.
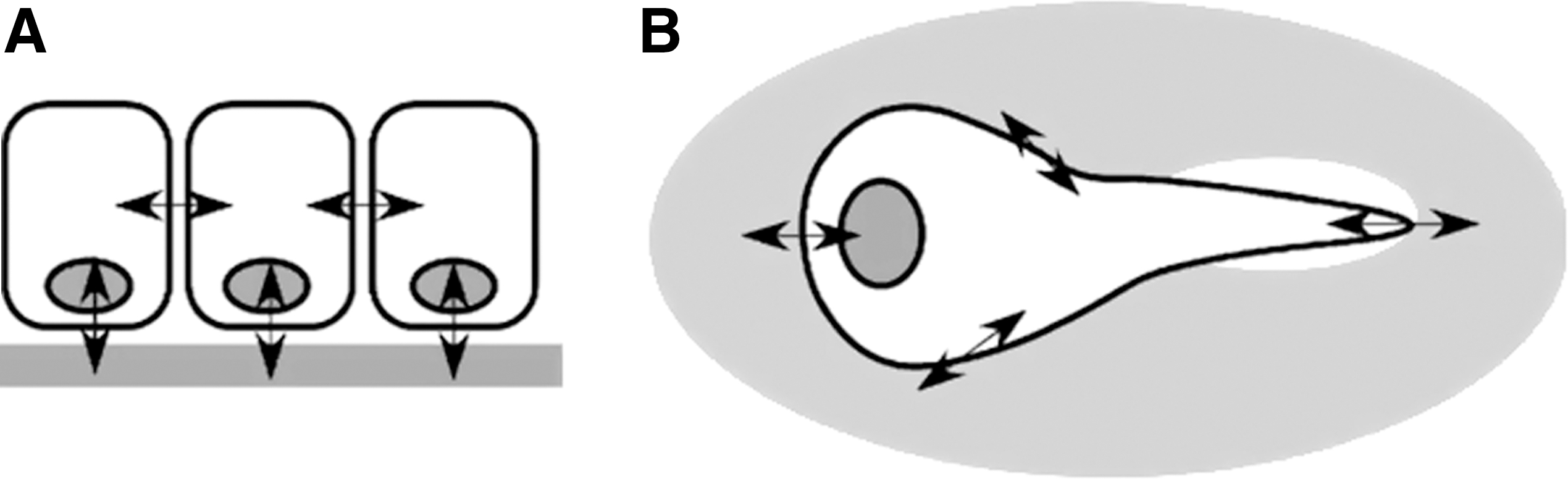
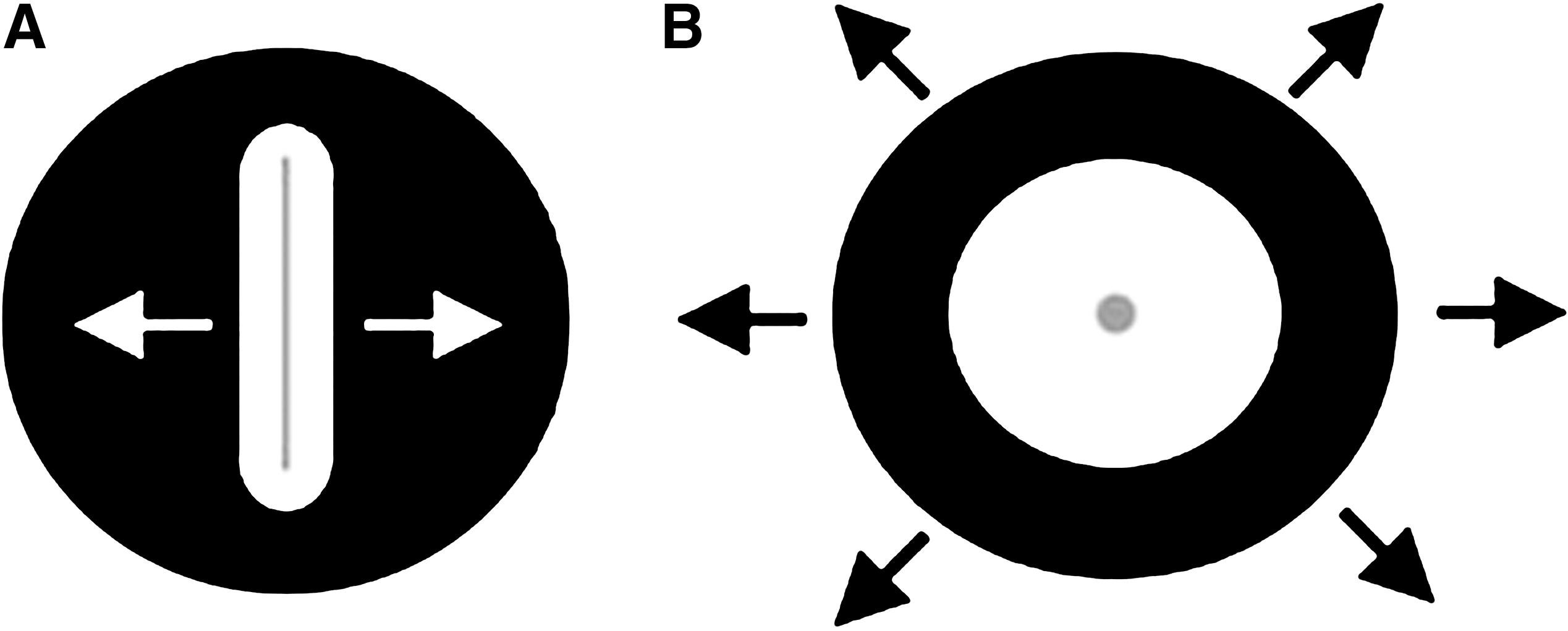
Applying a uniaxial stretch is one widely used approximation to study the effect of mechanical forces on lung cells (Fig. 3A). In this approach, cells are cultured on elastomeric strips or chambers and stretched from one or both ends, delivering a tensile force to the cells along a single axis. Kato et al. 19 used this method to study the effects of a cyclic uniaxial stretch on cyclooxygenase (COX) expression in lung fibroblasts. TIG-1 cells were cultured on fibronectin-coated silicon chambers and exposed to a cyclic uniaxial stretch, resulting in the up-regulation of both transcription and translation of COX-2. Another study by Hu et al. 20 utilized a similar approach using a “four-point” loading apparatus in which A549 cells were cultured on collagen I–coated bending dishes supported at two points from below. The bending dishes were stretched over these supports at two points from above, creating a homogeneous uniaxial stretch to study the up-regulation of intercellular adhesion molecule (ICAM)-1 transcription and translation in uniaxially stretched A549 cells. Uniaxial stretching experiments have also been conducted on strips of intact lung tissue. Mijailovich et al. 21 stretched rabbit lung parenchymal tissue strips sutured to a servo-controlled lever in order to study the mechanical response of the tissue to a uniaxial stretch. Although the uniaxial stretching of cells and tissues has provided much insight into lung biomechanics and biochemical behavior, it does not necessarily provide a representative model of the forces experienced by lung tissue in vivo.
Examples of
