
Abstract
Human choriocarcinoma-derived BeWo cells express high levels of breast cancer resistance protein (BCRP/ABCG2) with no functional P-glycoprotein (P-gp) (ABCB1) activity, making them a potential model to study bidirectional ABCG2-mediated drug transport. However, the original BeWo clone (B24) available to researchers does not form confluent monolayers with tight junctions required by the model. Our aim was to adapt culture conditions to attempt to generate confluent BeWo monolayers for drug transport studies using the standard B24 clone. BeWo cells (B24; American Type Culture collection [ATCC]) were cultured in six-well plates or polycarbonate millicell inserts in a number of media formulations, growth supplements, and basement membrane substitutes. Cells were examined for confluence by microscopy, and transepithelial electrical resistance (TEER) was measured daily; monolayer permeability was assessed when TEER had stabilized. Optimal growth rates were achieved in culture conditions consisting of Medium 199 (M199) supplemented with epidermal growth factor (EGF; 20 ng/mL), vitamin supplements, and 10% fetal calf serum (FCS) with collagen coating. A TEER of 170 Ω in 0.6 cm2 inserts was achieved 2 weeks after seeding under optimal conditions. The cell-impermeable diffusion marker 5(6) carboxy-2,7dichlorodihydrofluorescein (C-DCDHF) had a permeability coefficient of 3.5×10−6 cm/s, indicative of minimal paracellular permeability. ABCG2 expression, as determined by immunoblotting, remained unaffected by confluency. In conclusion, we describe culture conditions for the B24 BeWo clone that facilitate the formation of monolayers with tighter junctions and reduced paracellular transport compared to previously published models. These growth conditions provide a good model of ABCG2-mediated drug transport in a human placental cell line.
Introduction
It has proved difficult to determine the characteristics of P-gp or BCRP drug substrates using molecular modeling. 8 Therefore, experimental characterization of drug efflux properties requires the development and application of in vitro models using cell lines with specific efflux properties. 9 The Caco-2 cell line, for example, expresses P-gp on its apical surface and forms a polarized epithelial monolayer in a culture with tight junctions to limit paracellular transport, making it ideal for bidirectional transport studies of P-gp efflux. 8,9 Currently, the only equivalent cell line for analysis of BCRP substrates is a Madin Darby canine kidney (MDCK) cell line transfected with human BCRP, 10 which is not widely available.
The placenta is the tissue with the greatest amount of BCRP expression, and placental trophoblast cells, which form the main epithelial barrier between the maternal and fetal circulations, are characterized by high levels of BCRP expression. The BeWo cell line is derived from human trophoblast cells and possesses two key trophoblast characteristics: high endogenous expression of BCRP 11,12 and propensity for cell–cell fusion and differentiation—a process known as syncytialization by which a multicellular epithelial membrane is formed across which all substances must pass in order to leave or enter the fetus. 13 In addition to expressing significant amounts of BCRP, BeWo cells express little ABCB1/P-gp, making them ideal for BCRP transport studies. 14,15 Other transporters are present on BeWo cells such as ABCB2 and some of the ABCC series of transporters, but these are not considered as important for drug efflux as BCRP and P-gp. Like many other human cell lines, BeWo cells will grow in most standard culture media in the presence of fetal calf serum (FCS) and are capable of secreting their own basement membrane to adhere to the surface of plastic culture dishes. However, under normal culture conditions these cells do not form confluent cell layers; instead, they aggregate and form discrete colonies. 14 At high seeding concentrations the cell colonies syncytialize and retract from other colonies, creating large gaps that preclude the development of intact monolayers. 15,16 This characteristic renders BeWo cells unusable for bidirectional transport studies. Although a subclone of BeWo cells (B30) has been shown to form tight junctions and has been used in transport studies, there has been very limited use of this subclone and the tight junctions described still allowed substantial paracellular transport. 17,18 Thus, our aim was to assess whether it is possible to adapt the culture conditions to encourage the B24 clone of BeWo cells available through national cell banks to display tight junctions and form a monolayer barrier in culture, with the ultimate goal of developing a useable in vitro model for BCRP-mediated transport studies.
Materials and Methods
Topotecan hydrochloride, 5(6)-carboxy-2,7-dichlorodihydrofluorescein (C-DCDHF), bicinchoninic acid (BCA), collagen type I from calf skin, anti-β-actin monoclonal antibody, the Dulbecco's modified Eagle's medium (DMEM), and Medium 199 (M199) were supplied by Sigma-Aldrich. Fumitremorgin C and forskolin were supplied by Enzo Life Sciences, Inc. PSC-833 was kindly donated by Novartis Pharmaceuticals. The Roswell Park Memorial Institute (RPMI) medium was from Hyclone. GIBCO minimum essential medium (MEM) nonessential amino acids (NEAA; Cat. No. 11140-050), GIBCO MEM vitamin supplement (100×; Cat. No. 11120-052), recombinant human epidermal growth factor (rhEGF) Ham's F-12 Nutrient Mixture with Kaighn's Modification (F12K medium), FCS (Cat. No. 10100-139 and 10100-147), GlutaMAX™, and peroxidase-labeled goat anti-mouse immunoglobulin G (IgG) antibody were supplied by Life Technologies Australia Pty Ltd. FCS was also supplied by Australian Commonwealth Serum Laboratories. Mouse monoclonal anti-human cytokeratin 7 antibody was from DAKO Australia. Precast electrophoresis gels were from PAGEgel Inc. Anti-P-gp MDR1 (G-1) mouse monoclonal, as well as anti-MRP1 mouse monoclonal (QCRL-1), and anti-MRP2 (M2 III-6) mouse monoclonal antibodies were supplied by SantaCruz Biotechnology, while anti-BCRP monoclonal antibody (BXP-21) was purchased from Chemicon (Life Technologies Australia Pty Ltd.). SuperSignal West-Pico Substrate was from Pierce Chemical. All other reagents were supplied by Sigma-Aldrich Pty. Ltd.
