
Abstract
Voltage-gated calcium channels represent important drug targets. The implementation of higher throughput electrophysiology assays is necessary to characterize the interaction of test compounds with several conformational states of the channel, but has presented significant challenges. We report on the development of a high-throughput, automated electrophysiology assay for Cav2.2 on the IonWorks Barracuda™ platform. The assay provides an assessment of the potency of the test compound on the resting/closed and inactivated states of the channel in the same assay run. Inclusion of the heavy metal chelator 2,3-bis(sulfanyl)propane-1-sulfonate in the assay solutions improved the data quality by reversing a loss of current seen in wells directly above the ground electrodes. We hypothesize that the loss of current is caused by block of Cav2.2 currents by silver ions originating from the electrodes.
Introduction
Intracellular calcium concentration is a key regulator of muscle contraction, hormone and neurotransmitter secretion, and gene transcription. Voltage-gated calcium channels link membrane potential to intracellular calcium concentration and represent important drug targets. This is illustrated by the more than 60 calcium channel blockers in clinical development in the 1980s. 1 At the time, clinical development focused on the subfamily of Cav1 (L-type) channels and resulted in the approval of numerous important drugs used to treat hypertension and angina. Despite the expression and important physiological role of Cav1 channels in skeletal, cardiac, and smooth muscle, in endocrine cells, and in the peripheral and central nervous system, L-type calcium channel blockers are generally well tolerated. 2 This functional selectivity for inhibition of Cav1 channels in vascular smooth muscle beds and resulting safety and tolerability profile is attributed in large part to the state-dependent mechanism of these drugs, interacting preferentially with open and inactivated states of the channel. 3,4
Similar to early Cav1 blockers, blockers of voltage-gated sodium channels were found empirically and were shown in in vivo studies to have benefit as local anesthetics, anticonvulsants, and antiarrhythmics. As with Cav1 blockers, therapeutically beneficial sodium channel blockers, such as lidocaine, inhibit sodium channel function by binding selectively to channels in inactivated conformational states. 5
More recently, attention has been focused on the Cav2 subfamily of voltage-gated calcium channels and on their potential as analgesic targets. Much of the interest comes from the clinical efficacy of the Cav2.2 (N-type) selective peptide blocker ziconotide, marketed as Prialt™. 6 In addition to the intrathecal route of administration, the use of ziconotide is limited by the common occurrence of adverse events, including dizziness, nausea, and confusion. By analogy to blockers of Cav1 and voltage-gated sodium channels, it is hypothesized that a state-dependent mechanism of inhibition may lead to better tolerability of Cav2.2 blockers. 7
Determining the potency of drug interactions with specific conformational states of voltage-gated channels requires excellent voltage control combined with high-temporal solution. These are the hallmark of electrophysiological measurements, but typically either absent or inadequate in traditional higher throughput flux or binding assays.
Although manual electrophysiology assays can determine state-dependent parameters with high fidelity, their low throughput permits profiling of only a limited number of compounds, relegating the use of these assays mostly to target validation and late stage preclinical assessment. Recently, several automated electrophysiology platforms have emerged. 8 Medium throughput platforms, such as PatchXpress™ (Molecular Devices, Sunnyvale, CA), Q-Patch™ (Biolin Scientific, Stockholm, Sweden), and PatchLiner™ (Nanion Technologies, Munich, Germany), are characterized by high-resistance seals, approaching those typical of manual electrophysiology, and high-fidelity signal amplification and conditioning, at the expense of higher operating cost and lower throughput. The IonWorks™ instruments (IonWorks HT and IonWorks Quattro; Molecular Devices) represent the first truly high-throughput platforms reaching the market. Depending on the specifics of the assay and cell line, these instruments may achieve near 100% success rate, but assays are limited by lower fidelity of voltage control and by the inability to clamp membrane potential during compound addition. IonWorks Barracuda™ (IWB; Molecular Devices) was launched in 2010 with the intent of maintaining all of the IonWorks Quattro capabilities while eliminating most of its deficiencies. Seal resistances on IWB remain in the range of 50–200 MΩ, but voltage clamp is continuous in 384 wells in parallel. This makes IWB suitable for recording from most classes of ion channels with high-success rate, and successful profiling of voltage-gated 9,10 and rapidly desensitizing ligand-gated channels 11 has been reported.
