
Abstract
The association between the kainate receptors (KARs) GluK1 and GluK2 and the modifying proteins neuropilin- and tolloid-like 1 (NETO1), neuropilin- and tolloid-like 2 (NETO2), and postsynaptic density protein 95 (PSD95) is likely to produce distinct GluK1 and GluK2 pharmacology in postsynaptic neurons. However, little is known about their corresponding modulatory effects on GluK1 and GluK2 activity in high-throughput assays for cell-based drug discovery. Using heterologous cells that potentially mimic the response in native cells in a fluorescence imaging plate reader (FLIPR) assay, we have investigated assays that incorporate (1) coexpression of GluK1 or GluK2 with their modulatory proteins (NETO1, NETO2, PSD95) and/or (2) enablement of assays with physiological concentration of native GluK1 and GluK2 agonist (glutamate) in the absence of an artificial potentiator (e.g., concanavalin A [Con A]). We found that in the absence of Con A, both NETO1 and NETO2 accessory proteins are able to potentiate kainate- and glutamate-evoked GluK1-mediated Ca2+ influx. We also noted the striking ability of PSD95 to enhance glutamate-stimulated potentiation effects of NETO2 on GluK1 without the need for Con A and with a robust signal that could be utilized for high-throughput FLIPR assays. These experiments demonstrate the utility of heterologous cells coexpressing PSD95/NETO2 with GluK1 or GluK2 in native cell-mimicking heterologous cell systems for high-throughput assays and represent new avenues into the discovery of KAR modulating therapies.
Introduction
Ionotropic glutamate receptors are glutamate-gated ion channels that mediate the majority of fast excitatory synaptic transmission in the central nervous system (CNS) and are involved in numerous physiological and pathophysiological processes.
1
These receptors consist of three different classes: N-methyl-
In recent years, progress has been made in understanding the auxiliary subunits associated with native KARs. 5 Several proteins such as postsynaptic density protein 95 (PSD95), 6,7 neuropilin tolloid-like 1 & 2 (NETO1 & 2), 8 kainate receptor-interacting protein for GluR6 (KRIP6), 9 protein interacting with C-kinase 1 (PICK1), 10 and glutamate receptor-interacting protein 1 (GRIP1) 10 have been identified as KAR interacting proteins, but only NETO1 and NETO2 were recently generally accepted as KAR auxiliary proteins. 5,8 It remains largely unknown how these proteins regulate the activities of KAR pharmacological modulation agents.
PSD95 is a major scaffolding protein enriched at glutamatergic synapses. 11 –16 Literature evidence 17,18 suggests that PSD95 may play a key role in modulating clustering of neurotransmitter receptors, adhesion molecules, ion channels, cytoskeletal elements, and signaling molecules at postsynaptic sites. A role for PSD95 in regulating AMPA receptor insertion and retention at the synapse has been well documented by numerous studies. 13 –16,19 –21 PSD95 has been shown to influence surface expression of NMDA receptors and receptor desensitization, suggesting a role for NMDA receptor-PSD-95 interaction in glutamate-mediated signaling processes. 22 –24
With electrophysiological recordings, it was demonstrated that coexpression of PSD95 with GluK2 in transfected cells modulates GluK2 receptor function by either strongly reducing desensitization 6 or accelerating desensitization recovery, 7 which was explained by GluK2 clustering through binding to PDZ1 domain of PSD95. However, it is unclear how PSD95 modulates GluK1- or GluK2-mediated calcium influx in high-throughput assays and if coexpression of PSD95 and GluK1 or GluK2 will affect the potencies of pharmacological modulation agents for GluK1 or GluK2.
NETO1 and NETO2 are brain-specific CUB (protein domains of C1r/C1s, Uegf, and Bmp1) domain-containing transmembrane proteins. Previous reports suggest that NETO1 is a component of the NMDAR complex, critical for maintaining the abundance of NR2A-containing NMDARs in the postsynaptic density. 25,26 Furthermore, recent studies demonstrate that both NETO1 and NETO2 are auxiliary subunits that complex with native KARs to modulate their function. 5,27,28 With electrophysiological recordings, it was demonstrated by several groups 27 –34 that coexpression of GluK1 or GluK2 with either NETO1 or NETO caused the substantial increase in glutamate sensitivity and alterations of desensitization onset, or recovery of KARs.
KARs are crucial for some forms of fundamental synaptic plasticity in the CNS 35 and have been implicated in a wide range of neurological and psychiatric disorders 36,37 ; thus, there is major interest in identifying new pharmacological agents that target these receptors. 38,39 Exploring the interaction of KARs with their auxiliary subunits may represent attractive opportunities to find pharmacological modulation agents for KARs.