The use of biaxial distension adds force in an additional direction in an attempt to better model in vivo mechanical forces in vitro (Fig. 3B). One early example of such an approach utilized vacuum pressure to apply a tensile or compressive force directly to plastic petri dishes to study protein expression in tendon cells. 22 Another approach developed by Dassow et al. 23 used precision-cut rat lung slices that were mounted on polydimethylsiloxane membranes and subjected to a cyclic biaxial stretch within a bioreactor, showing increases in cellular calcium levels in stretched tissue. More typically, cells are cultured on flexible membranes that are built into multiwell culture plates. The plates are then used in conjunction with a bioreactor designed to distend the flexible bottoms of the plates, delivering both circumferential and radial strains to the cultured cells. One method of applying a biaxial force to cells involves a convex deflection of the culture surface from below, using air 24 or liquid. 25,26 Another involves pushing the culture surface down over a post 27 or driving a piston upward to stretch the culture membranes. 28 One of the most common methods currently employed involves the use of vacuum pressure to pull the culture membranes down over a fixed post. The Flexcell strain unit (Flexcell International, Hillsborough, NC), one commercially available instrument commonly used in mechanotransduction studies, uses this approach. With this device, cells are cultured in plastic six-well plates that contain flexible membranes as the bottom culture surface. Vacuum pressure is utilized to pull the flexible membranes down over a Delrin post (Fig. 4). Flexcell units have been used rather extensively to study the effects of mechanical strain on lung cell differentiation, 29 –31 proliferation, 32 –34 and gene expression 35 –37 to name a few examples. Although widely used in basic science research, this device has limited use in drug discovery due to its low throughput. Screening and lead optimization of NMEs require dose–response screening to determine overall activity (potency and efficacy) in different functional assays. This can efficiently be done only in a high throughput formats (i.e., in 96-well or higher density plates).