Recent attempts at recording Cav2 currents on IWB were successful, but lacked sufficient stability to continuously clamp membrane potential near the half-inactivated voltage. 10 To assess the interaction of a test compound with the inactivated channel, it is desirable to maintain a defined fraction of channels in the inactivated state continuously to avoid effects of channel kinetic properties and compound binding rates on the measured blocking potency. 5
We have designed a protocol that enabled us to measure block of calcium current at resting and partially inactivating membrane voltages within the same recording. A problem we encountered during assay development was the nonuniformity of calcium currents across the patchplate. Here we show evidence that nonuniformity resulted from silver ions partially blocking calcium currents in wells above the ground electrodes. Uniformity can be restored by inclusion of the heavy metal chelator 2,3-bis(sulfanyl)propane-1-sulfonate (DMPS).
Materials and Methods
Materials
Unless otherwise specified, all cell culture reagents were obtained from ThermoFischer Scientific (Waltham, MA), and all other reagents were obtained from Sigma-Aldrich (St. Louis, MO).
Cell Culture
An HEK293 cell line stably expressing rat Cav2.2 α1B (NM_001195199.1), β3 (NM_012828.2), and α2δ (NM_012919.2) was generated by transfecting cells simultaneously with pcDNA3.1-based expression constructs for all three subunits. Resistance markers used were G418, zeocin, and hygromycin. Stable expression was confirmed by western blotting with the following antibodies: Alomone Cat# ACC-002 for α1B, Abcam Cat# ab16717 for β3, and ProScience Cat# 28–220 for α2δ. For use in a fluorometric imaging plate reader-based calcium influx assay, which is not the subject of this article, the resulting cell line was further transfected with a pJBO2-based expression construct encoding the rat Kir2.1 subunit (NM_017296.1) and puromycin selection marker. Cells were cultured at 37°C in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 1% nonessential amino acids, G418 (200 μg/mL), hygromycin (100 μg/mL), zeocin (50 μg/mL), and puromycin (0.1 μg/mL).
For IWB experiments, cells were cultured in T-150 flasks (Corning, Corning, NY) to 80%–90% confluence. Cells were rinsed twice with D-PBS without calcium and magnesium and dissociated by incubating in 3 mL of TrypLE™ (ThermoFischer Scientific) for 5 min. Cells were then resuspended in 10 mL of media, gently triturated, and centrifuged for 5 min at 1,000 rpm. The supernatant was aspirated and the cells were resuspended in 5 mL Hanks-balanced salt solution (HBSS)-based external solution (see “Solutions” section).
Solutions
During IWB recordings, cells were supplied and seals were established in an external solution consisting of HBSS supplemented with 20 mM HEPES and pH adjusted to 7.4 using NaOH. Once stable seals were established, the HBSS was replaced by the recording solution containing the following (in mM): 135 NaCl, 10 BaCl2, 1.2 MgCl2, 0.5 EGTA, and 20 HEPES; pH was adjusted to 7.4 using NaOH. The internal solution used contained the following (in mM): 90 K-Gluconate, 40 KCl, 3.2 MgCl2, 3.2 EGTA, and 5 HEPES; pH was adjusted to 7.2 using KOH. The membrane perforating agent amphotericin B was prepared as a 28 mg/mL stock solution in dimethyl sulfoxide (DMSO) on the day of the experiment and added to the internal solution at a concentration of 0.1 mg/mL.
Electrophysiological Recordings
The IWB instrument was primed with intracellular and extracellular solutions and 9 μL of cells (5M/mL) was added to each well. Recordings were performed in population patch clamp (PPC) mode, and the whole cell recording configuration was established by 8 min incubation with amphotericin B. Sampling frequency was 5 kHz. The command voltage protocol consisted of three linked (concatenated) protocols: cells were initially voltage clamped at −100 mV until current, determined using 20 ms test pulses to 10 mV, had stabilized. Cells were then depolarized to a holding potential corresponding to ∼60% channel availability. After a baseline recording, test compound was added and block was monitored with 20 ms test pulses to 10 mV. After 10 min, cells were repolarized to −100 mV. For a detailed assay protocol, see Table 1.
Protocol for Cav2.2 Electrophysiology Assay on IWB
HBSS, Hanks-balanced salt solution; IWB, IonWorks Barracuda.
Data Analysis
Data acquisition, leak subtraction, and initial analysis of peak currents during each test pulse were performed using IWB software (version 2.0.2; Molecular Devices Corporation, Union City, CA). Acceptance criteria are shown in Table 2. Compounds were tested at 10 concentrations in quadruplicate, all currents were normalized to DMSO controls on the same plate, and mean and standard error of the normalized currents were fit to the Hill equation using GraphPad Prism 6.02 software (GraphPad Software, San Diego, CA). All error bars represent the standard error of the mean. Student's t-test was used to determine statistical significance.