Immunohistochemistry
BeWo cells growing on 6- and 24-well plates or 0.6-cm2 inserts were visualized by staining with anti-cytokeratin 7 to determine levels of confluence. Cells were fixed in freshly prepared 4% paraformaldehyde in phosphate-buffered saline (PBS) for 10 min and subsequently blocked with 5% casein in PBS for 1 h. After incubation with primary antibody for 1 h, cells were washed in PBS and peroxidase-labeled secondary anti-mouse IgG antibody was added. After an hour, cells were subject to four stringent washes with PBS and color was developed using the 3,3′-diaminobenzide peroxidase substrate kit (Vector Labs); nickel was omitted for the staining of cells on the 6- and 24-well plates, but included when cells cultured on inserts were stained. Images were collected on a Nikon DS-5Mc camera attached to a Nikon Ti inverted phase microscope using NIS imaging software (Nikon Corp.).
Protein Determination
BeWo cells were scraped and homogenized in a protein lysis buffer consisting of 120 mM NaCl, 24 mM Tris base, 1% Triton-X-100, 0.1% sodium dodecyl sulfate (SDS), and 0.2% deoxycholic acid in water, pH 7.4. The lysis buffer was supplemented with Sigma Ultrafast general protein inhibitor tablets just before use. Protein concentrations in the lysates were determined using a micro-BCA method adapted for use with multiwell plates on a VersaMax plate reader (Molecular Devices). Curve fitting and data extrapolation were performed using SoftMax Pro40 software (Molecular Devices).
Western Blotting for Efflux Protein Detection
Thirty micrograms of proteins was added to each well of a 4%–12% Tris-glycine gel and subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) before transfer onto nitrocellulose membranes. They were washed in Tris-buffered saline/0.05% Tween 20, and then probed with monoclonal primary antibodies directed against P-gp MDR1 (G-1), BCRP (BXP-21), MRP1 (QCRL-1), and MRP2 (M2 III-6) diluted to a final concentration of 0.5 μg/mL. In parallel, blots were also probed with mouse anti-β-actin antibody at 1:10,000 dilution incubated at room temperature for 1.5 h with gentle rocking. After addition of horseradish peroxidase-labeled goat anti-mouse IgG (1:5,000 dilution), blots were developed by enhanced chemiluminescence and visualized after 20 min exposure using an ImageQuant 350 image analyzer (GE Healthcare).
Optimization of Culture Conditions
BeWo cells were seeded at 30,000 cells/cm2 on 6- or 24-well plates (BD Falcon, BD Biosciences) and at 100,000–125,000 cells/cm2 on 0.6 cm2 Millicell polycarbonate inserts (Merck Millipore). A variety of culture parameters were then evaluated: (1) use of different media formulations (DMEM, F12K, RPMI, or M199), (2) addition of NEAA and/or MEM vitamin solution, and (3) different batches and suppliers of FCS (CSL, Life Technologies). These combinations were also tested with and without culture surface coating (collagen, poly-
Bidirectional Transport
Filter inserts were transferred to fresh 24-well plates for the transport studies. The studies were conducted using an assay medium consisting of Hank's balanced salt solution (HBSS) supplemented with both glucose (Ajax chemicals) and HEPES to give final concentrations of 20 and 10 mM, respectively. The pH was adjusted to 7.4 using 1 M NaOH.
Cells were incubated in a prewarmed assay medium with or without an efflux inhibitor for 30 min at the correct pH, and then rinsed in the same medium. TEER was measured immediately before the start of the experiment and the assay medium with or without inhibitors were placed in the receiver chambers. We used topotecan hydrochloride (20 μM), a typical BCRP substrate, rhodamine 123 (5 μM), a typical P-gp substrate, or C-DCDHF, a paracellular transported compound, added to the donor chamber of each well (apical chamber [Ap] for apical to basolateral studies, and basolateral chamber [Bas] for basolateral to apical studies). The Ap and Bas chambers received 0.3 and 0.6 mL of medium, respectively. Samples were removed from the receiver chamber at various time points over a 3-h period. Constant volumes were maintained by adding the prewarmed medium to the receiver chambers to maintain an equilibrium pressure differential between the volumes in the donor and receiver chambers. The selective BCRP inhibitor fumitremorgin C (5 μM) was used to block functional BCRP activity, while the P-gp inhibitor PSC-833 (4 μM) was also used to provide comparative non-BCRP efflux inhibition data. During the transport study these modifying agents were also present in the donor chamber with our test compounds and in the receiver chamber, at the same concentrations as stated above. Fluorescence was measured in 96-well plates using a FLUOstar Optima fluorescence plate reader. Filters used included a 485-nm excitation filter and a 520-nm emission filter for C-DCDHF and rhodamine 123, while a 355-nm excitation filter and a 520-nm emission filter were used for topotecan detection similar to that used previously (Table 1). 19
Workflow for Optimized BeWo Bidirectional Transport Study
1. 1 mg/mL collagen I dissolved in 0.9 M acetic acid spread over 0.6-cm2 millicel polycarbonate inserts in 24-well plates.
2. Use Class II biological hood under ultraviolet light, aspirate all solution in inserts.
3. Wash off any acetic acid present around the inserts at least three times.
4. Wait overnight before using inserts. Allow to air-dry in sterile hood under ultraviolet light.
5. Generate final concentration of 10% FCS, 2 mM glutamax, and 2 mL of a 100× minimum essential medium vitamin mix solution.
6. Trypsinize cells for 5 min. Need cells from 75-cm2 flask to generate cell density to cover a 24-well study.
7. Allow full day for cells to plate down in humid 37°C incubator with 5% CO2.
8. Add rhEGF at 20 ng/mL from 100 μg/mL stocks to the medium immediately before feeding cells.
9. Maintain cells in 37°C incubator with 5% CO2.
10. Measure TEER using an EVOM meter with an insert probe (EndOHM 12).
11. Replace the medium with HBSS and incubate some wells with the BCRP inhibitor.
12. Add topotecan in HBSS with or without BCRP inhibitors to donor sides and collect aliquots from receiver side for up to 3 h.
