
Abstract
Spinal muscular atrophy (SMA) is a severe autosomal recessive disorder affecting one in every 10,000 live births. The disease is characterized by loss of alpha-motor neurons in the spinal cord that leads to progressive atrophy and weakness of limb and trunk muscles. This neuromuscular disorder results from deletions and/or mutations within the survival motor neuron 1 (SMN1) gene, leading to a pathologically decreased expression of functional full-length SMN protein. Here we report on the investigation to measure SMN protein levels through electrochemiluminescence immunoassay (ECLIA). This simple assay is a highly quantitative method able to measure SMN protein levels in human, mouse, and rat samples throughout a wide working range with low intra- and interassay error. The sensitivity for human SMN is 30 pg/mL and provides a new tool for the set up of high-throughput screening for basic research. Moreover, we describe a novel tool for a noninvasive assessment of SMN in buccal cells derived from healthy donors, SMA carriers, and SMA patients. The availability of a validated quantitative ECLIA should improve the investigation of novel compounds for the treatment of SMA.
Introduction
Spinal muscular atrophy (SMA) is an autosomal recessive neuromuscular disorder characterized by hypotonia, progressive proximal muscle weakness, and respiratory distress that result from sequential degeneration of alpha-motor neurons in the anterior horn of the spinal cord. 1 Although carrier rates vary considerably among different ethnic groups, 2 SMA is one of the most common monogenetic disorders, with an incidence of 1 in 6,000 to 1 in 10,000 live births, 3,4 and a leading genetic cause of infant mortality. Based upon the severity of its clinical manifestations, namely the age of disease onset and the motor milestones achieved, the spectrum of SMA can be subdivided into four types. 5 In the most severely affected patients (Type I), symptoms manifest by 6 months of age and usually lead to death by respiratory failure before age 2, whereas Type II and III patients present with first symptoms after the age of 6 months and develop the ability to sit (Type II) or walk (Type III), respectively. Comparatively, mildly affected patients with a disease onset after the age of 30 and a normal life expectancy are classified as Type IV SMA.
In 1995, the survival motor neuron 1 (SMN1) gene was identified as an SMA-determining gene, 1 and it has since been shown that homozygous deletion or mutational inactivation of SMN1 is causative of all four types of SMA. The broad clinical spectrum of the disorder can, in part, be explained by its unique genetic background: the SMN1 gene is located within a duplicated and inverted 500 kb chromosomal segment on 5q13 that comprises a neighboring nearly identical gene called SMN2. The two genes differ in five base pairs only, all of which are located toward their 3′ ends, 1,6 and they are both ubiquitously expressed. Full-length transcripts derived from both SMN1 and SMN2 encode an identical 294 AA product, the SMN protein. However, due to a critical C-to-T transition at position +6 of exon 7 of the SMN2 gene, only about 10% of its transcripts are correctly spliced full-length mRNAs, while the vast majority of transcripts derived from SMN2 lack exon 7, 5,7,8 resulting in the translation of a truncated SMNΔ7 protein that is highly unstable and cannot functionally replace SMN. 9,10
Although the homozygous inactivation of SMN1 in SMA patients cannot be fully compensated for, a small amount of functional SMN protein is still produced by the SMN2 gene. Consequentially, an increased number of SMN2 copies is thought to moderate the lack of SMN protein, thus reducing SMA severity, which explains why SMN2 copy number has been shown to inversely correlate with the clinical severity of the disorder. 11 –13 In SMA patients, the SMN2 copy number can reach up to eight and usually varies between 1 and 4 copies, 12 whereas a complete loss of SMN2 in an SMN1 null genome is embryonically lethal in humans 14 —in other words, every SMA patient possesses at least one copy of SMN2 and is thus expected to produce a basic level of SMN protein.