Acceptance Criteria for IWB Cav2.2 Assay
Chemicals
Compound serial dilutions were prepared in DMSO and diluted into external recording solution before the experiment. Final DMSO concentration was 0.3%.
Results
Cav2.2 Currents Recorded on IWB
Barium currents conducted by recombinant Cav2.2 channels were recorded successfully on the IWB high-throughput electrophysiology platform. Barium was chosen as the charge carrier to minimize calcium-dependent inactivation 12 and to block currents conducted by the Kir2.1 channels coexpressed in our stable cell line. 13 Figure 1A shows a family of current traces evoked by depolarizing pulses from a holding potential of −100 mV. Similar to what is seen in manual electrophysiology recordings, the current activated rapidly and inactivated slightly during the 100 ms test pulse. Normalized peak currents, recorded 6 and 28 min after obtaining the perforated patch recording configuration, were plotted against test pulse voltage and are shown in Figure 1C. The current–voltage relationship shows a relatively steep voltage dependence of current activation, with a slope of 2.4 mV compared to 3.8±0.5 mV (n=11) in manual electrophysiology experiments on the same cell line. The relatively steep voltage dependence may be an indication of imperfect voltage control on IWB. However, stability over time was excellent. Steady-state inactivation was examined with a series of 1 s conditioning pulses, followed by a test pulse to 0 mV (Fig. 1B). Fitting the Boltzmann equation to the data indicated a half-inactivation voltage of −20.3 mV with a slope of 7.3 after 6 min (Fig. 1D). As seen for voltage dependence of activation, steady-state inactivation was stable over at least 28 min.

Rat Cav2.2 currents recorded in population patch clamp mode on IonWorks Barracuda.
State-Dependent Pharmacology
To assess the effect of test compounds on channels in resting and in inactivated states, we devised a concatenated protocol using the multiple protocol mode of IWB (Fig. 2A, bottom panel). The holding potential was initially set to a voltage (−100 mV) where most channels are in closed (resting) states available for activation, and current amplitudes were monitored with test pulses to 10 mV. Next, the holding potential was adjusted to correspond to ∼40% channel inactivation. From day to day, we saw some variability in this value, and holding potentials between −38 and −45 mV were used. Current amplitudes were again monitored with test pulses to 10 mV at 2 min intervals, and test compounds were added after two baseline pulses. After a 10 min compound equilibration, the holding potential was returned to the starting voltage. The example in Figure 2A shows the normalized peak currents measured during each test pulse for a control vehicle addition (middle panel), and representative current traces were recorded at the indicated times during the protocol (top panel). Only experiments with channel inactivation ranging from 35% to 60% were used in the analysis.