13. Apparent permeabilities (Papp) can be determined for each direction of transport from apical to basolateral and basolateral to apical.
BCRP, breast cancer resistance protein; EGF, epidermal growth factor; FCS, fetal calf serum; HBSS, Hank's balanced salt solution; M199, Medium 199; PBS, phosphate-buffered saline; rhEGF, recombinant human epidermal growth factor; TEER, transepithelial electrical resistance.
Data Analysis
Drug transport through cell monolayers was calculated both as a simple amount passing the monolayer per min, which would vary depending on the concentration used in the donor compartment, and as an apparent permeability coefficient as calculated previously.
20
Briefly, our permeability coefficient equation (derived from Eqs. 1 and 2) allows for changes in the concentration of test compound in the donor chamber as the permeability experiment progressed.
where Cl. vol is clearance volume (mL); n, time (min); n
fin, final time point (min); A
a, amount in acceptor compartment at time n (pmol);
Results in this study are presented as the mean±standard error of the mean (SEM). Significant differences between values were examined using the Student's two-tailed unpaired t-test with results considered significant if P<0.05.
Results
Optimization Studies
The culture of BeWo cells over 1 week in the various medium formulations and supplements revealed marked differences in growth. The DMEM was the poorest performing medium with very little growth observed; the RPMI medium was marginally better, but neither resulted in sustained cell growth past 3 days, leaving more than 50% of the available surface unoccupied by day 5 (results not shown). Cells grown in the F12K and M199 medium resulted in the formation of isolated clusters of cells, achieving ∼70% coverage. NEAA supplementation of the F12K medium increased cell growth marginally, while MEM vitamin supplementation also enhanced attachment and growth. No differences were observed between the different batches of FCS used. Subsequent studies employed F12K media with NEAA/vitamin supplementation or the M199 medium with added vitamins.
With respect to coating options, collagen coating promoted denser cell clusters and enhanced confluency (Fig. 1). Gelatin was less effective than collagen, and poly-
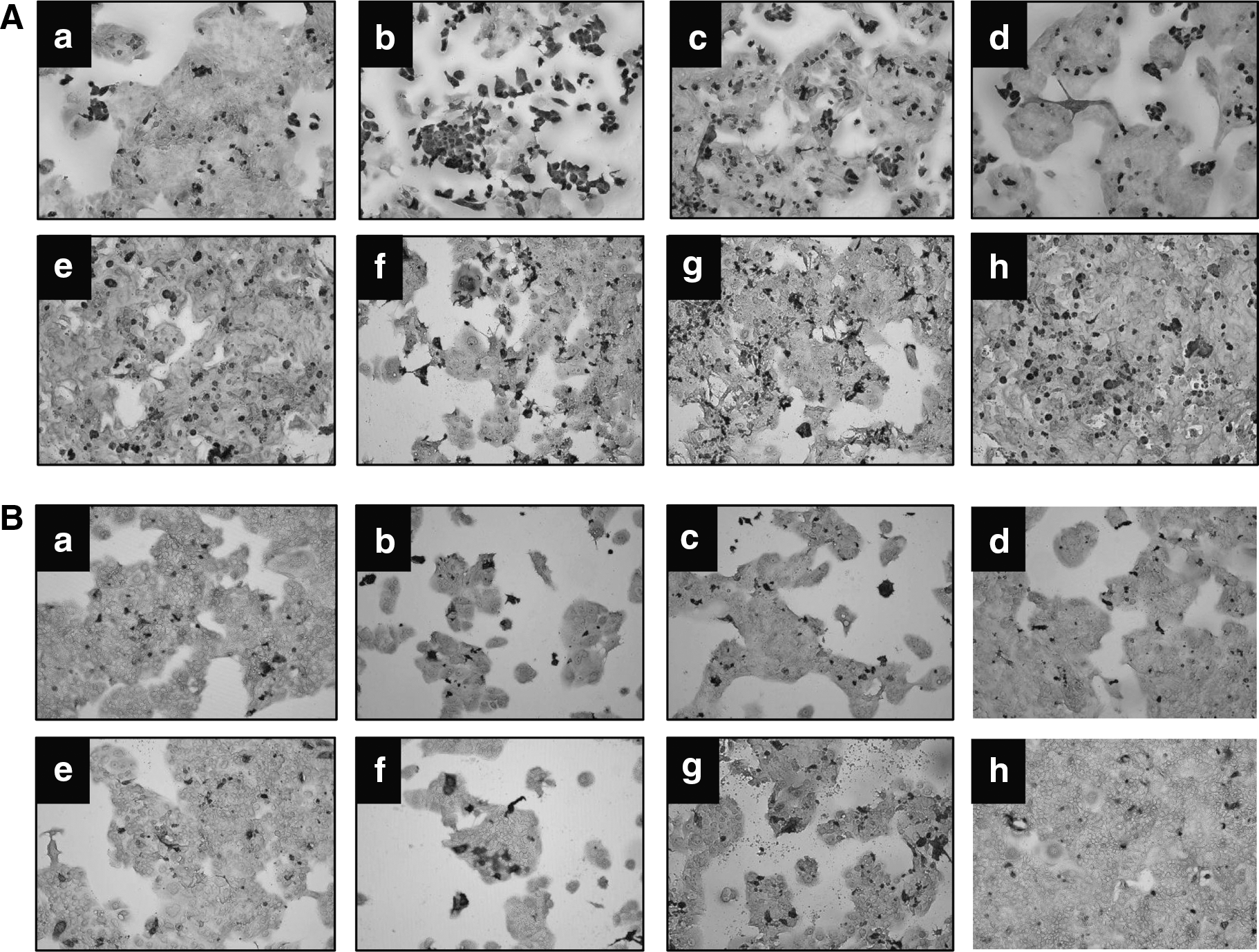
Microscopic analysis of BeWo cell growth after 7 days in varying culture conditions.