Given the particular genetics that gives rise to SMA, enhancing the production of full-length SMN protein from the SMN2 gene, which is present in all patients, has become a major therapeutic approach. 15,16 For any drug discovery program based on that mode of action, a key prerequisite is the reliable and accurate quantification of SMN protein levels in various fluids, cells, and tissues. Moreover, as some of the preclinical drug candidates eventually enter clinical testing, these quantification methods also need to perform rapidly and on a high-throughput basis. To meet these criteria, different SMN-specific enzyme-linked immunosorbent assays (ELISAs) have been developed during the past years that allow for both SMA drug screening and SMN measurement in human and mouse tissue samples, including peripheral blood mononuclear cells (PBMCs). 16 –19
Although ELISAs can detect much lower protein quantities than classical western blotting (dynamic range of 50–3,200 pg/mL), 19 in terms of sensitivity they are outperformed by a recently developed detection method called electrochemiluminescence immunoassay (ECLIA) whose principle of function is described elsewhere. 20,21 The aim of this study was the development and verification of this high-throughput assay for simple and accurate quantification of SMN protein levels that allows a highly sensitive detection limited to 30 pg/mL.
Materials and Methods
All the samples used in this study were taken after the participants provided their consent by signing a statement of agreement. The SMA carrier provided written informed consent before providing buccal cells used in this study. For the SMA patient, we obtained written informed consent from his guardian.
Antibodies
We used a primary mouse SMN monoclonal antibody as the capture antibody from Sigma-Aldrich (clone 2B1, #S2944) and a secondary SMN rabbit polyclonal antibody from Proteintech (#11708-1-AP); the goat anti-rabbit HRP-Sulfo TAG™ antibody from Meso Scale Discovery (MSD) was applied as a detection antibody. The recombinant human SMN1 standard protein was purchased from Enzo Life Sciences (#ADI-NBP-201-050).
Cell Culture
The cell lines used for the experiments were F11 (a fusion product of the mouse neuroblastoma cell line N18TG-2 with embryonic rat dorsal root ganglion neurons), 22 human Schwann cells (hSCs, kindly provided by Prof. Ahmet Höke, John Hopkins University), 23 and an SMA fibroblast cell line (#GM22592; Corriel Cell Repositories). The cells were maintained in Dulbecco's modified Eagle's medium, including high glucose and stabilized glutamine (Invitrogen), supplemented with 10% fetal bovine serum (FBS, from PAA Laboratories; for hSCs, we used FBS from Sigma-Aldrich) and penicillin/streptomycin (Gibco®/Life Technologies). The hSC medium was further enriched with 0.2% glucose (Gibco) and 2 μM forskolin (Sigma-Aldrich). Cells were incubated in a humidified atmosphere (5% CO2, 37°C) for 48 h before sample preparation.
Sample Preparation
Cells obtained from the culture of different sources were harvested and washed twice with phosphate-buffered saline (PBS, pH 7.4), centrifuged at 250 g for 10 min, and stored at −80°C for further use. In addition, human lymphocytes were obtained from isolated fresh blood samples with Lymphoprep™ (Axis-Shield) according to the manufacturer's procedure. The cell pellets were dissolved in ice-cold lysis reagent (1×tris-buffered saline pH 7.4, 1% triton X-100, 1 mM ethylenediaminetetraacetic acid, 1 mM ethylene glycol tetraacetic acid, 0.05% sodium dodecyl sulfate, 0.1% sodium deoxycholate, and 2% nonidet-P40) with freshly added protease inhibitors (1 mM phenylmethylsulfonyl fluoride [from Sigma], 5 mM sodium fluoride [Merck Chemicals], 1 mM sodium orthovanadate, and protease inhibitor cocktail [#PI8340 from Sigma Aldrich]) and incubated on ice for about 25 min before centrifugation at 10,000 g for 12 min at 4°C. Then, the supernatant was analyzed by the BCA Protein Assay Kit (Thermo Scientific Pierce) for protein concentration.
Buccal Cell Preparation
Buccal cells were harvested using MasterAmp Buccal Swab Brushes (Epicentre Biotechnology) by firmly pressing a brush against the inner cheek and swabbed while twirling for 60 s on each cheek. Both brushes were then immersed in the same tube containing 500 μL of ice-cold PBS and twirled gently to dislodge the cells. Samples were then microcentrifuged at 10,000 g for 10 min at 4°C, and the cell pellet was lysed in the ice-cold lysis reagent as described in the sample preparation section. The protein concentration was calculated using the BCA protein assay as described above.