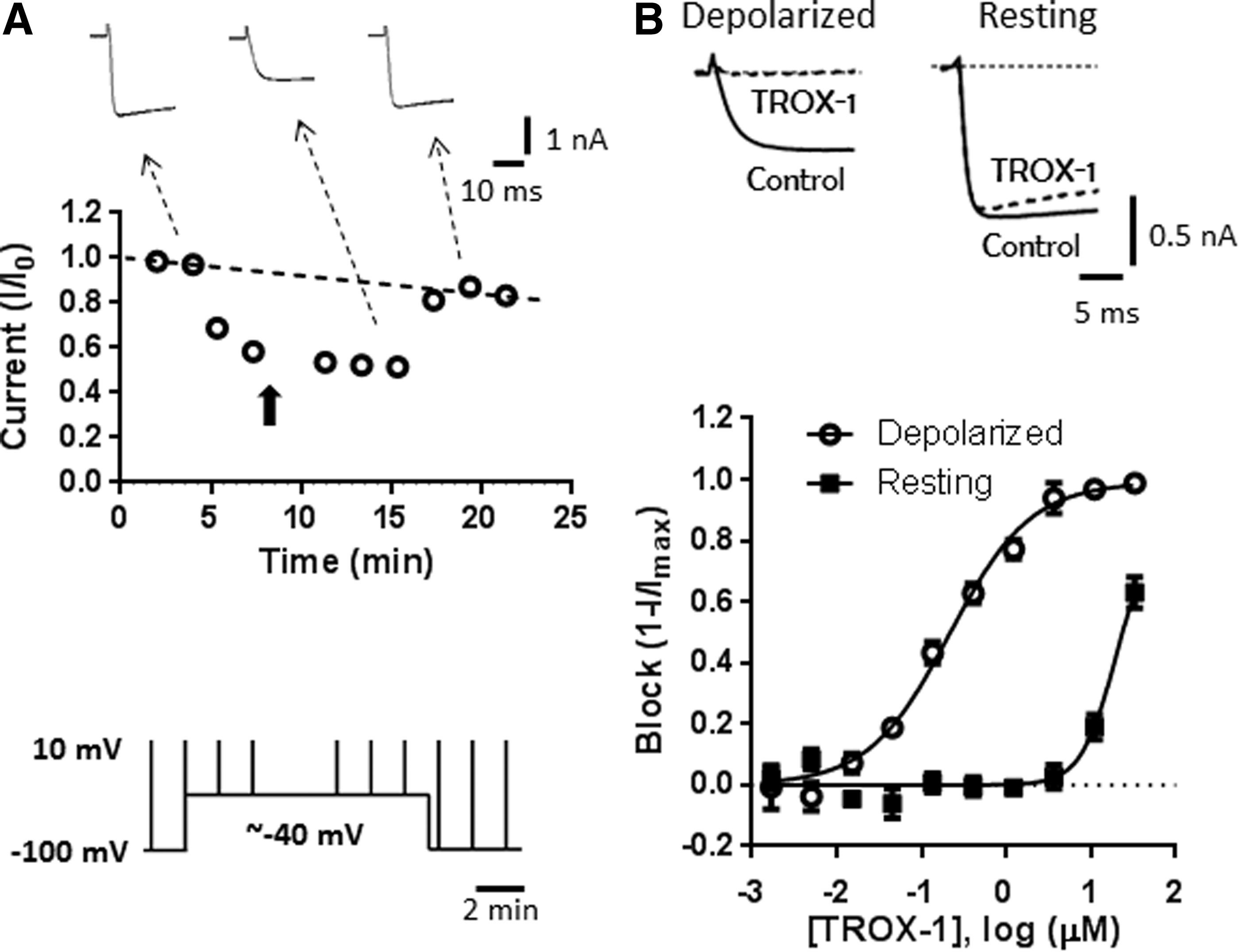
Quantification of voltage-dependent Cav2.2 block on IonWorks Barracuda.
An advantage of this protocol is the ability to determine compound effect on channels in two different conformations (resting and inactivated) within the same recording. Effects on channels in the inactivated conformation are evaluated by determining a block relative to DMSO controls during the last test pulse from the depolarized holding potential. Block of channels in the resting state is defined as a block measured ∼3 min after returning to −100 mV. For compounds that recover slowly from inactivated state inhibition, this protocol may overestimate the resting state potency. Differential effects at the two different holding potentials are illustrated in Figure 2B for the known state-dependent Cav2 blocker TROX-1. 14 At 1 μM, TROX-1 had little effect on currents evoked from −100 mV, but almost completely blocked currents evoked from the more depolarized holding potential of −40 mV. Fitting the data with the Hill equation yielded IC50 values of 0.19±0.06 and 25.3±5.9 μM (n=20) at depolarized and resting holding potentials, respectively.
Spatial Uniformity of Cav2.2 Current
Although initial experiments looking at activation and steady-state inactivation curves indicated good stability of the recordings over time (Fig. 1), we wanted to assess the stability of currents at partially inactivating voltages in more detail. To this end, we calculated the ratio of the currents evoked by the second and last test pulse given from the more depolarized holding potential (see protocol in Fig. 2A). These ratios are averaged across five experiments for each well in the patchplate. Figure 3A shows a heat map of the ratios across patchplate positions. The heat map clearly shows four areas of the patchplate where currents were substantially reduced during the last test pulse after buffer addition compared to the control currents before buffer addition. The areas of reduced currents coincide with the location of the four ground electrodes situated underneath the patchplate in the reservoir that holds the internal solution. Figure 3B illustrates the effect this nonuniformity of currents has on a typical 10-point concentration–response curve. In this experiment, TROX-1 was titrated in rows across the patchplate, with two titrations per row starting in columns 2 and 13. Concentration–response curves are constructed from wells at the periphery of the patchplate, where there was no loss of current with time (in black), and from rows that included the areas above the ground electrodes (in red). Although IC50 values only differed by twofold (99 nM vs. 49 nM), the effect of the nonuniformity on the shape of the curve is clearly visible.