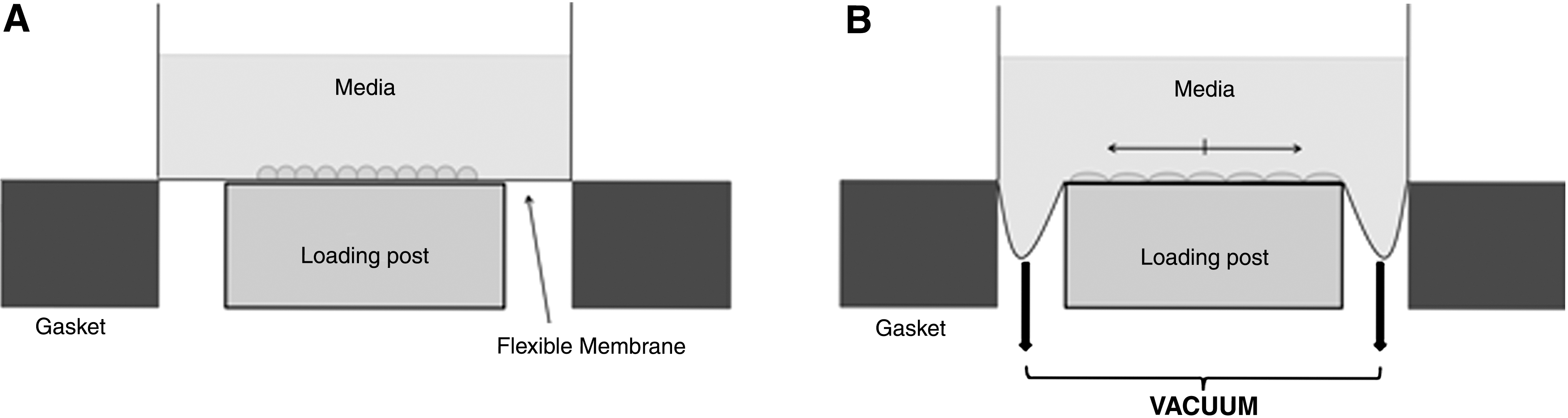
Schematic of the Flexcell Bioflex plate, showing the plate in a rest position
Recent attempts at creating microplate formats have resulted in miniaturized culture conditions that can be exposed to similar mechanical force profiles as those previously found in larger plate formats. Kamotani et al. 38 modified an automatically refreshing Braille machine in order to deliver cyclic tensile force to cells in microplate format. Cells were cultured on custom-fabricated flexible PDMS “chips,” and a tensile force was delivered to each well from below by raising the pins of the Braille machine. Moraes et al. 39 utilized a similar PDMS microarray in which a tensile force is delivered via a loading post driven by the application of positive pressure from below. Systems attempting to capture even greater biological complexity have recently been explored. Huh et al. 40 created microchannel systems that culture epithelial and endothelial cells on opposing sides of a porous PDMS membrane, re-creating the alveolar-capillary barrier. Mechanical strain is delivered by applying vacuum in chambers lateral to the culture membrane to simulate alveolar expansion during breathing. Chen et al. 41 and Krishnan et al. 42 cultured cells in gel substrates of varying stiffness to mimic different stiffness of in vivo soft tissue microenvironments and used punch indentation with a microneedle to deliver a uniaxial or biaxial external strain to cells cultured in these gels. Unlike Chen et al. 41 and Krishnan et al., 42 whose approach is currently in a larger culture format, Mih et al. 43 have developed 96- and 384-well formats for assaying cellular responses to varying matrix stiffnesses. These recent approaches are exciting improvements in replicating increasingly complex in vivo conditions in cultured cells. These micro-format approaches are also more amenable to high throughput screening of molecular libraries, making these culture systems potentially quite attractive for drug discovery and development. However, such plate formats are yet to be commercialized.
Cellular Responses to Stretch In Vitro
The transduction of a mechanical stimulus into a cellular biochemical response occurs in a number of ways, and mechanotransduction has been the subject of a number of reviews. 44 –46 An externally applied stretch can result in the opening of stretch-activated ion channels or in cytoskeletal changes resulting from interactions between two cells or between a cell and its extracellular matrix (ECM). When integrins at the boundary between a cell and the ECM are activated through interactions with ECM components, they cluster and promote association and activation of integrin-associated proteins, forming focal adhesion complexes. This in turn causes a cascade of cellular events that ultimately bring changes in proliferation and differentiation (Fig. 5).
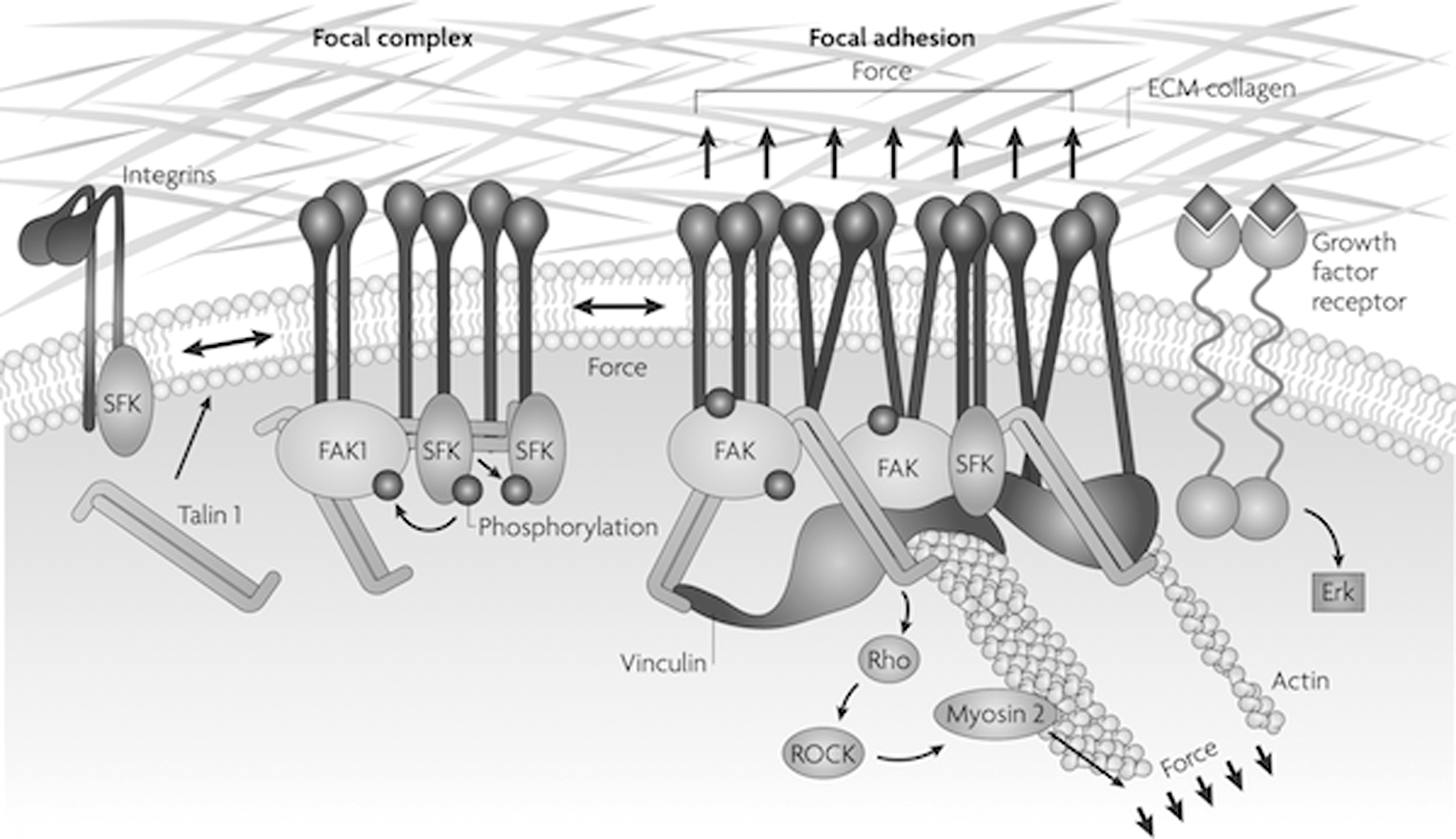
Mechanotransduction through interaction of integrins with the extracellular matrix (ECM). Activated integrins cluster promotes the association and activation of integrin-associated proteins and forms mature focal adhesion complexes. This leads to cellular pathways that modulate the cytoskeletal network through modification of actins and microtubules or activate pathways that lead to changes in gene expression and proliferation. Used with permission from Butcher et al. 47