As mentioned above, the modulation of KAR kinetic properties by NETO1, NETO2, and PSD95 was documented in the literature, but the potential applications of different compositions of these subunits in drug discovery is largely unknown. It was well known that synaptic KARs display properties that differ from recombinant receptors. In particular, the slow decay of KARs synaptic currents contrasts with the rapid deactivation and desensitization of KARs expressed in heterologous cells. 27 –33,40
We hypothesize that the coexistence of NETO1, NETO2, and PSD95 with GluK1 and GluK2 in postsynaptic neurons 5 –7,28,41,42 may have distinct effects on GluK1 or GluK2 pharmacology such as potency alterations of KAR agonist and antagonist compounds. The potential distinct pharmacology was not assessed in previous heterologous cell-based drug discovery screening assays where only GluK1 or GluK2 alone was expressed.
In this study, we have explored the effect of NETO1, NETO2, and PSD95 on GluK1 and GluK2 responses to kainate- or glutamate-evoked challenges. We discovered that NETO1, NETO2, and NETO2/PSD95 are able to modulate GluK1- and GluK2-mediated calcium influx in different ways. Strikingly, we observed that physiologically relevant combinations of GluK1 or GluK2 with NETO1 or NETO2 or PSD95/NETO2 resulted in distinct pharmacological effects and enabled GluK1 and GluK2 high-throughput assays within heterologous cell systems.
Interestingly, to reduce agonist-dependent desensitization of the ion channel, it has been necessary to include an artificial potentiator (e.g., concanavalin A [Con A]) in previous versions of high-throughput GluK1 and GluK2 compound screening assays with a heterologous cell expressing GluK1 or GluK2 alone. 43 Con A does not exist in native cells and is not required in these assays. Thus the findings in this report and subsequent assay enablement may present new avenues into the discovery of KAR modulating therapies.
Materials and Methods
Constructs
Human GluK1 (transcript variant 2, NM_175611.2) and GluK2 (transcript variant 1, NM_021956.4) complementary deoxyribonucleic acids (cDNAs) were cloned from a human brain cDNA library by polymerase chain reaction (PCR) and subcloned into the pcDNA3.1-hygromycine vector. There are two variants of amino acids on position 621 of both GluK1 and GluK2–arginine (R) or glutamine (Q). 44 –46 Upon glutamate stimulation, R versions are least active and Q versions are more active GluK1 or GluK2 receptors because the presence of the positively charged amino acid (R) in the inner channel pore makes the receptor channel impermeable to Ca2+. 46
To generate stable cell lines expressing highly active GluK1 or GluK2 receptors, we generated versions of GluK1 (GluK1Q) and GluK2 (GluK2Q) stable cell lines incorporating glutamine (Q) in these studies. NETO1, NETO2, and PSD95 cDNA were cloned from a human brain cDNA library and subcloned into pcDNA3.1-neomycin (NETO1, NETO2, and PSD95) and pcDNA3.1-zeocin vectors (NETO2). To detect expression of NETO1 and NETO2, constructs of pcDNA3.1-neomycine-NETO1-HA and pcDNA3.1-NETO2-HA constructs were also generated in which hemagglutinin (HA) (YPYDVPDYA) tag coding sequences were inframe cloned to 3′-end of NETO1 and NETO2.
Cell Culture and Generation of Stable Cell Lines
Monolayer culture of human embryonic kidney 293 (HEK293) cells was grown at 37°C in a humidified air atmosphere with 5% CO2 using DMEM/F12 (3:1; Invitrogen, Carlsbad, CA) supplemented with 2 mM Glutamax (Invitrogen), 20 mM HEPES (Invitrogen), 10% heat-inactivated fetal bovine serum (Invitrogen), and 1% penicillin–streptomycin solution (Invitrogen). HEK293 cell-based stable cell lines were cultured in the same medium with the addition of appropriate selection antibiotics dependent on the expression vector used.
The HEK293/GluK1Q stable cell line was generated by transfecting the pDNA3.1-GluK1Q plasmid into HEK293 cells with Fugene 6 (Roche Diagnostics, Indianapolis, IN) according to the manufacturer's instructions. Forty-eight hours after transfection, cells were divided and cultured with a HEK293 cell-culturing medium containing hygromycin (0.5 mg/mL) for 2 weeks. Individual clones were selected and confirmed for expression of functional GluK1Q by their response to glutamate-evoked Ca2+ influx with a FLIPR assay (described below). The HEK293/GluK1Q stable cell was maintained in a medium containing 0.25 mg/mL hygromycin.
The HEK293/GluK1Q/NETO1 stable cell line was generated by transfecting pcDNA3.1-NETO1-HA plasmid into cells stably expressing functional GluK1Q. Transfected cells were selected with HEK293 cell-culturing medium containing hygromycin (0.25 mg/mL) and neomycin (1 mg/mL) for 2 weeks. Individual clones were selected, and expression of NETO1 in HEK293/GluK1Q/NETO1 cells was confirmed by Western blot with HA antibody (Roche Diagnostic). The HEK293/GluK1Q/NETO1 stable cells were maintained in a HEK293 cell-culturing medium containing hygromycin (0.25 mg/mL) and neomycin (0.25 mg/mL).
HEK293/GluK1Q/NETO2 and HEK293/GluK1Q/PSD95 stable cell lines were generated with the same procedure previously described to generate the HEK293/GluK1Q/NETO1 stable cell line with the pcDNA3.1-NETO2-HA or pcDNA3.1-PSD95 plasmid being utilized, respectively.