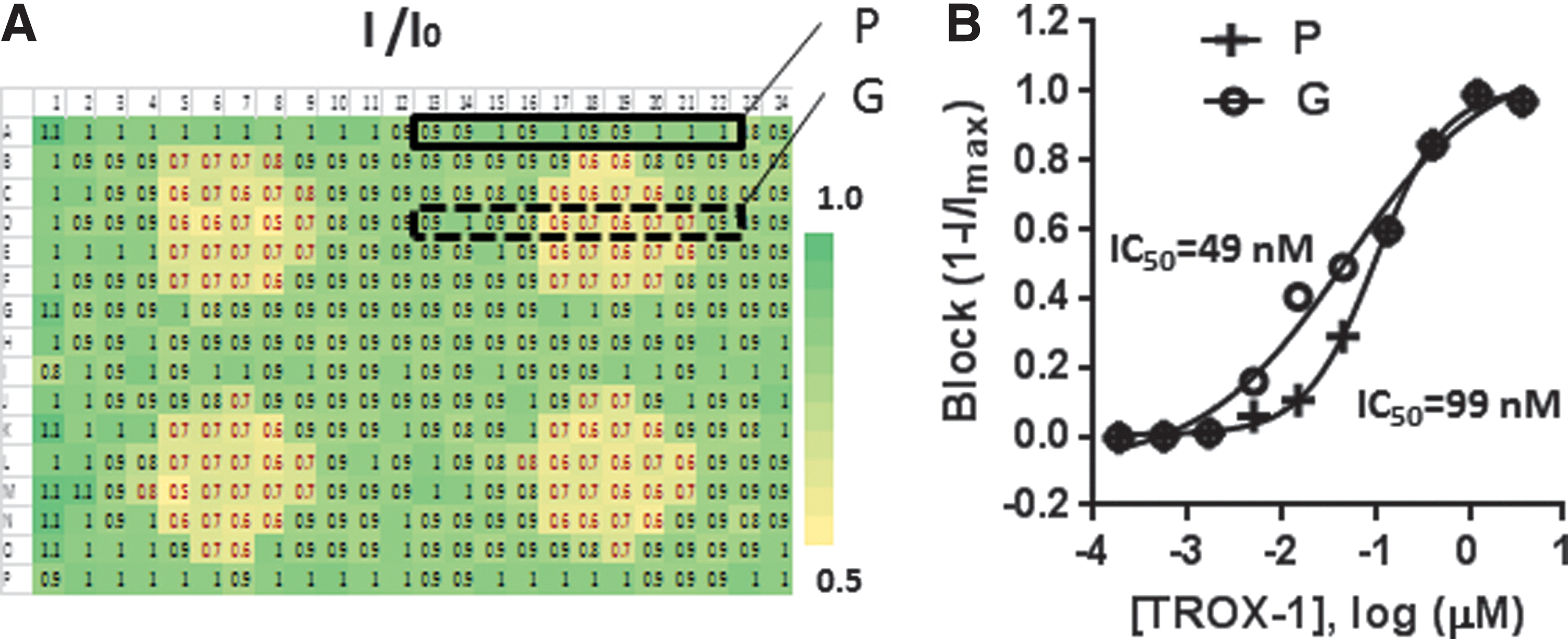
Nonuniformity of currents on IonWorks Barracuda.
Block of Current by Silver
As is typical for voltage clamp experiments on any platform, the IWB uses Ag/AgCl electrodes. Since the currents that are conducted during a typical experiment especially using the PPC mode 15 are large, the size of the compressed AgCl pellet that is a component of the common ground electrode is substantial. Inhibition of Cav3.1 (T-type) calcium currents by Ag ions has been reported, 16 and we hypothesized that the nonuniformity seen in the IWB recordings may be caused by Ag ions coming off the ground electrode. Therefore, we evaluated the effect of AgCl on Cav2.2 currents on IWB (Fig. 4A). AgCl was titrated from a top concentration of 5 μM, where it produced complete block of Cav2.2 in the absence of changes in the pH of the external solution. AgCl was equipotent at partially inactivating (IC50=0.42 μM) and at resting (IC50=0.33 μM) holding potentials. Next, we explored AgNO3, a silver salt with better aqueous solubility (Fig. 4B). High concentrations of AgNO3 (10–30 μM) were often associated with a decrease in seal resistance; however, no change in seal resistance was observed in 1.2 μM AgNO3, a concentration that produced ∼80% block of Cav2.2 currents. As expected, the potency of AgNO3 for blocking Cav2.2 currents was similar to that of AgCl with IC50 values of 0.49 and 0.44 μM at partially inactivated and resting holding potentials, respectively. In contrast, NaNO3 had no effect on Cav2.2 currents at concentrations up to 30 μM (data not shown).