Proliferation
Much in vitro research has been conducted using bioreactors to study the relationship between lung dynamics and cellular behavior, and many of these studies have focused on the fetal development of lungs. Fetal breathing movements expose the lungs to mechanical forces even before they serve the purpose in gas exchange, and play an important role in fetal lung development. 48 Changes in proliferation that result from mechanical engagement of the cell (Fig. 6) have been observed in a number of pulmonary cell types (Table 1).
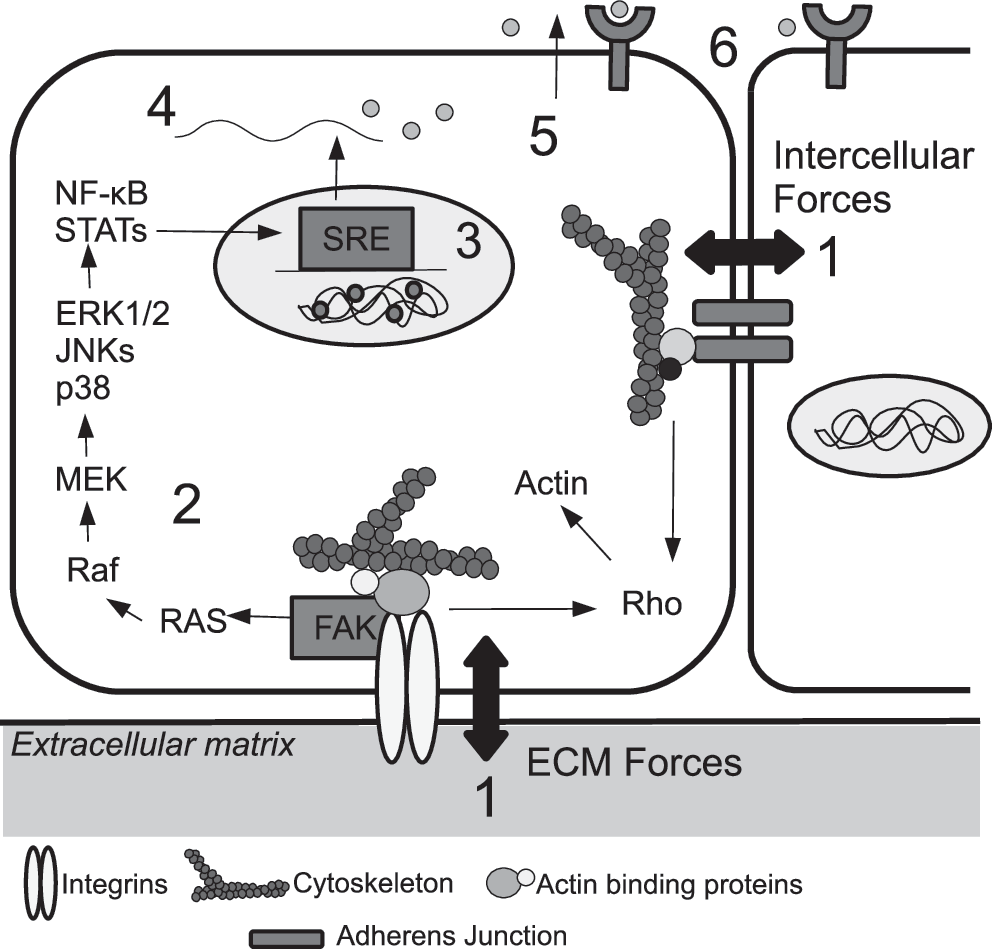
Mechanotransduction can affect cellular proliferation through a cascade of events. A signal is initiated at the exterior of the cell through interactions of integrins with the ECM or at cell–cell junctions (1), which in turn activates various cell-signaling pathways (2). An example depicted here involves Focal Adhesion Kinase (FAK), which is constitutively associated with integrins, and is activated when integrins interact with components of the ECM. FAK-activated changes in transcription factor activity lead to changes in gene transcription through activation of stretch responsive elements (SREs) (3). Protein translation (4), and secretion of protein end products such as growth factors (5) are also affected, resulting in further effects on the same or neighboring cells (6). Adapted with permission from Liu et al. 4
Comparison of Cellular Responses to Stretch
Fibroblasts play an important role in extracellular matrix deposition and regulation of alveolar structure, and the role of mechanical stimulation in their proliferation is a subject of debate. Human lung IMR-90 fibroblasts showed an increase in proliferation when exposed to a cyclic biaxial stretch in two-dimensional cultures, and cellular replication is further increased in IMR-90 cells cultured in media extracted from the wells of mechanically stimulated cells. 54 In contrast, fibroblasts isolated from rats during the canalicular stage of development demonstrated a decrease in proliferation in response to a tensile force. 53 Liu et al. 55,57 studied both two-dimensional cultures and “organotypic” cultures, three-dimensional co-cultures composed of mixed fibroblast and epithelial cells isolated from rats during the canalicular stage of lung development (at 19 days of gestation). While measurement of DNA incorporation of [3H]thymidine showed no significant change in proliferation in two-dimensional cultures, increases in proliferation in three-dimensional organotypic cultures in response to a cyclic uniaxial stretch were observed. The decreases in proliferation in two-dimensional fibroblast cultures 53 and the increases in three-dimensional organotypic cultures 55,57 could be attributed to differences in the applied stretch (uniaxial or biaxial) or differences in cell–cell and cell–ECM contacts between two- and three-dimensional cultures. 56 Xu et al. 58 reported that strain-enhanced proliferation is gestation dependent, with proliferation being the highest during the early canalicular stage at 19 days of gestation. This gestational response to mechanical force appears to be regulated by the mesenchyme. These mostly undifferentiated and loose connective tissue cells of mesodermal origin easily migrate and may have different responses to drugs when compared to cells of epithelial origin. Note that epithelial-mesenchymal transitions and vice versa (mesenchymal-epithelial transitions) are critical in cancer pathology, and hence the effect of forces could be important in screening applications in oncology. 40
