
Abstract
Recombination-based cloning techniques have in recent times facilitated the establishment of genome-scale single-gene ORFeome repositories. Their further handling and downstream application in systematic fashion is, however, practically impeded because of logistical plus economic challenges. At this juncture, simultaneously transferring entire gene collections in compiled pool format could represent an advanced compromise between systematic ORFeome (an organism's entire set of protein-encoding open reading frames) projects and traditional random library approaches, but has not yet been considered in great detail. In our endeavor to merge the comprehensiveness of ORFeomes with a basically simple, streamlined, and easily executable single-tube design, we have here produced five different pooled screening-ready libraries for both Staphylococcus aureus and Homo sapiens. By evaluating the parallel transfer efficiencies of differentially sized genes from initial polymerase chain reaction (PCR) product amplification to entry and final destination library construction via quantitative real-time PCR, we found that the complexity of the gene population is fairly stably maintained once an entry resource has been successfully established, and that no apparent size-selection bias loss of large inserts takes place. Recombinational transfer processes are hence robust enough for straightforwardly achieving such pooled screening libraries.
Introduction
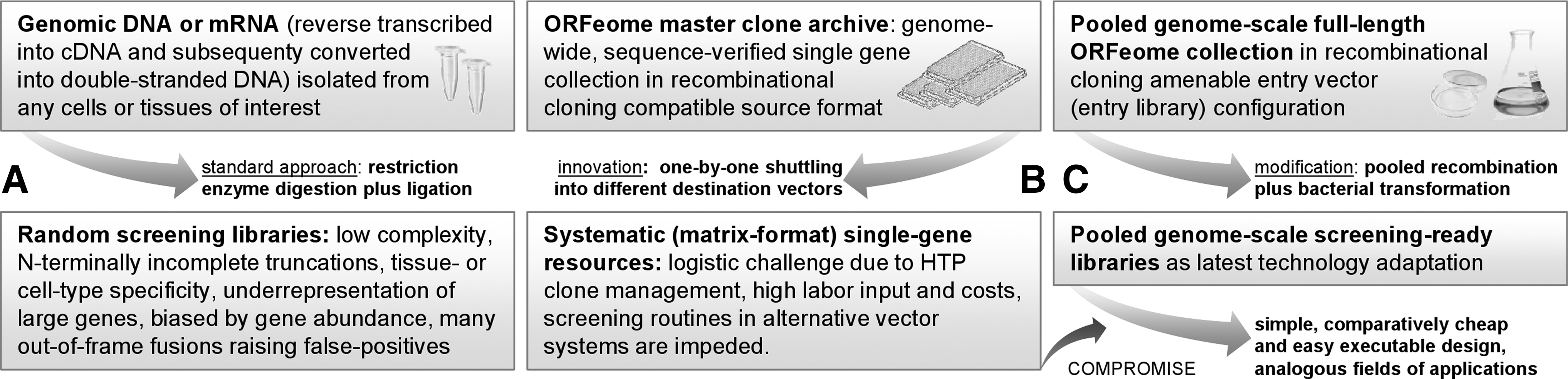
Comparison of our pooled library arrangement to traditional, random library construction approaches and the original ORFeome concept.
Materials and Methods
Cloning of Pooled Staphylococcus aureus PCR Products into pDONR™-zeo by Recombination Between attB and attP Sites
Equal volumes of 2,562 unique attb-site-flanked PCR products each corresponding to a defined full-length ORF originating from our recently published S. aureus strain Mu50 ORFeome construction project 10 were pooled (resultant stock concentration: 225 ng/μL) and without any further purification directly inserted into Gateway® entry vector pDONR-zeo (Life Technologies™, Grand Island, NY) by standard recombination between attB and attP sites (referred to henceforth as BP). Two independent 10 μL reactions each containing equimolar ratios of 150 ng insert DNA, 150 ng vector, and 2 μL BP clonase™ II mix were performed and incubated at room temperature for 20 h. The mixture was pooled, purified via Illustra's GFX™ PCR DNA and Gel Band purification kit (GE Healthcare, Uppsala, Sweden), transformed into two aliquots (100 μL) of electrocompetent Top10 Escherichia coli cells (theoretical efficiency: 109 CFU/μg supercoiled DNA; Life Technologies), and propagated in the liquid low-salt LB medium supplemented with 25 μg/mL Zeocin™ (Life Technologies). Plasmid DNA of the resultant S. aureus ORFeome entry library was prepared using the GenElute™ HP Plasmid Maxiprep kit (Sigma Aldrich, St. Louis, MO).