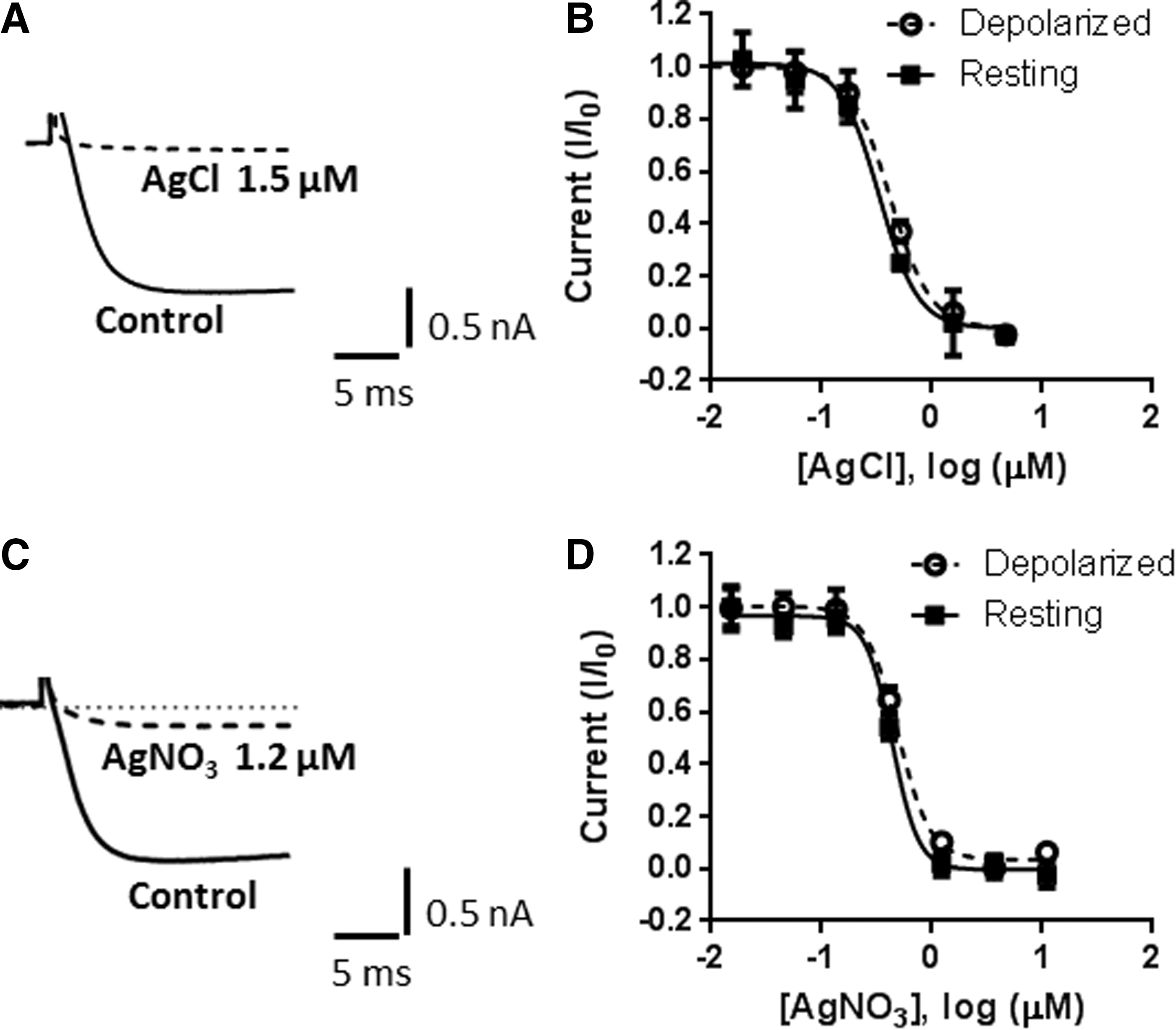
Block of Cav2.2 currents by silver. Representative traces
Block of Cav2.2 by AgNO3 is Reversed by the Chelator DMPS
DMPS is a heavy metal chelator approved for clinical use in cases of heavy metal poisoning. 17,18 It has high affinity for silver and relatively low affinity for calcium and magnesium. At high concentrations, DMPS applied in the extracellular solution blocked Cav2.2 current (IC50=5.4 mM; Fig. 5A, B). However, addition of 1 mM (extracellular and intracellular) DMPS completely prevented block of Cav2.2 by AgNO3, without affecting Cav2.2 currents in the absence of AgNO3 (Fig. 5C, D). Similar results were obtained with 0.1 mM DMPS (data not shown).