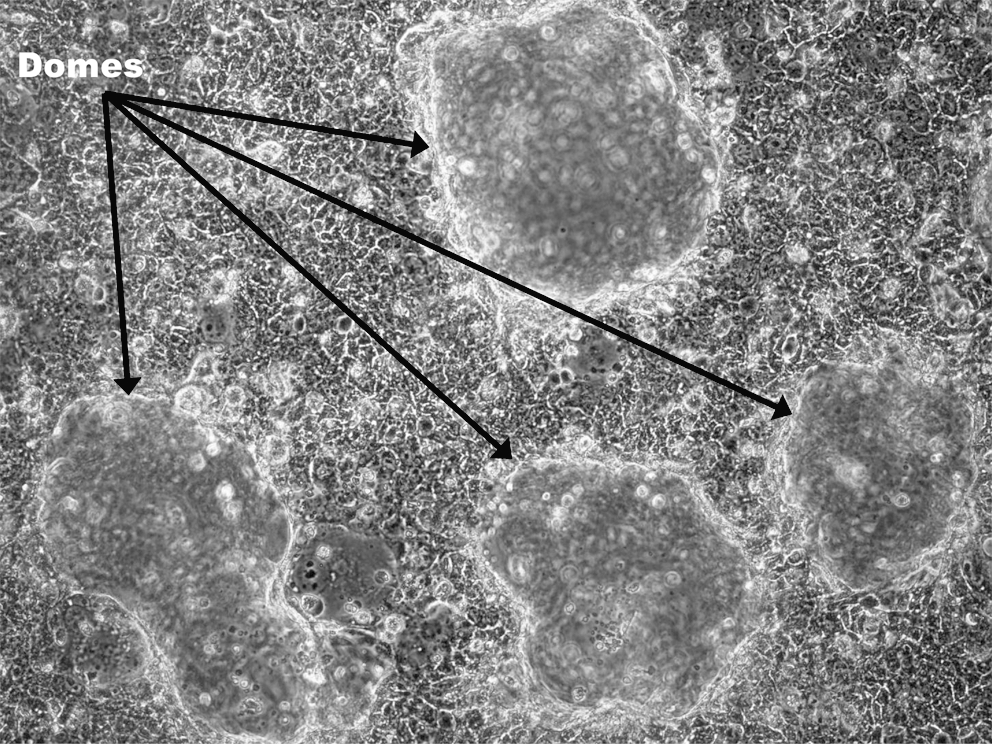
Dome formation in confluent cultures of BeWo cells grown for 7 days in 6-well plates coated with collagen 1 in M199 supplemented with 10 ng/mL EGF and vitamin supplement. 100× magnification on Nikon Ti inverted phase microscope.
Permeability Studies
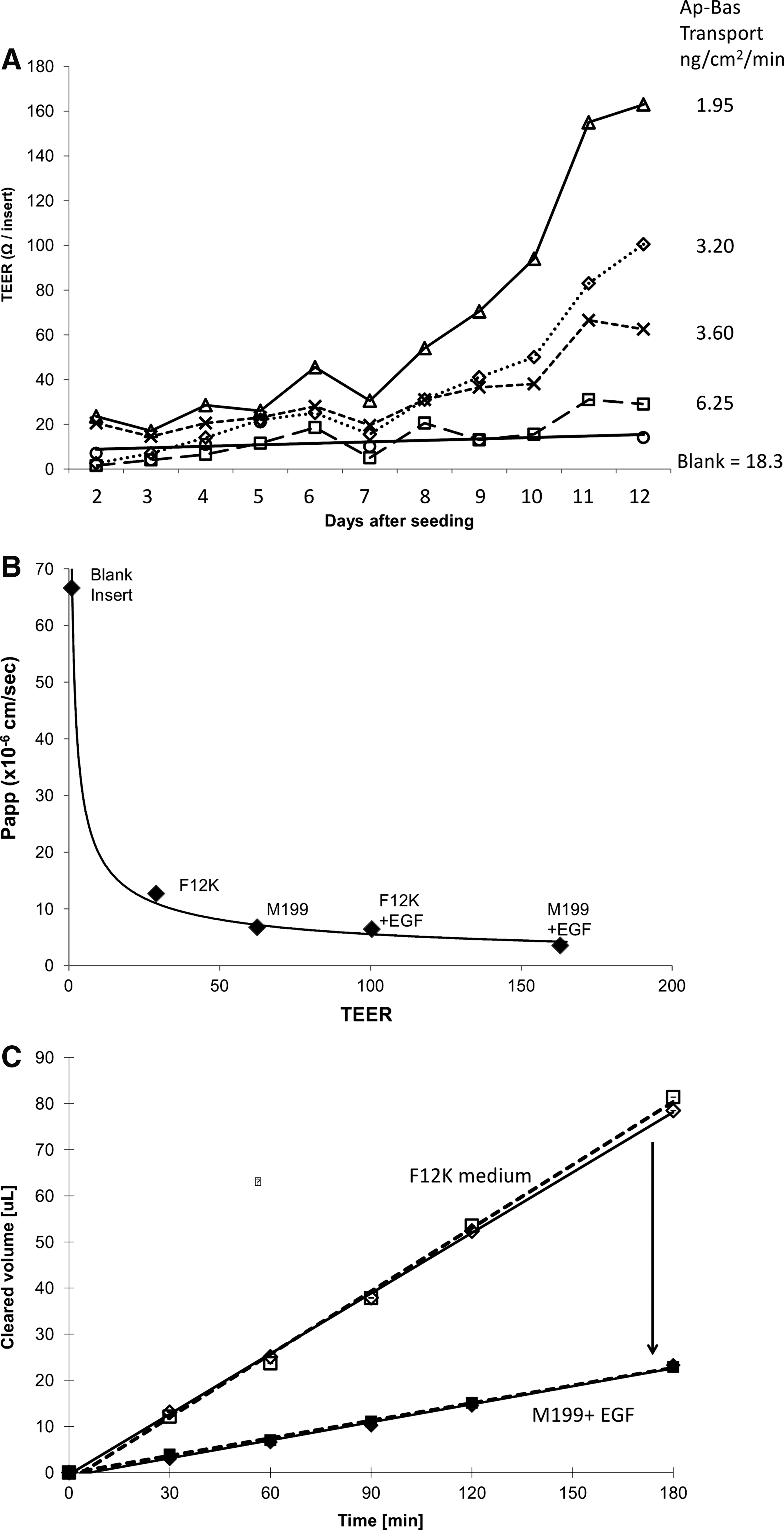
Cells were seeded on inserts at 100,000–125,000 cells/cm2, and TEER was measured daily for 12 days to assess the effects of culture optimization on cellular membrane permeability. Cell cultured at 100,000 cells/cm2 exhibited variable TEER measured between replicates of 30%. Therefore, a higher cell density of 125,000 cells/cm2 was chosen, resulting in more consistent tight junctions between replicates. Cells grown in the M199 medium plus epidermal growth factor (EGF)/vitamins on collagen-coated inserts clearly exhibited much higher TEER values than the other growth conditions (Fig. 3A), with readings above 165 Ω recorded. After 12 days, TEER values plateaued (data not shown), suggesting that optimal tight junction formation had been reached. Transport of the hydrophilic paracellular transport marker C-DCDHF (23 μM) was assessed on day 12 of culture. Ap-to-Bas transport rates were 18.3 ng/(cm2·min) (66×10−6 cm/s) in nonseeded collagen-coated inserts (i.e., no BeWo cells), 3.2 ng/(cm2·min) (6.0×10−6 cm/s) with BeWo cells cultured on collagen-coated inserts in the F12K medium plus EGF, and 1.9 ng/(cm2·min) (3.5×10−6 cm/s) with cells grown in the M199 medium plus EGF (Fig. 3B, C).