Changes in cell cycle controls and apoptosis have been investigated as possible mechanisms of the observed changes in proliferation in response to mechanical forces. 52,53,59 Cells progress through well-defined stages during the cell cycle prior to mitosis. The orderly duplication of nonchromosomal cellular contents in the G1 phase, followed by the duplication of chromosomes in the S phase, is necessary for mitosis and the proliferation of cells. Inhibition of a G1/S phase transition has been observed in vascular smooth muscle cells in response to cyclic stretch. 52 In a similar study, Sanchez-Esteban et al. 53 observed significant increases in quiescent (G0) and G1 phase cells and significant decreases in S phase cells in biaxially stretched fibroblasts, indicating an inhibition of cell cycle progression. The enhanced apoptosis observed is due to increased caspase-3 activation under stretched conditions. 53,59 This evidence suggests that stretch may serve as a mechanism for controlling proliferation and guiding normal lung development by inducing apoptosis and changes in cell cycle signaling events.
Although the cellular pathways potentially involved in proliferation changes in response to stretch are yet to be fully understood, changes in growth factor levels and activity, and ECM degradation and remodeling may also collectively modulate the proliferation of cells under stretched conditions. For example, hepatocyte growth factor stimulates alveolar type II cell DNA synthesis and proliferation. 60 Mechanical stretch induces the expression (and release) of hepatocyte growth factor from A549 cells at both the mRNA and protein level, which in turn results in increased activation of protein kinase C (PKC). 60 Stretch-induced activation of PKC has also been observed in organotypic fetal lung cell culture, 57 in addition to the increased expression of platelet-derived growth factor. 61 Upregulation of fibroblast growth factor receptor 62 and vascular endothelial growth factor (VEGF) 62,63 was observed in airway smooth muscle cells as a result of increased ERK pathway activation in response to cyclic stretching. 63 Apart from direct increases in growth factor transcription and release, the growth factor activity is also modulated through their interaction with extracellular glycosaminoglycans and proteoglycans (PGs). Rat fetal lung cells exposed to a cyclic and intermittent fetal-type mechanical strain demonstrated increased production of both glycosaminoglycans and PGs. 64 Binding of growth factors to PGs may enhance their activity and induce cellular proliferation. Changes in the amount of secretion of fibronectin, an important component in the degradation and remodeling of the extracellular matrix, have been observed in stretched organotypic mixed lung cell cultures. 65 Epithelial cells preconditioned with mechanical stimulation and co-cultured with fibroblasts synthesized more fibronectin, collagen III and IV, and matrix metalloproteinase-9 compared to nonmechanically stimulated controls. 66 These observations demonstrate that mechanical forces are critical in influencing cellular proliferation, extracellular matrix components, metabolism, and possibly the pharmacology of drugs tested.