SMN ECLIA Protocol
The capture antibody was coated in the 96-well Multi-Array High Bind plates (MSD) by overnight incubation at 4°C. Next day, the plates were blocked at room temperature with 125 μL MSD Blocker A. The plates were washed thrice with PBS-Tween (0.05%) before adding the samples (25 μL per well diluted in 1% MSD Blocker or cell lysis buffer). The sample preparations were incubated at room temperature for another 90 min or at 4°C for 16 h in case of buccal cell lysates. Each well was incubated with the polyclonal rabbit anti-SMN antibody (0.5 μg/mL in 1% MSD Blocker A) for 75 min followed by one incubation of 0.75 μg/mL of MSD Sulfo-TAG™ goat anti-rabbit detection antibody in 1% MSD Blocker A. One hundred fifty microliter Tris-based Read Buffer T (1×) with surfactant (MSD) was added to the plate and signals from ECL were captured by the Sector Imager 2400 reader (MSD) and recorded as signal counts (Table 1).
Spinal Muscular Atrophy Electrochemiluminescence Immunoassay Protocol Table
PBS, phosphate-buffered saline; RT, room temperature; MSD, Meso Scale Discovery.
SMN Copy Number
Quantification of SMN1 and SMN2 copy number in four healthy donors was performed by Multiplex Ligation-dependent Probe Amplification (MLPA) using P021-A2 and P060-B2 SALSA MLPA® probemix (MRC-Holland) according to the manufacturer's instructions. Genomic DNA was extracted from blood lymphocytes by standard salt precipitation, 24 and a total of 150 ng was used in each reaction. MLPA was run in duplicate for each donor and four different DNA standards.
Statistical Analysis
Statistical analysis of the measurement data was performed with GraphPad Prism software. Results are presented as mean±standard deviation and mean±standard error of the means. Statistical significance of differences was examined by using the Student's t-test. Significant differences are marked in the figures with *(P<0.05), **(P<0.01), and ***(P<0.001).
Results
Assay Specificity, Reproducibility, and Sensitivity
The ECLIA is based on 96-microwell plates with carbon electrodes built into the bottom for ultrasensitive detection. 25 A standard curve with recombinant human SMN1 is shown in Figure 1. Recombinant human SMN1 can be measured over a wide range (0.76–5,000 pg per well with R 2 =0.9983) with high accuracy, which equals a dynamic range of 30–200,000 pg/mL.