Expression of NETO2 in HEK293/GluK1Q/NETO2 cells was also confirmed by Western blot with the HA antibody. Expression of PSD95 in HEK293/GluK1Q/PSD95 cells was confirmed by Western blot with a PSD95 antibody (Abcam, Boston, MA).
The HEK293/GluK1Q/PSD95/NETO2 stable cell line was generated by transfecting pcDNA3.1-zeo/NETO2-HA plasmid into an established stable cell line containing HEK293/GluK1Q/PSD95. Transfected cells were selected with the HEK293 cell-culture medium containing hygromycin (0.25 mg/mL), neomycin (0.25 mg/mL), and zeocin (0.1 mg/mL) for 2 weeks. Individual clones were picked, and expression of NETO2 and PSD95 in HEK293/GluK1Q/PSD95/NETO2 stable cells was confirmed by Western blot with HA and PSD95 antibodies, respectively. The HEK293/GluK1Q/PSD95/NETO2-HA stable cells were maintained in a HEK293 cell-culture medium containing hygromycin (0.25 mg/mL), neomycin (0.25 mg/mL), and zeocin (0.05 mg/mL).
The stable cell-line generation procedure described previously was applied to generate stable lines of HEK293/GluK2Q, HEK293/GluK2Q/NETO1, HEK293/GluK2Q/NETO2, HEK293/GluK2Q/PSD95, and HEK293/GluK2Q/PSD95/NETO2.
Western Blot
Cell pellets were collected from confluent cultures. Whole cell lysates were prepared by suspending cell pellets in an ice-cold cell lysis buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 1% Triton, 2.5 mM sodium pyrophosphate, 1 mM beta-glycerophosphate, 1 mM Na3VO4, 1 μg/mL leupeptin, 1 mM PMSF) and sonicating for 30 s on ice. Cell homogenates were kept on ice for 30 min and then centrifuged at top speed (14,000 rpm) in an Eppendorf microcentrifuge at 4°C for 30 min. Supernatant was collected as whole cell lysate and protein concentration of the samples was determined using the BCA Protein Assay (Pierce, Rockford, IL). For Western blot analysis, the whole cell lysate was fractionated on a 10% NuPAGE Bis-Tris gel and transferred onto a 0.2 μM nitrocellulose membrane (Novex, San Diego, CA) using a Hoefer Transblotter (Semiphor, San Francisco, CA).
The membrane was washed twice for 5 min each in TST (10 mM Tris-HCl, 150 mM NaCl, 0.1% Tween-20, pH 7.5), blocked with 5% bovine serum albumin–TST for 1 h at room temperature (RT) on a shaker platform (Lab-Line), and then incubated with the respective primary antibodies overnight at 4°C. GluK1Q, GluK2Q, and β-actin antibodies were purchased from Millipore (Bedford, MA), respectively. On the next day, the membrane was washed thrice for 5 min each with TST. An appropriate HRP (Cell Signaling Technology, Boston MA)-conjugated secondary antibody was diluted 1:2,000 in 5% milk–TST and incubated with the membrane for 1 h, shaking at RT. Finally, the membrane was washed as described above, followed by a single wash with phosphate-buffered saline for 5 min. Respective protein on the membrane was then visualized by chemiluminescence (Amersham Bioscience, Piscataway, NJ).
Fluorescence Imaging Plate Reader Assay
Glutamate-evoked GluK1Q or GluK2Q receptor-mediated Ca2+ influx in various GluK1Q- or GluK2Q-containing stable cell lines was measured using a fluorescence imaging plate reader (FLIPR) assay (all FLIPR signals mean Ca2+ influx hereafter). Cells were plated at 6 × 104 cells per well in black 96-well clear-bottom plates (Becton Dickinson, Bedford, MA) and incubated 40–48 h at 37°C to achieve 80%–100% confluence. The cell-culture medium was removed, and cells were washed once with 100 μL/well of an assay buffer (HBSS supplemented with 10 mM HEPES and 3.7 mM CaCl2). Cells were loaded with FLIPR dye fluo-4 (Invitrogen) in a mixture containing 17 μL of 4 mM Fluo-4 (dissolved in DMSO), 17 μL of 20% pluronic acid in 20% DMSO (Invitrogen), 2 mL of Hi-FBS (Invitrogen), and 18 mL of assay buffer for 90 min at RT in the dark.
Loaded dye was removed, and cells of each well were washed once with 100 μL of assay buffer. A 50 μL volume of assay buffer or Con A (GluK1Q and GluK2Q artificial potentiator) (Sigma, St. Louis, MO, 0.025% in assay buffer) was added to each well with Te-Mo (Tecan, Morrisville, NC) and plates were kept at RT for 15 min in the dark. Subsequent addition of reagents and measurements was conducted by a programmable FLIPR instrument (Molecular Device, Sunnyvale, CA). A two addition program was employed for all FLIPR measurements. Fifty microliter of assay buffer was added to each well as the first addition. To generate glutamate or kainate concentration-response curves (CRC), solutions containing a range of 2× glutamate or kainate concentrations were made in the assay buffer and 100 μL of each concentration was added to each well as the second addition.