Topotecan hydrochloride is a known BCRP substrate. Bidirectional transport studies through confluent BeWo cells resulted in Ap to Bas transport of 6.0×10−6 cm/s (2.77 ng/[cm2·min]), while Bas to Ap transport was 7.8×10−6 cm/s (3.67 ng/[cm2·min]) (Fig. 4). Each transport analysis was done in triplicate (Fig. 4); error bars representing SEM values are shown, but due to their size are obscured and difficult to see. The difference in the rate of Ap versus Bas topotecan transport was significant with P<0.005. Addition of fumitremorgen C, a potent BCRP inhibitor, abolished the difference in transport direction (P<0.773) (Fig. 4). The P-gp inhibitor PSC-833 was also used, but Bas to Ap transport remained significantly higher than Ap to Bas transport (P<0.024). To illustrate specificity for BCRP transport, additional studies using the P-gp substrate rhodamine 123 were performed. Transport of rhodamine 123 through BeWo cells layers was shown to be 6.9×10−6 cm/s in both direction without the addition of any inhibitors (Fig. 5) (P<0.967). The addition of 4 μM PSC-833 did not significantly alter transport rates (P<0.499), nor did the use of fumitremorgin C (P<0.889), suggesting that P-gp substrates are not actively transported in this model.
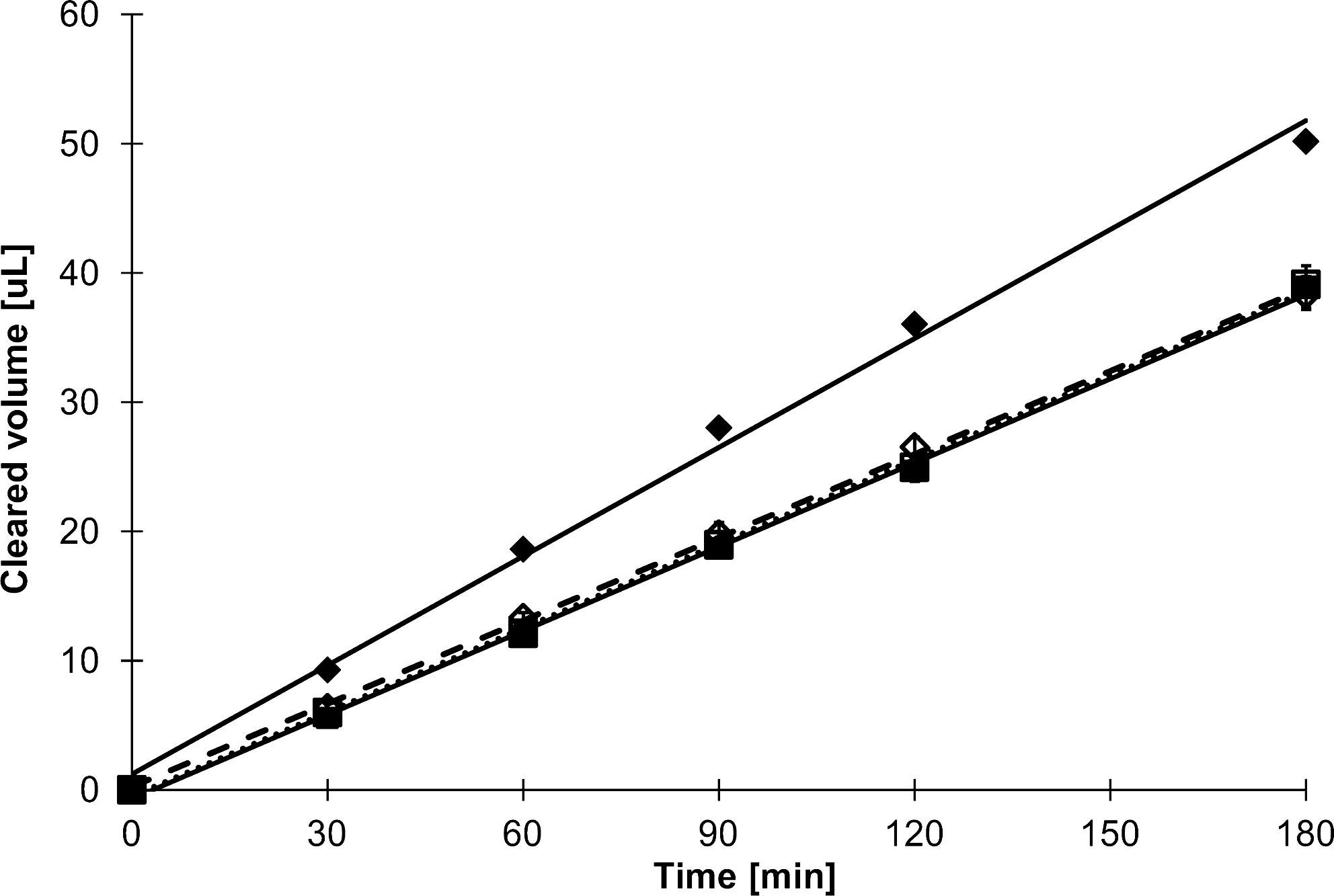
Bidirectional transport of topotecan hydrochloride (20 μM) by BeWo cells grown on 0.6-cm2 polycarbonate inserts at day 14 of culture. Cells were maintained in simple balanced salt solution for the 3-h study. Fumitremorgin C (5 μM) or vehicle was added 30 min before the start of the experiment and was maintained in both Ap and Bas chambers. Results are shown for topotecan hydrochloride alone (■, ⧫) or with fumitremorgin C (□, ◊). Results are shown for both Ap to Bas (□, ■) and Bas to Ap (◊,⧫) directions for each culture condition. Data shown are mean±SEM (n=3). Error bars may not be visible due to their small size. SEM, standard error of the mean.
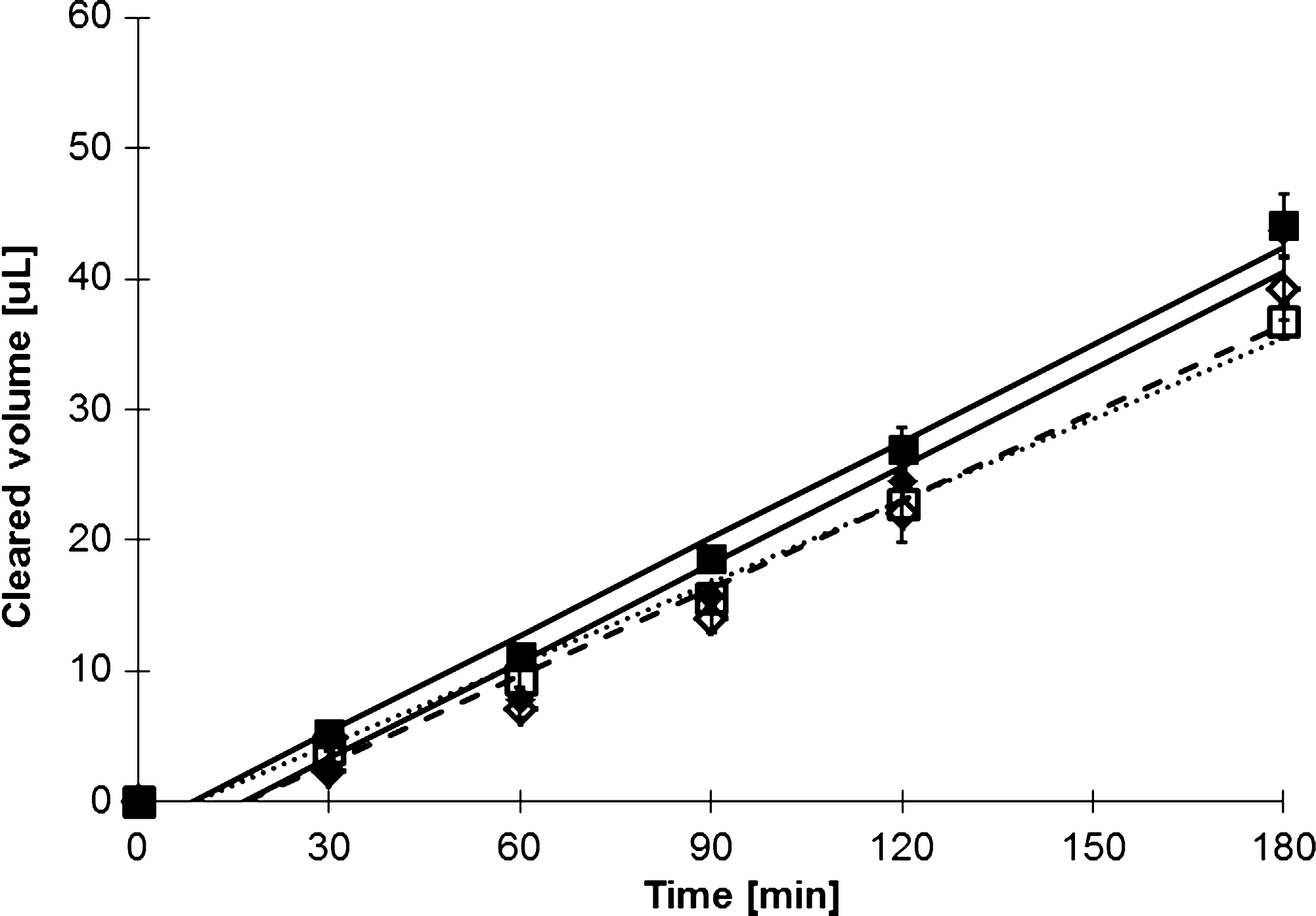
Assessment of bidirectional transport of rhodamine 123 (5 μM) in BeWo cells grown on 0.6-cm2 polycarbonate inserts at day 15 of culture. Cells were maintained in simple balanced salt solution for the 3-h study. P-gp inhibitor PSC-833 (4 μM) or vehicle was added 30 min before the start of the experiment and was maintained in both Ap and Bas chambers. Results are shown for rhodamine 123 alone (■, ⧫) or with PSC-833 (□, ◊). Results are shown for both Ap to Bas (□, ■) and Bas to Ap (◊,⧫) directions for each culture condition. Data shown are mean±SEM (n=3). Error bars may not be visible due to their small size. P-gp, P-glycoprotein.