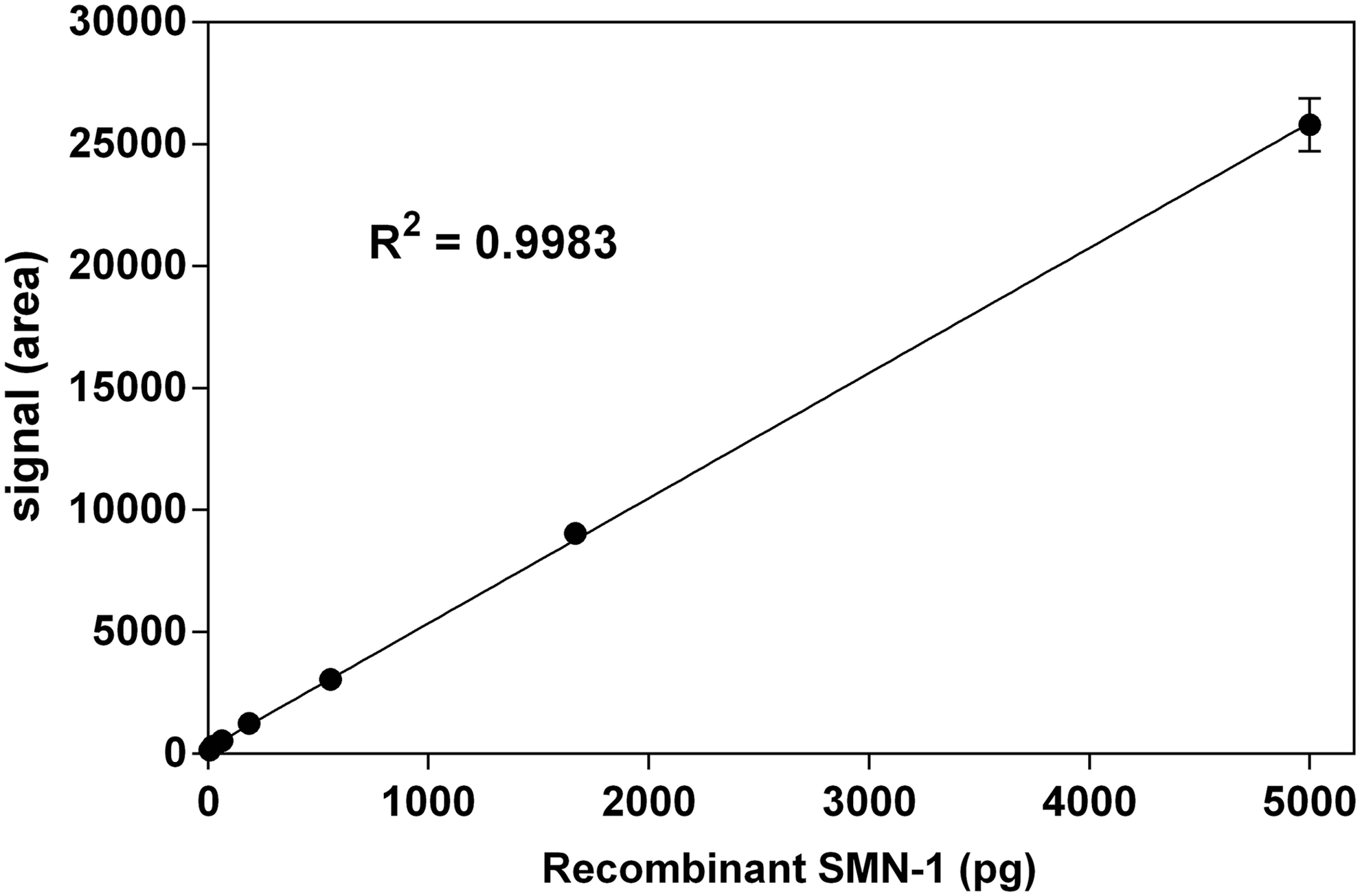
Standard curve of recombinant human SMN1. Error bars show standard error at each dilution (n=3). Linear regression in a range from 0.76 to 5,000 pg per well was generated using GraphPad Prism software. SMN, survival motor neuron.
The recovery rate was estimated by spiking a definite amount of different cell lysates with low and high amounts of recombinant human SMN1. The recovery rate of spiked recombinant SMN1 with human SMA fibroblasts was 80% (high amounts) and 90% (low amounts). Similarly, we estimated a recovery rate for F11 cells of 79% (high amounts) and 93% (low amounts) (Data not shown).
Validation of the Assay
The SMN ECLIA is able to quantify SMN protein levels in a variety of cell lines and tissues of human, mouse, and rat origin. The working range was found to vary among different sample types (Table 2), which on one hand is most likely proportional to the concentrations of SMN per mg protein in different tissues and on the other hand also reflects differences in the reactivity of the assay toward the SMN protein of different species. The measurement of different samples on three ensuing days yields an intraday precision of coefficient of variation (CV) ≤2.73% and an interday precision of CV ≤7.09% (Table 3).
Working Ranges of Different Sample Materials
Suggested working ranges for various human, mouse, and rat sample materials.
SMA, spinal muscular atrophy; DRG, dorsal root ganglion.
Intra-Assay and Interassay Precision of Survival Motor Neuron Electrochemiluminescence Immunoassay
Intra-assay precision (n=3 wells per day) and interassay precision (n=3 separate days).
SD, standard deviation; SEM, standard error mean; CV, coefficient of variation.
Quantification of SMN Protein Levels in Human Buccal Cells
To investigate whether our ECLIA would be sensitive enough to quantify the SMN protein in human buccal cells, we isolated buccal cells from one male healthy adult. The protocol adapted to this was as described in the Materials and Methods section because higher accuracy was achieved by overnight incubation of the samples.