The resulting fluorescence was quantified using the FLIPR instrument at an excitation wavelength of 488 nm and an emission wavelength of 540–590 nm set by a bandpass filter obtained from Molecular Devices. Both additions were composed of a baseline imaging (5 images with 1 s interval), the first interval signal imaging (20 images with 1 s interval) and the second interval signal imaging (60 images with 6 s interval). Thus, total measurement time course was 370 s, 185 s for each addition. All assays were carried out at RT. The FLIPR protocol used in this study is summarized in Table 1.
Protocol for Fluorescence Imaging Plate Reader Assay
1. Naive HEK293 cells or HEK293 cells expressing various combinations of GluK1Q or GluK2Q with NETO1, NETO2, or PSD95 cells were plated in poly-
2. After plating cells into plates, keep plates in room temperature for 1 h, and then transfer to 37°C incubator with 90% humidity and 5% CO2.
3. The assay buffer is H8SS supplemented with 10 mM HEPES and 3.7 mM CaCl2. Add the assay buffer into wells, and then dump it off.
4. FLIPR dye mixture is composed of 4 mM Fluo4 in DMSO (17 μL), 20% pluronic acid in DMSO (17 μL), FBS (2 mL), and an assay buffer (18 mL).
5. Same as step 3.
6. Fifty microliters of 0.025% Con A in the assay buffer or control (assay buffer) was added in each well.
7. Run signal test to make signals between 80,000 and 10,000 fluorescence counts by adjusting exposure length, camera gain or laser strength.
8. Fifty microliters of assay buffer was added in each well by the FLIPR instrument.
9. Half-logdiluted glutamate or kainate(2×) in the assay buffer was added in each well by the FLIPR instrument.
10. Excition and emission.
FLIPR, fluorescence imaging plate reader; HEK293, human embryonic kidney 293; NETO1, neuropilin- and tolloid-like 1; NETO2, neuropilin- and tolloid-like 2; PSD95, postsynaptic density protein 95.
Quantitative Reverse Transcription–Polymerase Chain Reaction
Total ribonucleic acid (RNA) was isolated from cells using Qiagen's RNeasy mini kit by following the manufacturer's instruction. First-strand cDNA was synthesized using 1 μg of total RNA according to the manufacturer's protocol (ImPromII Reverse Transcription System; Promega). Three independent samples from each cell line were used for quantitative reverse transcription-PCR (qRT-PCR) analyses with primer/probe sets (NETO1, Hs0037151_m1; NETO2, Hs00983152_m1) and PCR Master Mix purchased from Applied Biosystems on an Applied Biosystems model 9700 Sequence Detector thermal cycler and normalized to 18S-RNA expression. Data were expressed as ddCT.
Data Analysis
The maximum peak signal − minimum signal (baseline) was exported and used for all FLIPR data analysis in this report. The assay unit for all FLIPR signals is fluorescence counts.
Since the first addition was the assay buffer and agonists (kainate or glutamate) for GluK1 and GluK2 were added only in the second addition, all FLIPR signal data were exported from image 86–170 for data analysis. Exported FLIPR data of CRC generated with either kainate or glutamate in various cell lines were analyzed by the iterative fitting program within GraphPAD Prism (GraphPAD Software). Log concentration-effect curves were fitted to logistic expressions for single-site analysis.
Results
Generation and Confirmation of GluK1Q and GluK2Q Stable Cell Lines
To study the potential modulation by NETO1, NETO2, and PSD95 on the rapid desensitization of GluK1 and GluK2 channels, HEK293 cell-based stable lines expressing GluK1Q, GluK2Q, NETO1, NETO2, PSD95, and different combinations of each were generated. The expression of GluK1Q, GluK2Q, NETO1, NETO2, and PSD95 in each line was confirmed by Western blot analysis (Fig. 1A, B).

Western blot confirmation of GluK1, GluK2, NETO1, NETO2, and PSD95 expression in various HEK293 stable cell lines as labeled in each panel.
There were no reliable commercially available antibodies available for either NETO1 or NETO2. To detect expression of NETO1 and NETO2 in cell lines containing either NETO1 or NETO2, HA was tagged to the C-terminal of NETO1 or NETO2. An antibody that recognized HA was used to confirm the expression of NETO1 and NETO2 in appropriate cell lines as indicated in Figure 1A and B.
Expression of functional GluK1Q and GluK2Q channels in GluK1Q- or GluK2Q-containing cell lines was confirmed by FLIPR assays (data presented below). To determine whether the HA tag interferes with the function of NETO1 or NETO2 in modulating GluK1 and GluK2 channel properties, stable cell lines coexpressing GluK1Q/NETO1 (no tag), GluK1Q/NETO2 (no tag), GluK2Q/NETO1 (no tag), and GluK2Q/NETO2 (no tag) were also generated. We compared cell lines expressing either GluK1Q or GluK2Q side by side, with or without HA-tagged NETO1 and NETO2 and observed no pharmacological differences in responses to either kainate or glutamate stimulation in FLIPR assays (data not shown).