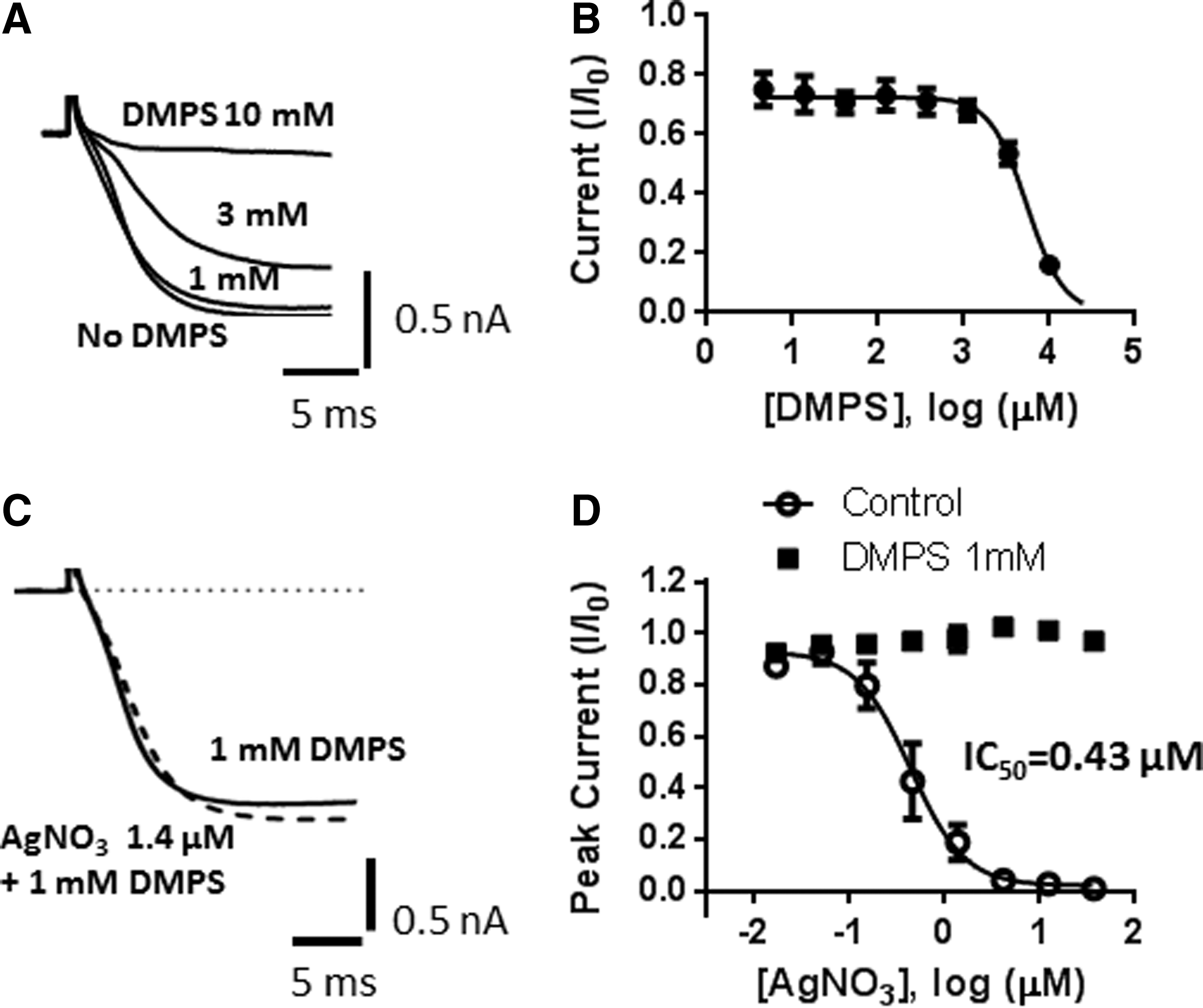
Block of Cav2.2 by AgNO3 is reversed in the presence of DMPS.
DMPS Prevents the Loss of Cav2.2 Current Over Electrodes and Improves the Uniformity
We next tested the effect of DMPS on the stability of Cav2.2 currents in our IWB concatenated protocol. DMPS (1 mM) was initially added to the IWB internal solution and substantially reversed the loss of current over the ground electrodes (Fig. 6A). DMPS also partially prevented the loss of current when added only to the external solution. Loss of current over the ground electrodes was prevented completely when DMPS was added to both the internal and external solutions (Fig. 6A), resulting in a uniform distribution of relative currents at the end of the depolarized phase (Fig. 6B). The effect on current uniformity was similar when DMPS concentration was reduced to 0.1 mM DMPS (data not shown). Importantly, DMPS did not interfere with the quality of the recordings as evidenced by seal resistances of 57.5±3.4 MΩ (n=9) versus 63.1±2.5 MΩ (n=10), success rates of 88%±9% (n=9) versus 93%±5% (n=10), and plate z-scores of 0.67±0.04 (n=9) versus 0.60±0.04 (n=10) in the presence and absence of 1 mM DMPS, respectively.