The linearity over a wide range of buccal cell lysates from a healthy donor prompted us to investigate SMN protein levels in a small cohort of healthy donors (n=4, Fig. 2). For this, we collected buccal cells from an unmatched group of controls thrice within a 48-h interval (Table 4).
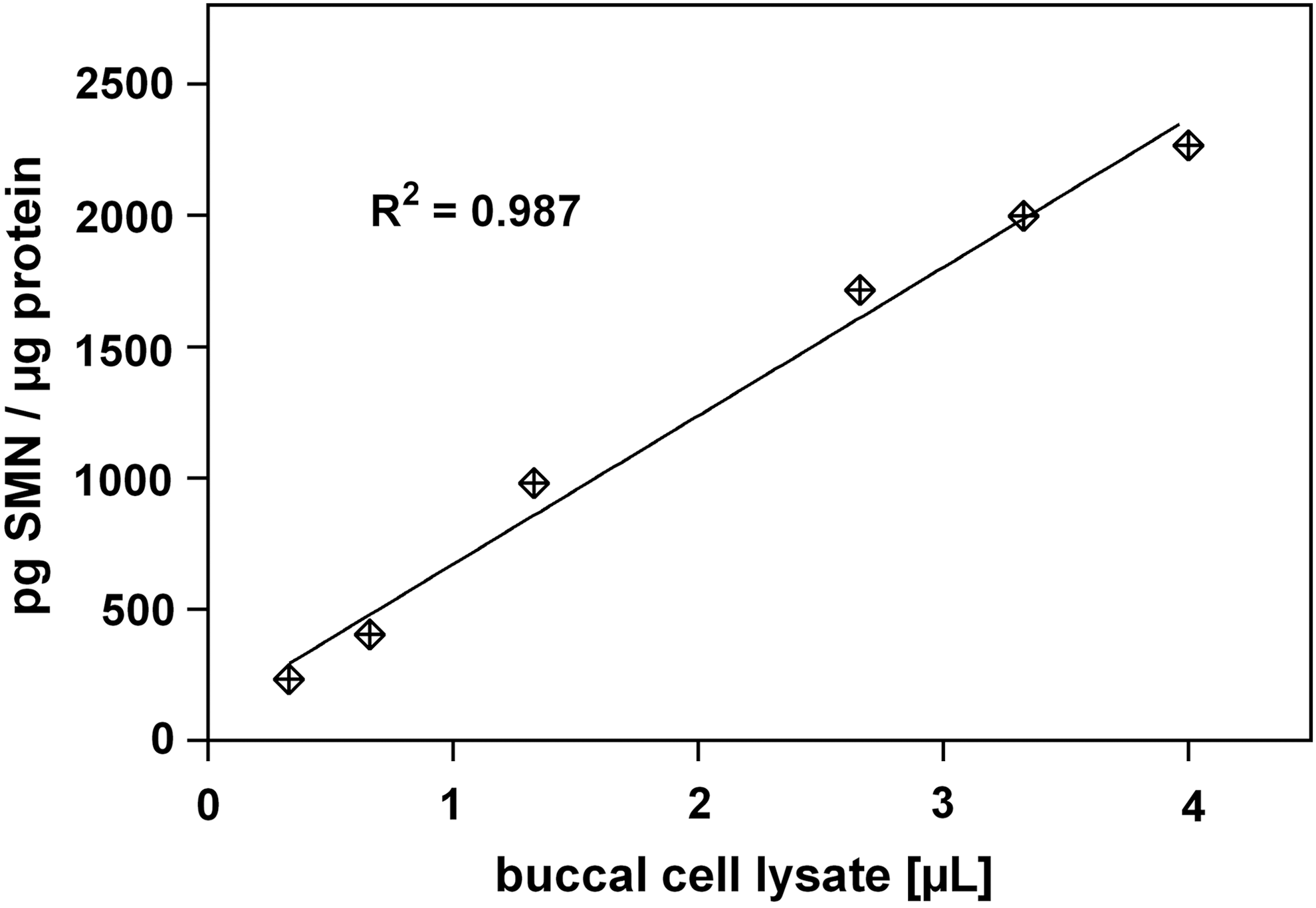
Linearity of buccal cell lysate. The assay remains sensitive and in the linear range (R 2 =0.98) from 0.3 to 4 μL buccal cell lysate per well.