Potentiating Effects of NETO1 and NETO2 on GluK1Q in Responding to Kainate or Glutamate Stimulation in the Presence or Absence of Con A
Kainate-evoked GluK1Q and GluK2Q currents (as measured electrophysiologically) decay in tens or hundreds of milliseconds. 47 –49 GluK1Q or GluK2Q expressed in heterologous cell systems also deactivates and desensitizes rapidly after glutamate stimulation. 1,50 Due to these characteristics, in-vitro studies utilizing GluK1 and GluK2 cell lines to characterize potential pharmacological agents require the presence of potentiators to reduce or remove the desensitization of GluK1 or GluK2 and thus enable the measurement of pharmacological responses in high-throughput assays such as FLIPR.
The most common potentiator used in GluK1 or GluK2 FLIPR assays is the plant lectin, Con A. 43 However, in GluK1 and GluK2 pharmacological screening studies, high-throughput assays utilizing Con A were a concern regarding the translatability of compound potency and efficacy from the heterologous cell systems to native cells and ultimately to humans. Thus, we sought to enable GluK1 and GluK2 high-throughput assays in the absence of Con A for drug characterization studies.
NETO1 and NETO2 were recently reported as homomeric GluK2 receptor modulators with the possible potentiation of GluK1- and GluK2-channel kinetic properties. 5,27,29,30,34 In this study, we evaluated whether NETO1 or NETO2 was capable of modulating agonist responses to kainate and glutamate in GluK1Q- or GluK2Q-expressing cells, thus enabling GluK1Q and GluK2Q cell assays to be used in high-throughput assays in the absence of Con A.
In the presence of Con A, compared to GluK1Q alone, neither NETO1 nor NETO2 modulated kainate-evoked GluK1Q-mediated FLIPR signals (Fig. 1A). However, in the absence of Con A, both NETO1 and NETO2 were able to potentiate kainate-evoked GluK1Q-mediated FLIPR signals (increased about two to threefold) (Fig. 2B).
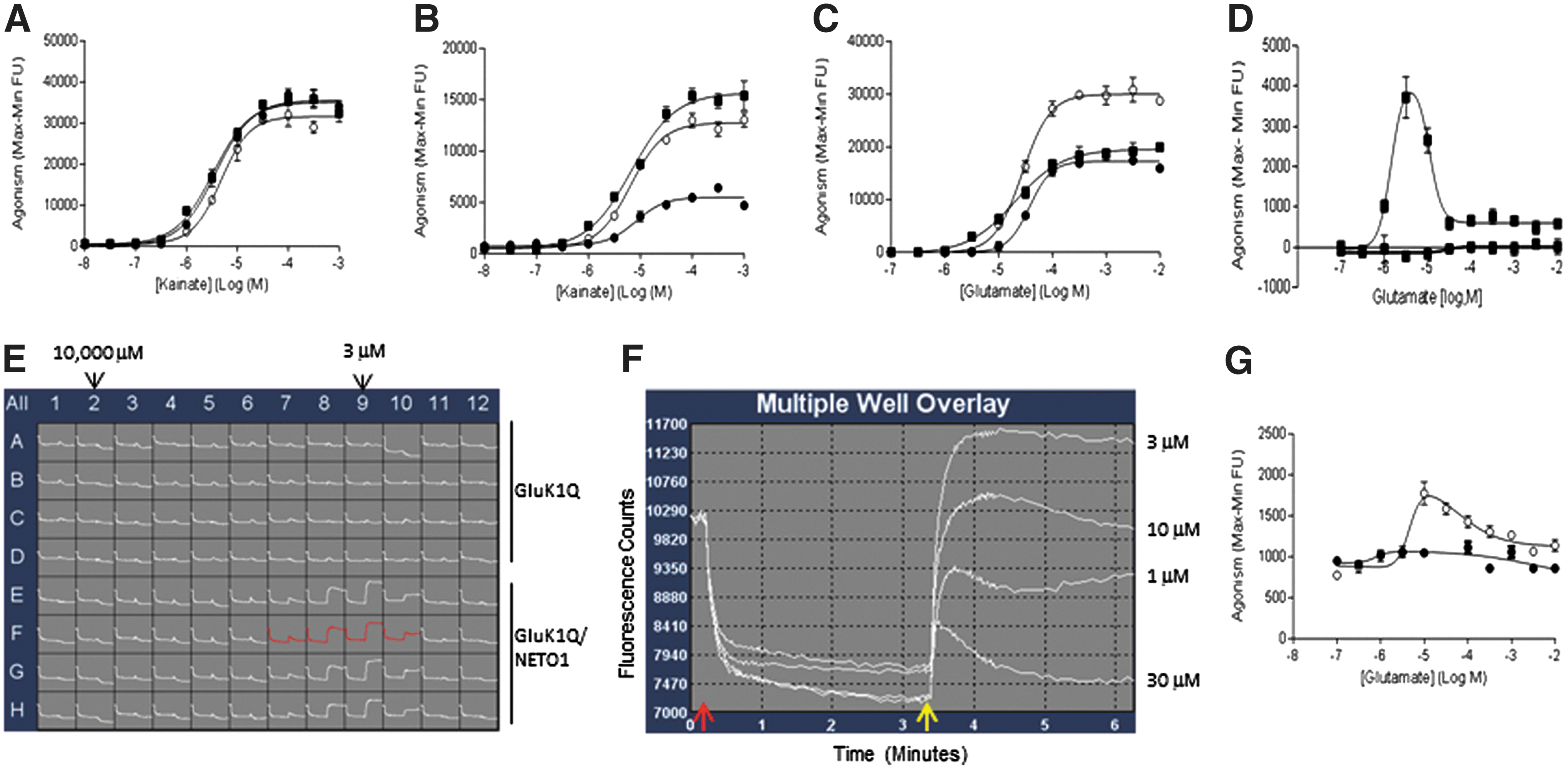
Results from FLIPR assays to measure modulatory effects of NETO1 and NETO2 on kainate or glutamate response in the presence or absence of Con A in cell lines expressing GluK1Q alone (●), GluK1Q/NETO1 (■), and GluK1Q/NETO2 (○).