Mechanical strain also affects the proliferation of lung cancer cells of different origins and characteristics. Many investigators have studied the stretch-dependent proliferation and the cellular pathways involved using various lung cancer cell lines, including lung adenocarcinoma epithelial cell lines, H441 33,34,51 and A549, 17,34,67 and H358-bronchioalveolar carcinoma. 67 Changes in MAPK/ERK signaling in response to mechanical forces have been investigated to elucidate the corresponding changes in proliferation of these cells. MAPK/ERK signaling is responsible for a variety of transcriptional changes in the cell including for genes and proteins ultimately involved in cell cycle control. Mechanical forces have been shown to stimulate ERK1/2 signaling in lung epithelial cancer cell lines, 20,33 as well as in primary lung epithelial cells via epidermal growth factor receptor (EGFR) tyrosine kinase. 30,49 Chaturvedi et al. 34 demonstrated that the mechanical strain-induced proliferation and ERK1/2 phosphorylation in human H441 pulmonary epithelial cells are mediated by Src and focal adhesion kinase. These findings have important implications in cancer drug discovery since most of the anti-cancer agents are mitotic inhibitors and target cell division (proliferation).
Differentiation
Mechanical forces are also important in fetal lung cell differentiation, predominantly in alveolar epithelial cell differentiation. Alveolar epithelial cells differentiate into Type I cells, which play a structural role in the lung, and Type II cells, which secrete surfactant necessary for respiration. Changes in the expression of surface markers for Type I and Type II cells were detected in response to stretch. 29,37,50,68 A static distension decreases the expression of Type II markers, surfactant proteins A and B (SP-A and SP-B), in H441 epithelial cells 37 and in rat Type II cells, 50 indicating that a static distension promotes Type I cell differentiation. Conversely, expression of the Type II phenotype is increased with mechanical contraction 68 and cyclic distension, with a cyclic distension increasing expression of SP-A and SP-B and the production of surfactant phospholipids in fetal rat lung epithelial cells. 29 Cyclic stretching of isolated rat Type II cells was later shown to increase the Type II markers SP-B and SP-C, with further increase in SP-C protein expression observed with the addition of fibroblasts. These increases were due to increased expression of SP-B and SP-C mRNA. 29 The type of mechanical stimulation (static versus cyclic) determines the expression of Type I and Type II cell phenotypes, suggesting that both distension and contraction are important for balancing the differentiation of Type I and Type II cells in the lung.
One possible mechanism for the stretch-induced changes in gene expression could be through the cyclic AMP (cAMP) pathway. cAMP is an important second messenger responsible for changes in multiple cellular pathways. A cAMP-PKA–dependent signaling pathway mediates changes in phenotypic expression in fetal rat lung Type II cells in response to cyclic strain. 31 Type II cell maturation under stretched conditions may also involve EGFR activation. It has been reported that, under stretched conditions, EGFR plays a role in the mediation of fetal Type II cell differentiation 30 and increases ectodomain shedding of mature EGF ligands, transforming growth factor (TGF)-α and heparin-binding epidermal growth factor (HB-EGF) in fetal rat lung cells. 69 Further probing for Type II markers, SP-B and SP-C, in the cells exposed to TGF-α and HB-EGF showed increased levels of SP-B and SP-C, indicating that TGF-α and HB-EGF promote Type II cell maturation. 69 These observations highlight the importance of the fetal breathing movements on the activation of cellular pathways and the critical roles they play in the normal epithelial cellular differentiation in the lung.
Injury
Despite mechanical stretching being important for normal fetal lung development, excessive stretching often results in lung injury. Mechanical ventilation, a treatment often used for patients with respiratory distress or for preterm neonates, is the primary cause of lung injuries due to excessive stretching. The high lung tidal volume associated with ventilator-induced lung injuries (VILI) often results in increased inflammatory responses and abnormal behavior of lung cells. Copland et al. 70 studied the progression of VILI at a cellular level in rat models and reported that 30 min of mechanical ventilation up-regulates the genes responsible for stress proteins and inflammatory mediators. However, an additional 60 min of mechanical ventilation resulted in lung injuries. The airway inflammatory response involves the increased recruitment of neutrophils to the airway by chemoattractant signaling of inflammatory mediator, interleukin (IL)-8, which is released in response to stretch. IL-8 is a strong chemoattractant of neutrophils and reportedly up-regulates in response to a 30% 71 or 40% 72 cyclic deformation in alveolar epithelial cells. The mechanism by which IL-8 is increased in response to stretch may vary depending on cell type. For example, ERK1/2 and p38 activation were shown to play a role in increased IL-8 in stretched bronchial epithelial cells; 73,74 however, alveolar epithelial cells under stretched conditions demonstrated a dependence on the activation of stress kinase, c-Jun NH2–terminal kinases. 75 In addition to increased IL-8 release, increases in macrophage inflammatory protein-2, a rat analog of IL-8, 76 and in ICAM-1 in human alveolar cells 20 are also observed in response to cyclic mechanical deformation. Up-regulation of these pro-inflammatory mediators contributes to the development of inflammation and edema seen in ventilated patients in response to excessive mechanical stretch.