Pooling and Amplification of a Human Entry Gene Collection
The commercially available Homo sapiens ORFeome v1.1 contains 8,270 full-length, sequenced ORFs (
High-Efficiency Transfer of Donor Libraries in Various Destination Plasmids
Attachment-site-tagged S. aureus and human entry library pools were in a simple, single-tube reaction shuttled into the recombination-compatible destination vector pADgate2 (in-house adapted from Clontech's yeast two-hybrid [Y2H] prey vector pGADT7 via Life Technologies, Gateway vector conversion system) and in isopropyl β-D-1-thiogalactopyranoside–inducible bacterial expression vectors pDEST™15 (equipped with a glutathione S-transferase [GST]-tag; Life Technologies) and/or pDEST™17 (carrying a 6xHIS tag) using the Gateway LR clonase II Plus mix (Life Technologies) according to the manufacturer's protocols, except that a 6× scale was used and recommended proteinase K digestion was omitted. Reactions were purified by running through Illustra's spin columns (GE Healthcare), eluted in 12 μL 10 mM Tris-HCl, pH 8.0 each, and completely transformed into 100 μL electrocompetent Top10 E. coli cells. For selection of successfully recombined vectors, cells were plated on 10 large (145 mm diameter; Nunc, Roskilde, Denmark) LB plates supplemented with 100 μg/mL ampicillin. After estimation of overall efficiency of both in vitro recombination and bacterial transformation, clones of two plates each were scraped from solid-surface agar, and plasmid DNA was recovered using the GenElute HP Plasmid Maxiprep kit (Sigma-Aldrich).
RTq PCR Evaluation of Recombination (Shuttling) Efficiencies
Accession IDs of some randomly chosen, differentially sized (Table 1) genes known to be included within either our S. aureus PCR product pool or the human ORFeome v1.1 were subjected to Batch Entrez (
List of Staphylococcus aureus and Homo sapiens Genes Randomly Chosen Based on Three Different Coding Sequence Size Categories (Small, Medium, and Large) and Specific Primer Pair Combinations Used in Quantitative Real-Time Polymerase Chain Reaction Studies to Validate Shuttling Efficiencies
Antibiotic resistances employed for data normalization are included at the bottom.
Results and Discussion
Pooled ORFeome Libraries Can Be Efficiently Shuttled Without Loss of Gene Content
Genome-wide protein–protein interaction studies via the classical Y2H technology, for instance, in array (matrix) reminiscent attitude wherein a set of ORFs is cloned as both bait (fused to a transcription factor DNA-binding domain) and prey (attached to an activation domain) constructs and next introduced into isogenic reporter strains of opposite mating type crucial for the selection of diploids that are able to grow on selective medium, are among the most widespread downstream areas of operation of systematic genome-wide ORFeome collections. 11 Alternatively, recombinantly expressed and E. coli–purified ORF clones—equipped with well-characterized affinity tags (6xHIS, GST, c-myc, and Streptag) or other functional moieties (e.g., green fluorescent protein)—serve as highly versatile fundaments for diverse functional HTP-profiling studies (in vitro interaction assays, drug target spectrum delineation, bioimaging, crystallography, antibody generation, etc.), with most valuable hints for basic biology, assay optimization plus prioritization of novel therapeutics alike. 