Interestingly, lower magnitude kainate-evoked GluK1Q-mediated FLIPR signals were detected in HEK293/GluK1Q-alone cells in the absence of Con A. Our original hypothesis that might explain this observation was that there may be basal levels of endogenous NETO1 and/or NETO2 expression in host HEK293 cells. Indeed, basal levels of NETO1 and NETO2 expression were detected in HEK293-naive cells with quantitative RT-PCR (Fig. 3A, B).
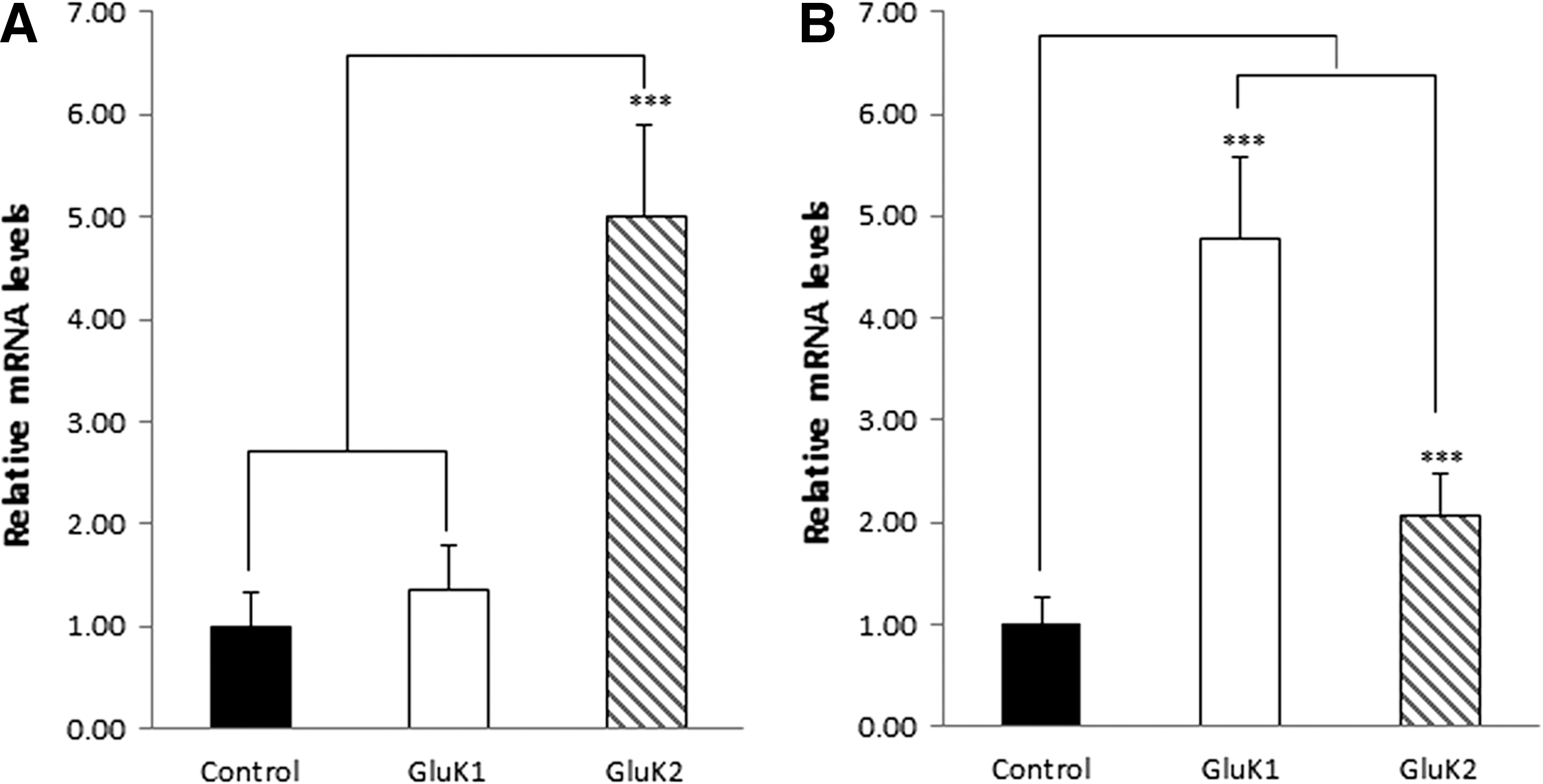
Quantitative reverse transcription–polymerase chain reaction analysis of endogenous NETO1 and NETO2 expression in various HEK293 cell lines as labeled in each panel. Controls were naive HEK293 cells. Expression values of NETO1 or NETO2 are relative to naive HEK293 (black column).