Repair of a damaged epithelial barrier after injury is critical for restoration of healthy lung barrier function. Artificial “wounds” created in cell monolayers subjected to both cyclic compression and elongation showed decreased ability to “repair,” with small relaxation time between deformations being directly related to less wound closure. 77 Desai et al. 78 demonstrated that airway epithelial cell wound closure is ultimately decreased in response to stretch due to the modest cellular migration. Phosphatidylinositol 3-kinase (PI3-K) is typically activated in response to a wound and subsequently contributes to cellular motility. Cyclic stretch inhibits PI3-K activation and decreases cell migration after wound formation. Mitigating the decreased cellular migration and wound repair resulting from high levels of stretch is an important factor in recovery from injury for mechanically ventilated patients.
Mechanical stimulation has also been shown to play a role in bacterial growth in the lung. An important complication associated with ventilated patients is the increased risk of nosocomially acquired bacterial pneumonia. Epithelial cells exposed to a cyclic stretch comparable to a stretch experienced during ventilator therapy resulted in increased acidification of the growth environment and the growth of several bacterial strains. 79 The Na+/K+-ATPase activity associated with the acidification 79 was previously shown to increase in epithelial cells exposed to a cyclic strain. 80,81 Therefore, exposing lung cells to increased amounts of stretch, such as during mechanical ventilation, may be an unintended and unfortunate contributor to the establishment of bacterial pneumonia in mechanically ventilated patients. The application of excessive mechanical stretch to the lungs, as seen with increased air volume and pressure associated with high-volume mechanical ventilation, has many adverse effects on the lungs at a cellular level. An understanding of the mechanical stimuli involved in VILI, their cellular pathways, and the deleterious effects they ultimately cause in the lung is necessary for the development of ideal ventilator therapies for injured patients and preterm neonates.
Applicability in Drug Discovery and Screening
As our understanding of the pathways involved in and influenced by mechanotransduction increases, the incorporation of physiologically relevant mechanical stimulation in cellular research paradigms will almost certainly become not only necessary but also routine. The current lack of incorporation of mechanotransduction has important implications for both our understanding of cellular processes in our bodies and for the development of new treatments for when these cellular processes go awry. Lung cancer is a particularly pernicious disease, being the leading cause of cancer-related death in 2009. 82 Recently, Kumar and Weaver 14 elegantly demonstrated in their review that tumor cells absorb and exert mechanical forces during the “force journey” starting from their participation in normal tissue homeostasis and continuing through all stages of tissue dysplasia, tumor cell invasion, and metastasis creating a dynamic relationship between tumor cells and their microenvironments. Therefore, it is reasonable to assume that the pharmacological responses of tumor cells to chemotherapeutic agents will also be influenced by mechanical forces constantly present in organ systems. For example, Onn et al. 83 reported that non-small cell lung cancer (NSCLC) cells demonstrate greater sensitivity to the microtubule inhibitor paclitaxel when implanted subcutaneously in the animal model than when implanted orthotopically in the lung. Among the many differences between the two tumor microenvironments, changes in mechanical stimulation experienced orthotopically in NSCLC's “native environment” versus that experienced subcutaneously could have played a significant role in altering the sensitivity of the cells to paclitaxel.
Another example elucidating the possible effect of mechanical forces on the pharmacological response of the tumor cells involves EGFR and vascular endothelial growth factor receptors (VEGFRs). 84 –88 The inhibition of EGFR and VEGFR activity in tumor cells has shown promise in the treatment of lung cancer. 84 However, such inhibition has been shown to be effective only in vitro. 84 –86 Although, no information is currently available on how the EGFR and VEGFR pathways respond to mechanical forces in human cancer cells, it has been shown that mechanical forces up-regulate the expression of these two receptors in normal epithelial 87 and endothelial cells. 88 It is possible that EGFR and VEGFR receptors of the tumor cells are also activated by mechanical forces in vivo, resulting in decreased potency of the molecular modulators and lead drug candidates identified using static cell culture screens. These differences in therapeutic response emphasize the need for improved drug screening methodologies that incorporate the effects of in vivo forces for identifying new anti–lung cancer agents that are efficacious not only in vitro but also in vivo.
Mechanical forces in the lung are also important from the drug delivery perspective. The successful delivery of plasmids for gene therapy in the lung is dependent in part on cellular changes stimulated by mechanical forces. It has been reported that a mild cyclic stretch using a 10% change in surface area applied to alveolar A549 cells results in increased nuclear localization of plasmids. 89 The cyclic stretch-induced nuclear localization can be attributed to the activation of the nuclear transcription factors activating protein-1 and nuclear factor-κB, leading to increased binding of these factors to plasmids, 89 and the stretch-induced reorganization of the cytoskeleton. 90 Increased vascular permeability and epithelial barrier disruption, conditions often observed with the excessive stretch associated with ventilator-induced lung injury, can also have important effects on drug delivery. Studies show that high magnitudes and frequencies of the applied stretch disrupt the epithelial barrier, resulting in increased barrier permeability. 91 In human pulmonary arterial endothelial cells, a pathologically relevant stretch amounting to an 18% change in surface area results in increased permeability due to increased disruptive ability of thrombin, a regulator of endothelial barrier permeability. 92 Further study has indicated that increases in permeability regulated by agonists, such as thrombin, in response to stretch involve destabilization of the microtubule network and the activity of the microtubule-associated protein GEF-H1. 93 Stretch-increased epithelial barrier permeability in Type II alveolar cells has also been observed due to disruption of tight junction structure and decreased cell–cell adhesion associated with depletion of ATP and actin filament perturbation in mechanically stimulated cells. 94 Stretch-induced epithelial disruption increases the relative proportion of large-sized pores formed in the barrier, as observed through the transport of tracer molecules, increasing the potential for the transport of large molecules across the barrier. 95 Increases in barrier permeability can affect drug delivery, allowing for increased diffusion of larger molecular entities across barriers relative to the diffusion seen without stretch. Consideration for the effects of stretch on barrier integrity may therefore be important for screening of new drug candidates.