Demographic Values of Healthy Donors, One Spinal Muscular Atrophy Carrier, and One Spinal Muscular Atrophy Patient
Individual values of age (years) at the study, sex, and copy numbers of SMN1 and SMN2 are shown. No genotyping was performed on SMA carriers and SMA patients in this study, and the fathers self-identified themselves as SMA carriers and patients based on prior testing and diagnoses.
SMA, spinal muscular atrophy.
Although we found a high variability in SMN protein levels (Fig. 3) in this group of healthy donors, we investigated the possibility to measure SMN protein levels of buccal cell lysates in an SMA carrier and patient. We collected buccal cells from one SMA carrier (male, 36 years old) and one SMA Type III patient (male, 6 years old) and measured SMN protein levels from buccal cell lysates with the ECLIA-based assay.
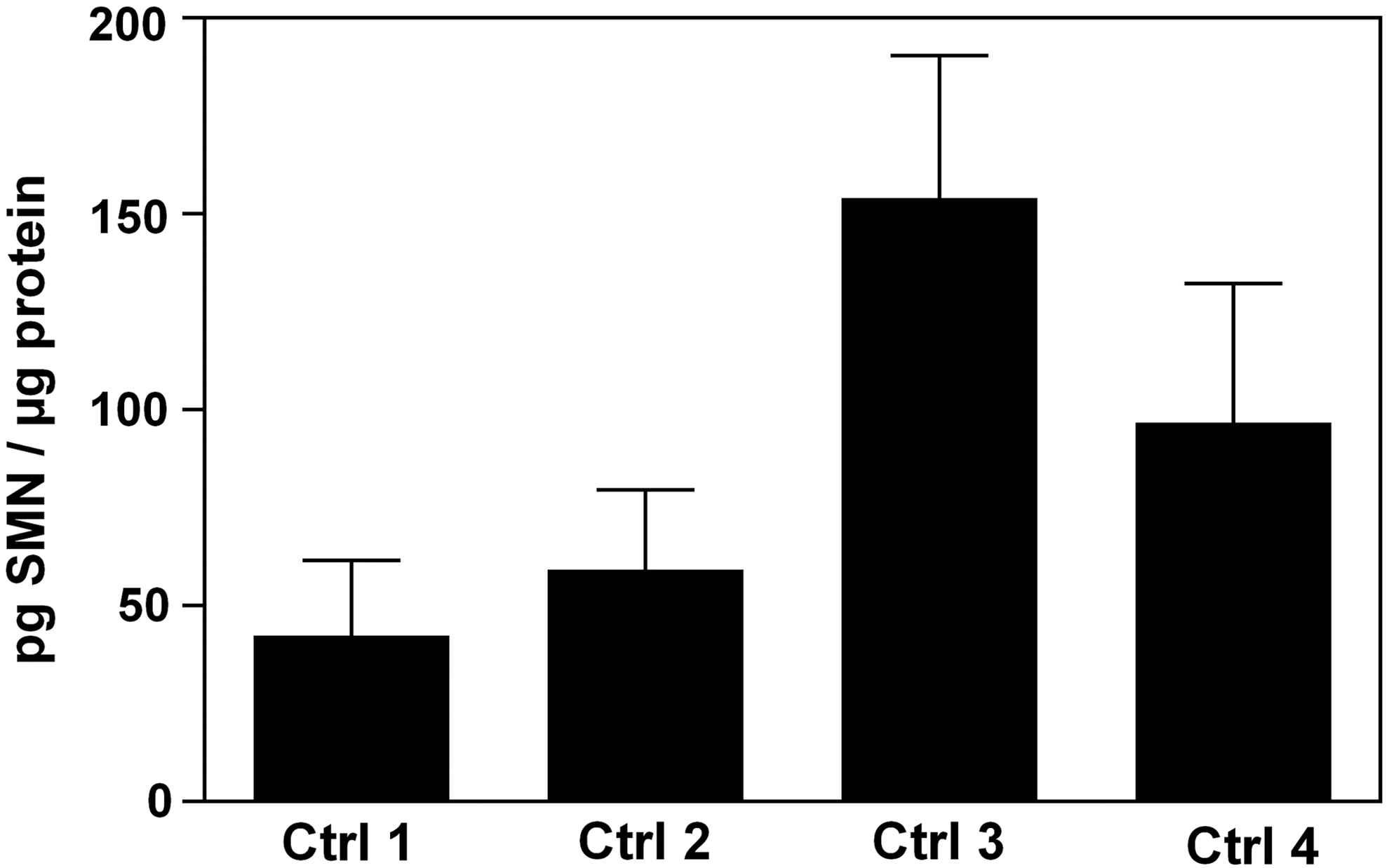
SMN protein levels in human buccal cells from healthy adults. Buccal cells were collected preprandial thrice from four healthy adults (Ctrl 1–4) on five consecutive days at every 48 h. SMN protein levels were measured in buccal cell extracts and normalized to protein levels. The data presented are mean±SD of triplicate wells (n=3). Ctrl, control; SD, standard deviation.