However, further investigation suggested that expression of GluK1Q significantly increased endogenous expression levels of NETO2 (Fig. 3B), but NETO1 (Fig. 3A) in HEK293 cells through an unknown mechanism. Thus, the kainate-evoked GluK1Q-mediated FLIPR signals detected in HEK293/GluK1Q cells mainly represented synergism of GluK1Q with NETO2. Interestingly, neither NETO1 nor NETO2 significantly changed the potency of kainate on GluK1Q in the presence or absence of Con A (Table 2) in FLIPR assays.
Comparison of EC50 Values of Kainate and Glutamate on Glukl2q and Gluk2q With or Without NETO1 and NETO2
Data are reported as mean ± SEM (n = 3).
No EC50 reported; no concentration-dependent curves observed.
Unlike glutamate, kainate is not an endogenous GluK1 ligand in humans. Therefore, to enable a GluK1 high-throughput assay that more closely mimics the native state, we explored the effect of NETO1 or NETO2 on potentiating glutamate-evoked GluK1Q-mediated FLIPR signals in the presence or absence of Con A. In the presence of Con A, NETO2, but not NETO1, was able to potentiate glutamate-evoked GluK1Q-mediated FLIPR signals (Fig. 2C). In the absence of Con A, both NETO1 (Fig. 2D–F) and NETO2 (Fig. 2G) were able to slightly potentiate glutamate-evoked GluK1Q-mediated FLIPR signals with a “bell-shaped” response curve.
The glutamate dose–response curves shown in Figure 2D were data analysis of the FLIPR traces presented in Figure 2E. The response traces in well F7–10 in Figure 2E were shown in Figure 2F. The FLIPR signals of maximum–minimum (max-min) between the starting image of the second addition (as pointed by the yellow arrow in Fig. 2F) to the last image were exported for data analysis. As shown in Figure 2F, each glutamate concentration had its own minimum line. The max–min data were exported with the maximum and minimum signals that were glutamate concentration specific.
For the GluK1Q/NETO1 (Fig. 2D–F) and GluK1Q/NETO2 (Fig. 2G) assays, peak response was achieved at 3 μM and 10 μM glutamate, respectively. These peak responses to glutamate were confirmed in three or more repeated experiments for each assay, suggesting that GluK1Q/NETO1 assay is slightly more sensitive to responding glutamate stimulation than GluK1Q/NETO2 assay in the absence of Con A. The signal/noise ratios at peak responses to glutamate for both GluK1/NETO1 (Fig. 2F) and GluK1/NETO2 (trace data is not shown) assays were less than twofolds.
Potentiating Effects of NETO1 and NETO2 on Gluk2Q in Response to Kainate or Glutamate Stimulation in the Presence or Absence of Con A
Similar studies were performed utilizing the GluK2Q FLIPR assays. In the presence of Con A, compared to GluK2Q alone, both NETO1 and NETO2 were able to slightly potentiate kainate-evoked GluK2Q-mediated FLIPR signals (Fig. 4A). In the absence of Con A, NETO1, but not NETO2, was able to robustly potentate kainate-evoked GluK2-mediated FLIPR signals (increased about three-fold) (Fig. 4B). Quantitative RT-PCR data suggested that GluK2Q expression increased endogenous NETO2 expression about onefold (Fig. 3B), but increased NETO1 expression about fourfold (Fig. 3A). The significant, but relatively less-increased endogenous NETO2 expression due to GluK2Q expression did not have a notable potentiating effect on GluK2Q in responding to kainate stimulation in the absence of Con A.