The response of cells to matrix stiffness also affects cellular responses to various drugs that inhibit cell growth. 43 Cells are able to detect and respond to variations in the stiffness of the substratum through interactions between integrins and the extracellular matrix, and this translates into changes in cellular proliferation and survival. 47,96 Vascular smooth muscle cells show increased platelet-derived growth factor receptor activity with increased matrix stiffness, resulting in increased proliferation. 97 Lung fibroblasts show increased activation with increased matrix stiffness, due to suppression of COX-2/prostaglandin E2 inhibition. 98 Proliferation in neuroblastoma cultures, however, is decreased with increases in matrix stiffness. 99 Interactions between cells are also modified by matrix stiffness. Endothelial cell monolayer integrity is affected by the rigidity of the underlying substratum, with increased stiffness resulting in increased formation of gaps between adjacent cells. 100 Matrix stiffness has recently been shown to play a role in the epithelial-to-mesenchymal transition in various cancer cell lines. 101,102 Although culture systems combining variable substratum stiffness with the application of an external stretch stimulus have been created, 41,42 they are yet to be used in drug screening applications.
Bioreactors as a means of mimicking the in vivo environment have only seen very limited use in drug discovery research. Kirsten et al. 103,104 have used perfusion bioreactors to study and characterize the effects of clinically relevant infusion rates and drug exposure of gemcitabine treatment regimens on cell death in NSCLC in vitro. To the best of our knowledge, however, the simulation of cyclic stretching on cells has not been investigated with regard to its effect on cellular responses to known and new drugs. A few studies have employed various drugs of known activity in order to clarify various cellular pathways involved in mechanotransduction. 33,51,55,56 Liu et al., 55 using leukotriene and prostaglandin inhibitors, demonstrated that the mechanical force-induced stimulation of fetal rat lung growth is independent of paracrine activity of leukotrienes and prostaglandins. They also reported that PKC inhibitor and phospholipase C (PLC) inhibitor significantly lowered the DNA synthesis, indicating that mechanical forces induce proliferation through activation of PLC and PKC. 56 Using the tyrosine kinase inhibitor genistein, Chess et al. 33 investigated the role of tyrosine kinases in strain-induced proliferation. In a different study that showed p42/44 MAP kinase inhibitors, PD98059 and UO126, block strain-induced proliferation, the same group concluded that the production of reactive oxygen species and p42/44 MAP kinase phosphorylation are also necessary in strain-induced proliferation of pulmonary epithelial cells. 51 Although the use of drugs in these studies has elucidated the various mechanotransduction pathways involved in cellular responses to forces, no drugs, to date, have ever been developed by incorporating the effect of mechanical forces during the early stages of drug screening.
Our group, in an effort to advocate the importance of dynamic cells culture models for drug screening, examined the effects of cyclic stretching on the pharmacological response of lung cancer cells and demonstrated that cyclic stretching significantly alters the effectiveness of the chemotherapeutic agents. 67 Our results showed that simulated mechanical forces that mimic normal respiration of human lung lowered the ability of four anti-cancer drugs, paclitaxel, doxorubicin, cisplatin, and a University of Kansas experimental drug, to inhibit the growth of A549 cells and increased the antiproliferative ability of Zactima™ in H358 lung cancer cells. 67 Our approach, if adopted in early stages of drug screening, may represent an important step in the development of physiologically relevant in vitro models, which holds great promises for decreasing the time, cost, and ultimately the failure rate of new drug candidates often seen in the transition from in vitro drug screening to preclinical animal studies and eventually to clinical trials.
To be able to incorporate the effects of mechanical forces into the early stage of drug screening, bioreactors that facilitate high throughput screening of chemical compounds or NMEs in the presence of simulated in vivo forces need to be developed. Unfortunately, such bioreactors are nonexistent and most of the existing approaches that deliver mechanical forces to cells in vitro are extremely low throughput, making their use cumbersome for drug screening applications. Screening and lead optimization of NMEs can efficiently be done only in 96-well or higher density plates using high throughput screening compatible screening assays. As discussed previously, miniaturizations of PDMS-based cell culture arrays are being developed that show promise in allowing for the concurrent study of a large number of cell samples under mechanically stretched conditions. 38,39,105 Such “microarrays” are important for the routine incorporation of mechanical forces into cell biology and drug development on a larger scale and will help bring mechanotransduction into the laboratory as a critical in vitro tool in drug discovery and development.
Footnotes
Acknowledgments
Disclosure Statement
No competing financial interests exist.