As shown in Figure 4, SMN was readily detected in buccal cell lysates from the SMA carrier and a Type III SMA patient. The SMN protein level in the SMA carrier was slightly reduced compared to a sex- and age-matched healthy donor (Ctrl 2). The amount of SMN protein in the SMA patient was highly increased in relation to both, the control and the carrier, but could not be compared to an age-matched healthy control in this preliminary assessment.

SMN protein levels in human buccal cells from SMA carrier and SMA patient. SMN protein levels in human buccal cells derived from SMA carrier and SMA patient compared to healthy control (Ctrl 2). The data presented are mean±SD of triplicate wells (n=3). SMA, spinal muscular atrophy.
Discussion
We developed and validated an ECLIA for the quantitative measurement of SMN protein levels in human, mouse, and rat samples. Human SMN, which is ubiquitously expressed, has a molecular weight of 38 kDa and localizes to both the cytoplasm and the nucleus. It is part of a multiprotein complex that contains multiple SMN molecules and a number of stoichiometrically interacting proteins, Gemins 2–8. 12 As with other SMN quantification methods, lysis reagents used in the ECLIA protocol have to be chosen carefully to ensure extraction and dissociation of both cytoplasmic and nuclear SMN. 16 In the protocol described here, a commercially available lysis buffer known to work well in the ECLIA setting (MSD) is used, slightly modified for SMN extraction. Compared to other extraction buffers used in preliminary trials, it yielded higher concentrations of SMN in relation to whole protein (data not shown), suggesting a more efficient extraction of nuclear SMN.
Both antibodies adapted for our ECLIA protocol are used in a well-established ELISA that has recently been used for measurement of blood SMN levels in a large clinical SMA study. 13,19 The primary antibody (2B1) is a mouse monoclonal antibody that binds to the N-terminus of human SMN at amino acid positions 14–20 19,26 ; the epitope for the secondary antibody (11708-1-AP), which is a rabbit polyclonal antibody, is located between amino acids 197 and 204 of human SMN. 19 As both epitopes are expected to be present in full-length SMN as well as in SMNΔ7, it is conceivable that both proteins are detected by our ECLIA, thus potentially interfering with the measurement of slight shifts in the ratio of SMN/SMNΔ7. This should be kept in mind when using the ECLIA in preclinical screenings for drugs that increase the rate of exon7-inclusion in SMN2-derived transcripts. Within the context of clinical studies, however, SMNΔ7 is unlikely to substantially distort the quantification of SMN in tissue samples, as the truncated protein contains a potent degradation signal that causes it to be rapidly degraded. 10
With regard to the assessment of SMN levels in murine samples using ELISA, concerns have been raised by Kobayashi et al. as to the differences in sensitivity. The authors report on a marked difference in reactivity of the assay to the SMN protein in humans and mice, respectively, with recombinant mouse SMN being detected at only about 10% the levels of recombinant human SMN. 19 It is reasonable to assume that this holds true for the ECLIA as well, as the two assays are based on the exact same primary and secondary antibody.