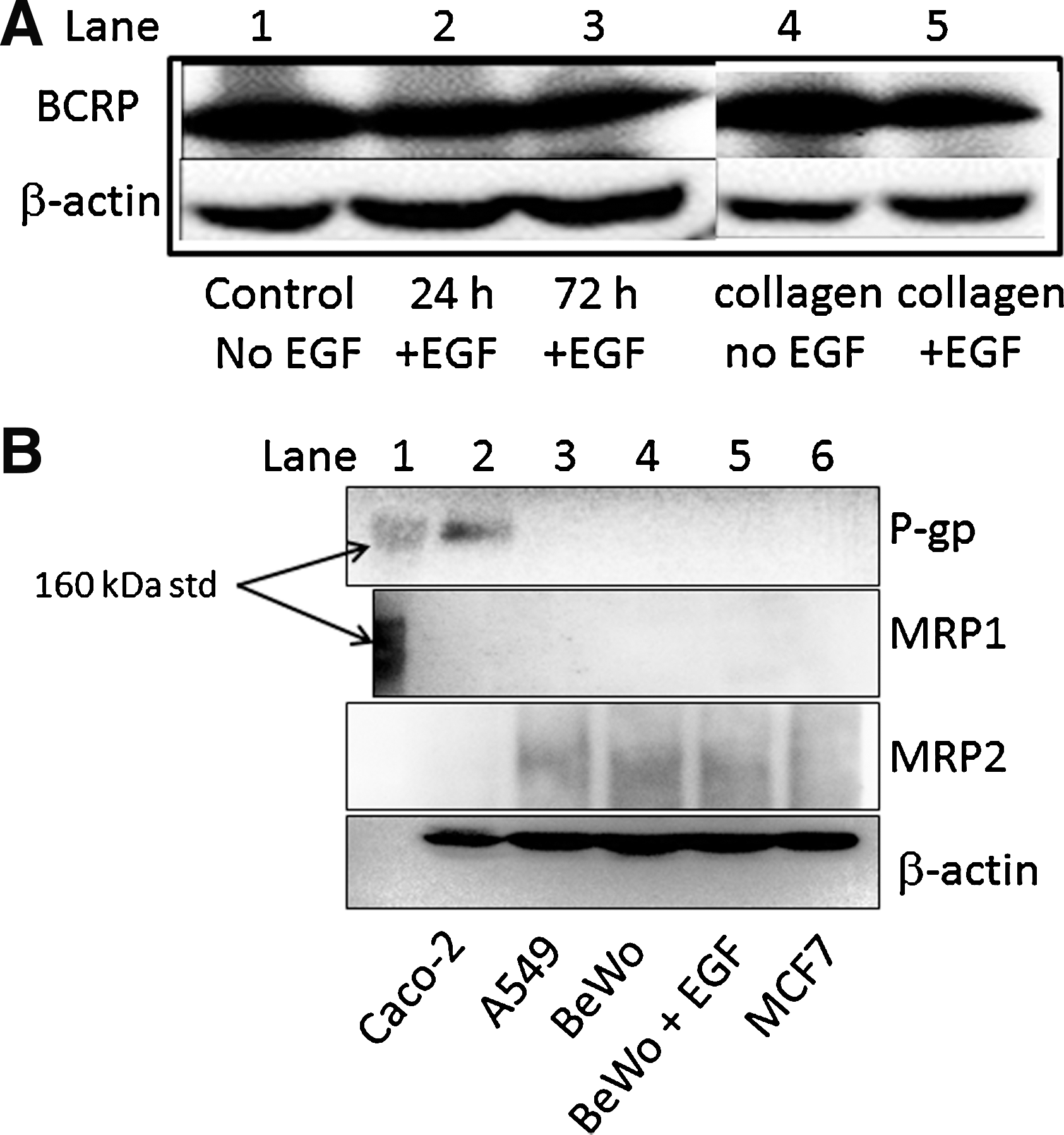
Western blotting was performed to identify levels of ABCB1 (P-gp), ABCG2 (BCRP), ABCC1 (MRP1), and ABCC2 (MRP2) proteins. BCRP was readily detected in the BeWo cells used in our study (Fig. 6A). No visible bands were detected for P-gp in the BeWo lysates under any experimental conditions, consistent with previous studies 21 (Fig. 6B). MRP1 was also not detected, but MRP2 was shown to be expressed, although the strength of the signal was not as strong as that for BCRP. There were no changes in BCRP abundance in the presence or absence of culture factors used to generate confluence and monolayer formation in this cell line.