4,12,13 At this juncture, our motivation was to merge the apparent benefits of ORFeome construction achievements with more user-friendly applicability profiles (Fig. 1). Starting either with bacterial genomic DNA-derived PCR products of 2,562 full-length, protein-encoding genes (reflecting an altogether 95% ORFeome coverage) flanked on both sides with site-specific (attB) recombinase recognition sites as in the case of S. aureus or directly with a pooled human ORFeome reverse proteomics platform comprising 8,270 full-length constituents, we first of all produced two different organism-specific pooled (genome-scale) ORFeome entry libraries, amplified them, and finally shuttled respective inserts—following a sixfold upgraded recombination protocol—into a screening-ready Y2H prey collection and a GST- and/or HIS-tag recombinant protein expression library each. Best possible reaction conditions, as determined by average transformation efficiency (routinely >7×106 individual clones per single library, thereby extensively covering the enclosed number of full-length ORFs) herein encompassed equimolar insert to vector ratios, a 20-h (overnight) reaction time, electroporation of the entire (purified) recombined mixtures in two aliquots of electrocompetent E. coli cells, and five plasmid preparations recovered thereof (data not shown in detail). Given the well-known high percentage of 5′ truncated constructs attributable to premature halts in reverse transcription of template mRNA and—despite laborious enrichment efforts, such as cDNA size-fractionation before PCR amplification 14 or 5′ cap structure biotinylation followed by biotin capture of full-length cDNAs 15 —the critical underrepresentation of large genes inherent to inefficient synthesis, transmission (ligation), and/or propagation in traditional genomic versus cDNA libraries, 16 one rational concern about using our straightforward pooled ORFeome library configuration was the explicit maintenance of specific gene content after recombination between different vector backbones. Therefore, RTq PCR of several randomly selected S. aureus and human genes of three different size categories (small, <480 nt for S. aureus and <620 nt for H. sapiens; medium, <1,080 nt/<1,501 nt; large: <2,200 nt/<4,400 nt) was performed before and after transfer in technical triplicates, each to appraise the reliability and hence overall quality of resultant gene collections (pooled ORFeome libraries). As gets apparent from Figure 2 that summarizes quantitative data on achieved shuttling efficiencies in the form of passed threshold cycle values (Cts), the initial BP recombination between attB-site-flanked PCR products or DNA segments and an attP containing the donor constitutes the most rate-limiting step. For instance, had both S. aureus SAV1569 and SAV2392 been lost during entry library preparation, a bottleneck perhaps partly circumventable through the future use of 6× formats was subsequently pursued for all LR shuttling procedures. Once a comprehensive entry resource (regardless of the original starting material) is at hand, the complexity of the ORF population is, however, fairly stably maintained as gets apparent by the fact that all constituents available within our S. aureus and H. sapiens entry libraries are similarly covered by resultant Y2H and affinity-tagged protein expression (destination) libraries. Assuming that entry library ct values with their inverse