The ECLIA described here has a broad dynamic range (0.76–5,000 pg/well with R 2 =0.9983) and a detection limit of 30 pg/mL for the human SMN protein. It is therefore more sensitive than sandwich format ELISAs, whose reported detection limits range from 25 ng/mL 18 to 50 pg/mL. 19 As a result, SMN levels of various origin can be reliably assessed even at great dilutions, which means the ECLIA is less prone to matrix effects than other protein quantification methods and thus less subject to intersample variation. In line with its enhanced sensitivity, the ECLIA readily detected SMN in buccal swabs of a Type III SMA patient, an adult SMA carrier, and a healthy adult donor. To our knowledge, this is the first report of SMN protein quantification in human buccal cells. As they are particularly easy to access, it is tempting to speculate that buccal cells might constitute a noninvasive and practicable way to monitor SMN levels in future trials of SMN-upregulating therapeutics. However, earlier studies using ELISA to assess SMN levels in different human tissue types have pointed at various pitfalls with regard to that approach. First, quantitative evaluation of murine tissues suggest that Smn expression varies widely across tissues of the same genotype, with Smn levels being found to be lowest in the primary tissues associated with SMA, that is, nerve, spinal cord, and muscle. 19 It has been pointed out that SMN levels measured in PBMCs do not necessarily fully reflect SMN expression in more disease-relevant cells like motor neurons or muscle cells. 13 The same holds true for buccal cells and any other peripheral tissues.
Second, age is an important factor to consider when assessing SMN levels in clinical specimens, as there is evidence of a general decline of SMN levels across tissue types and species, 16 and several studies using ECLIA to measure SMN levels in PBMCs of SMA patients, carriers, and controls have stressed the necessity to control for age when performing any statistical analysis. 13,19
Third, even during a short period of time, SMN levels measured in PBMCs fluctuate substantially within individuals; when assessed at five time points over the course of 1 month, blood SMN levels were shown to vary from 2 to 10 times in six different healthy adults. 19 Whether SMN levels measured in buccal cells using ECLIA are subject to such an extent of intraindividual fluctuation has to be evaluated by repeated within-subject testing, preferably normalizing SMN signals by cell counts instead of total protein.
Finally, the finding that there is no clear correspondence between SMN protein levels in blood cells and disease severity raises fundamental concerns with regard to using peripheral SMN protein levels as an indicator of response to SMA treatment; while a correlation of protein quantity with phenotype could be established for severely affected Type I and Type II patients, no substantial decrease in the SMN protein is detectable in Type III patients. 27,28 In line with this observation, a recent multicenter clinical trial using SMN ELISA to assess SMN levels in PBMCs of adult Type III SMA patients did not show any correlation between protein quantity and motor performance, leading the authors to suggest that SMN protein levels in blood cells are not suitable as biomarkers in clinical trials on SMA. 29 In light of the data acquired from Type III SMA blood cells, our preliminary finding of a high SMN protein level in buccal cells of one Type III SMA patient is not that surprising, given the age difference between the patient (6 years) and the healthy control (31 years). It is doubtful that SMN levels in buccal cells will provide a more reliable indicator of disease severity in a mildly affected SMA patient than PBMCs, thus compromising the use as a biomarker in SMN upregulation trials.
Taken together, the use of ECLIA to monitor SMN levels in buccal cells as a biomarker in clinical SMA drug trials is far from being straightforward. However, our data suggest that it can be effectively used to measure SMN protein levels as well as slight changes thereof in cell models of different species with high accuracy and specificity over a wide range. In terms of sensitivity, the ECLIA outperforms previous quantification methods. Furthermore, due to its relatively easy handling in a 96-well plate format, the assay may represent a novel tool of great value for high-throughput drug discovery screenings.
Footnotes
Acknowledgments
We gratefully thank Dr. Mark Dürr from Meso Scale Discovery (MSD, Germany) for his help with the ECLIA optimization and Dr. Oliver Brandau for collection of blood samples. Furthermore, we thank all donors for buccal cell samples.
Author Contributions
All the authors of this article fulfilled the criteria of authorship. H.S. performed the experiments. H.S., J.E., B.M.G., and F.L. designed the study and wrote the article.
Disclosure Statement
No competing financial interests exist.