Discussion
The BeWo cell line has been described in many studies of trophoblast biology, differentiation, and invasion, but seldom used in drug transport studies despite the fact that it expresses a number of functional drug transport proteins, including BCRP. 16 This is almost certainly due to its propensity for aggregation and fusion, which creates large gaps on the culture plate, precluding formation of an intact monolayer capable of effective membrane barrier function. 17,22 Most investigators employ the B24 BeWo clone for their studies, which is available commercially from public cell depositories such as American Type Culture collection (ATCC). Although an early report described (B24) BeWo cell cultures able to produce TEER values of 600 Ω·cm2, 23 suggestive of very effective tight junctions, this has not been subsequently replicated and it is widely believed that BeWo cells do not form confluent monolayers. However, one group of researchers, the Schwartz group in Washington, have described a specific BeWo subclone, B30, capable of forming confluency. 22 They have conducted several studies using this clone, including studies on iron 17 and linoleic acid transport. 18 Unfortunately, although a few other groups in Europe and the United States have subsequently used this subclone, there is some variability in the effectiveness of tight junction formation from these other groups. 24,25 Recently, one group from Europe attempted to generate monolayers from the B30 subclone of BeWo cells and were not able to get the tight junctions to increase beyond 14 Ω·cm2, with high fluorescein transport rates equivalent to 10% per hour, 24 while another study showed tight junctions to be around the 45 Ω·cm2 level, and 4% fluorescein transport per hour, 25 suggesting that there can be high variability in tight junction formation even with use of a subclone that has a higher propensity to generate monolayers. In addition, approval to use this subclone must come from specific research labs in the United States, as it is not held in commercial cell banks. In contrast, the B24 clone is widely distributed from official American and European cell line distribution sites, which allows ready worldwide access to laboratories interested in developing a BCRP transport model. Here we have outlined a specific set of culture conditions using standard B24 cells that facilitate the formation of confluent BeWo monolayers with tight junctions that will be useful for the study of trophoblast permeability, substrate specificity, and transport kinetics of efflux proteins such as BCRP/ABCG2 in vitro.
We evaluated a variety of culture conditions to arrive at a combination of medium, supplements, and culture surface coating needed to achieve confluency. We optimized the culture medium and vitamin content to achieve optimal growth. We also evaluated the effects of increases in intracellular cyclic adenosine monophosphate (cAMP) levels to promote differentiation and ERK1/2 and p38MAPK phosphorylation 26 –28 based on the reported effects of cAMP on tight junction formation between adjoining BeWo cells. 27 However, this strategy merely promoted aggregation and fusion with no benefits on confluence. We also evaluated addition of EGF since this is known to increase cell growth, ERK1/2 and p38MAPK activity, and α2-integrin expression. 29 The addition of EGF proved extremely successful in terms of promoting growth and confluence, particularly at the higher dose (20 ng/mL). It is important to note that EGF was not added until 24 h after seeding to allow normal attachment before growth stimulation, with medium and supplements added every 48 h. In our hands, 12 days of constant exposure to EGF were required to reach peak tight junction formation. Coating with collagen I was also essential for formation of BeWo monolayers in our study. This is despite the findings of Nakatsuji and coworkers that EGF treatment had no impact on the adhesion of BeWo cells to collagen IV coated plates. 29 Our observations are unlikely to be related directly to adhesion; it is more likely that cell-to-cell apposition is affected. The differences in EGF response could reflect a difference in the BeWo phenotype when grown on type I and type IV collagen. 29
The use of TEER measurements as a marker of barrier formation is a common technique, but subject to considerable laboratory-to-laboratory variance due to lack of consistency and reproducibility in measurement techniques and calibration. The original 1997 study detailing the transport properties of the B30 sub-clone showed fluorescein transport at 10×10−6 cm/s and TEER values around 60 Ω·cm2. 18 In a more recent publication using the B30 clone, Heaton and coworkers reported TEER measurements over 300 Ω·cm2, but with unexpectedly high rates of transmission of mannitol of 3.5% per hour. 17 Under our optimal growth conditions, BeWo cultures achieved TEER readings of 170 Ω, 8.5× greater than background, while the carboxyfluorescein transport rate was only 1.6% per hour (3.5×10−6 cm/s). These TEER inconsistencies may be due to methodological differences, e.g., the use of chopstick electrodes or endohm chambers to measure TEER. We have found that chopstick electrodes can provide inconsistent and variable data (>100 Ω between measures), so now we use fixed chambers that provide more accurate and reproducible results.
Stability of the transport characteristics of the model is an important consideration. In a previous study using the B30 sub-clone on collagen-coated polycarbonate inserts, confluency could not be maintained for more than 60 min. 30 In contrast, our model showed linearity in efflux for at least 3 h in BeWo cells maintained in a simple buffered balanced salt solution. Drug transport in polarized epithelia would be expected to exhibit marked differences in directionality. In our model although the A–B versus B–A efflux differential for the BCRP substrate topotecan was significant, it did not exceed 1.3-fold. Criteria established for P-gp-mediated efflux in Caco-2 cells suggest that transport differential should be at least two-fold to achieve a functional model with useful characteristics. 31 However, this assumes that the extent of polarized expression of the transporters of interest is similar to that of P-gp. The polarity of BCRP expression in BeWo cells has not been assessed, and is currently assumed to be similar to that of the placenta with the majority facing the apical side, although recent findings suggest that BCRP is also present on the maternal side of the placenta. 2 As our data showed some increased basolateral efflux of topotecan, and use of BCRP inhibitors only decreased Bas to Ap transport. If BCRP was present on both sides of the cells, use of a BCRP inhibitor would have decreased transport in both directions, and this was not observed. In addition, we have also shown that this model does not exhibit P-gp-mediated transport, as no P-gp protein was detected and bidirectional transport studies with a P-gp substrate found no evidence of P-gp-mediated efflux.
Therefore, in this study we have shown that in appropriate culture conditions, standard BeWo cells can generate monolayers with tight junctions for studies of bidirectional transport of high-affinity BCRP substrates and inhibitors. For low-affinity molecules, increases in the polarity of expression or the cellular barrier integrity would be required to provide an adequate level of sensitivity. Measurement of intracellular substrate concentrations might also enhance the utility of the model. While the potential presence of MRP2 must be taken into consideration when using this cell line as a model to study BCRP transport, the absence of P-gp or MRP1 provides a distinct advantage. In addition, the study of other transport proteins and processes, such as the transport of amino acids, fatty acids, and nanoparticles, could also be facilitated by the use of this model.
Footnotes
Acknowledgment
We thank Erin Bolitho from the Pharmacogenetics division within the School of Pharmacy at Curtin University for routine screening of the BeWo cells for mycoplasma and for the preparation of stock concentrations of poly-
Disclosure Statement
No competing financial interests exist.