DMPS restores uniformity of Cav2.2 currents recorded on IonWorks Barracuda.
DMPS Does Not Affect the Pharmacology of Cav2.2 Blockers
To examine potential effects of DMPS on the pharmacology of Cav2.2 channels, we titrated a set of well-characterized Cav2.2 blockers in the presence of DMPS. IC50 values obtained on IWB in the presence of DMPS generally agreed well with published data, suggesting that DMPS does not alter channel pharmacology (Table 3). The potency of ω-conotoxin MVIIA appears to be underestimated on IWB. This is most likely a limitation of the exposure interval not allowing enough time for block to reach steady state, rather than an effect of DMPS. Where possible, literature data represent manual electrophysiology experiments performed on recombinantly expressed Cav2.2 channels with barium as the charge carrier.
Potency of Reference Cav2.2 Blockers in the IWB Assay
Values represent mean ± standard error of the mean (n).
Literature IC50 values are electrophysiology data. Preference is given to manual electrophysiology performed on recombinant channels when available.
Estimated based on single concentration data.
Discussion
Voltage-gated calcium channels and, in particular, Cav2.2 are attractive drug targets. Although robust and reproducible screening assays can be developed based on fluorescent calcium indicator dyes, 19 –21 a more detailed characterization of the mechanism of channel inhibition requires reliable voltage control. Traditionally, the characterization of voltage- and use-dependent blocking mechanisms has been performed using manual electrophysiological techniques; however, higher throughput alternatives are required for target-based drug discovery. In recent years, a number of medium- to high-throughput automated electrophysiology instruments have been developed. 22 These instruments are now increasingly used for test compound optimization, cardiac safety testing, 23 and in some cases for primary screening. 8 Despite significant progress, developing automated electrophysiology assays for voltage-gated calcium channels has been challenging. 24 –26
Recently, assays for several voltage-gated calcium channels, including Cav2.2, have been developed on the IWB platform. 10 These assays used a combination of conditioning prepulses and pulse trains from a hyperpolarized holding potential to assess voltage- and use-dependent block of calcium channels by a few reference compounds. Although results obtained for the reference compounds were in good agreement with expectations based on literature values, the authors acknowledge that the assay protocol may not be ideal for all classes of calcium channel blockers. In particular, the relatively short duration of the depolarizing prepulse may not allow sufficient time for compounds with slower kinetics to reach steady state. A protocol that continuously voltage clamps cells at a holding potential associated with 40%–60% inactivation would mitigate against this potential shortcoming. In the study by Kuryshev et al., 10 extensive current run-down precluded this type of voltage protocol.
In this study, we present the development of a high-throughput electrophysiology assay for Cav2.2 on the IWB platform that combined continuous voltage clamp at two holding potentials. Holding potentials correspond to full availability of channels in the resting state and ∼40% channel inactivation, respectively. Using this protocol, we demonstrate good current stability and pharmacological validation. One issue we observed was loss of Cav2.2 current above the ground electrodes. We show that Cav2.2 channels are blocked by silver ions and that block is reversed in the presence of the heavy metal chelating agent DMPS. The addition of DMPS in the recording solutions prevented the nonuniformity of currents across the patchplate, especially in the areas above the ground electrodes. Importantly, DMPS did not affect calcium channel pharmacology.
Our data are consistent with a literature report showing that Na,K-ATPase is inhibited by low micromolar concentrations of silver and that block is reversed in the presence of DMPS. 27 Block of Cav3.1 by AgNO3 has been reported. 16 Similar to our data for Cav2.2, the inhibition of Cav3.1 was not voltage dependent and current kinetics was not altered; however, the potency of AgNO3 at Cav3.1 was substantially lower (<50% block at 10 μM).
Although our data are consistent with silver ions causing the Cav2.2 current loss above the ground electrodes in the IWB assay, we cannot rule out that block may be caused by other heavy metals present in the AgCl pellet, such as cadmium or nickel, which would also be chelated by DMPS. 17
Finally, although the use of DMPS in our assay buffers did not affect the quality of our recordings, we cannot rule out effects of the long-term use of DMPS on electrode performance. Therefore, we recommend using the lowest concentration of DMPS that will minimize ground electrode effects and restore uniformity to current measurements on IWB. In our recordings of rat Cav2.2, this concentration was 0.1 mM; however, details of the experimental protocol, including the cell line, holding potential, and length of the recording, may affect the optimal DMPS concentration.
Footnotes
Disclosure Statement
All authors are employees of Eli Lilly and Company.