proportionality to respective target gene amounts symbolize a hypothetical 100% gene content, accurate yields averaged over all screening-ready ORF collections (excluding the two staphylococcal genes lost during BP reaction) comprised 100.2%±3.15% for small, 102.8%±4.2% for medium, and 101.1%±5.4% for large S. aureus genes and, likewise, 103.5%±44.9% for small, 98.79%±2.03% for medium, and 100.0±4.4% for large H. sapiens genes. Of particular note, even large inserts, ranging from ∼2,000 nt (as is the case for S. aureus SAV1220 or SAV1634) up to 3,476 nt (human NRIP1) or actually 4,394 nt (COL1A1), were consistently transferred with comparable efficiencies. These observations indicate that the in vitro recombination process at pooled library scale does not show any destructive size-selection bias between entry and destination libraries, and hence does not negatively impact the complexity of resultant gene collections as might have been formerly anticipated. Given that a library's quality plus diversity symbolizes the most critical basis for various follow-on studies, our work proves that transfer processes are robust enough for straightforwardly and rapidly achieving such comprehensive screening collections.
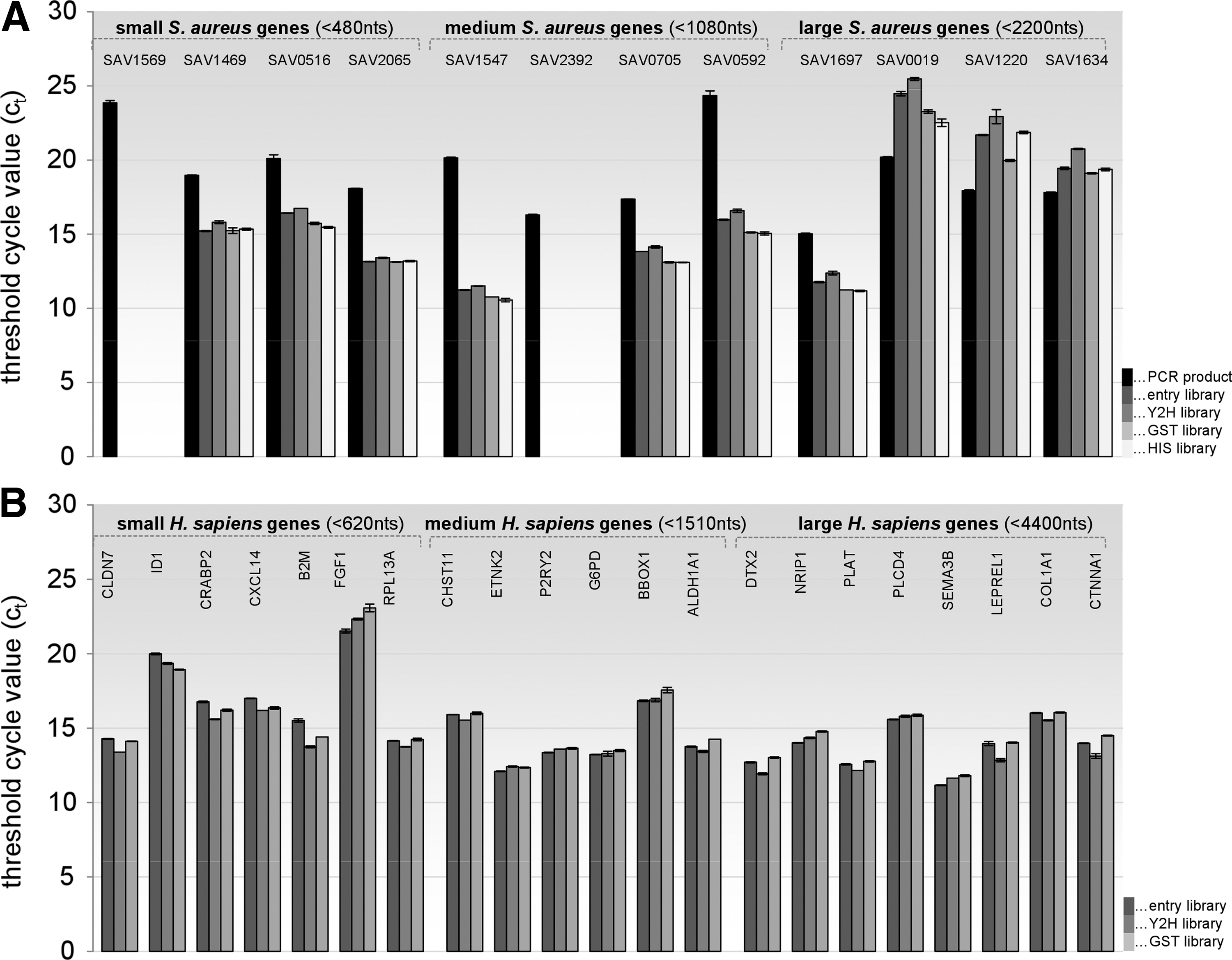
Shuttling efficiencies for differentially sized Staphylococcus aureus
HTP Application Schemes Enabled by Comprehensive ORFeome-Wide Screening Collections
One conceivable area of operation of such screening collections is as prey libraries in the field of Y2H assays that permit the unbiased HTP screening for novel protein–protein interactions. 17 –19 By representing more correct in-frame-fused candidate genes and offering a more complete search space, ORFeome-derived Y2H screening libraries can not only be supposed to significantly outperform classical Y2H libraries, but also might yield unexpected associations that could otherwise be missed or overlooked in more limited focused (biased) arrangements. According to a short literature survey, several applications have already been successfully attempted with recombined pooled (yet in part initially total RNA/reverse transcribed cDNA derived) gene libraries. Examples encompass the creation of Arabidopsis thaliana 20 or Brachypodium distachyon 21 entry resources and their subsequent transfer in Y2H destination plasmids, and the screening of pooled Kaposi's sarcoma-associated herpesvirus versus 688 Caenorhabditis elegans ORF collections (screening colonies of pooled ORFeomes SCOOP; pooled ORF expression technology POET) for soluble recombinant protein (over)expression. 6,7 Combining an organism's entire number of full-length protein-encoding ORFs into one single pool for subsequent entry and screening-ready destination library construction can evidently reduce required experimental steps (recombination reactions, E. coli transformation for plasmid propagation, plating, colony picking, plasmid isolation, etc.) to a mandatory minimum. Because of the reliability and maintenance of quality standards of resultant screening-ready libraries as evaluated by our RTq PCR validation of shuttling efficiencies, this strategy can be readily utilized for establishing large (even genome-wide) gene collections that might permit a ready broadening of numerous proteomics techniques to more global, systems level-inspired dimensions.
Footnotes
Disclosure Statement
No competing financial interests exist.