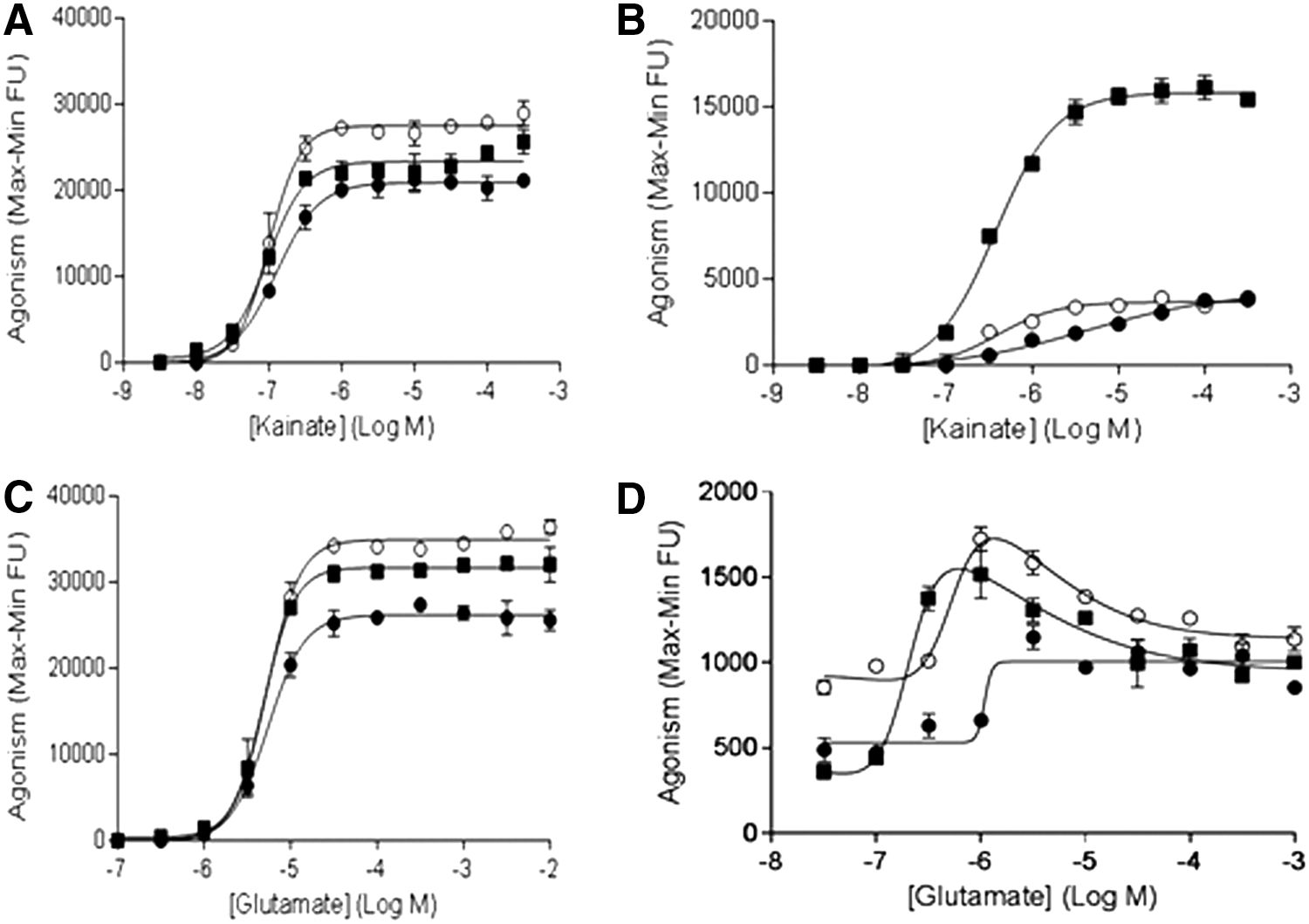
Results from FLIPR assays to measure modulatory effects of NETO1 and NETO2 on GluK2Q in responding to kainate or glutamate stimulations in the presence or absence of Con A among cell lines of GluK2Q alone (●), GluK2Q/NETO1 (■), and GluK2Q/NETO2 (○).
However, the Hill slope of the kainate-response curve in HEK293/GluK2Q cells (0.56) was different from the Hill slope obtained with HEK293/GluK2Q/NETO2 cells (1.24). This difference in Hill slope resulted in a kainate EC50 value on GluK2Q/NETO2 that was about 10 times lower compared with GluK2Q alone (Fig. 4B).
These results suggest that different expression levels of NETO2 in cells (e.g., HEK293/GluK2Q/NETO2 vs. HEK293/GluK2Q) affected the sensitivity of GluK2 to kainate stimulation. Overall, in the absence of Con A, coexpression of NETO1 or NETO2 with GluK2Q decreased the EC50 value for kainate compared to values obtained in the presence of Con A (Table 2).
Similar to kainate, in the presence of Con A, compared to GluK2Q alone, both NETO1 and NETO2 were able to slightly potentiate glutamate-evoked GluK2Q-mediated FLIPR signals (Fig. 4C). In the absence of Con A, both NETO1 and NETO2 (Fig. 4D) were able to slightly potentiate glutamate-evoked GluK2Q-mediated FLIPR signals in a “bell-shaped” manner. Peak glutamate responses were obtained with 1 μM glutamate for both GluK2Q/NETO1 and GluK2Q/NETO2. The signal/noise ratios at peak responses to glutamate for both GluK1/NETO2 and GluK1/NETO2 (trace data is not shown) assays were less than twofolds.
Potentiating Effects of PSD95 and PSD95/NETO2 on GluK1Q in Responding to Kainate or Glutamate Stimulations in the Presence or Absence of Con A
PSD95 has been reported to modulate the function of both AMPA and NMDA receptors. 20,51 In this study, we evaluated the ability of PSD95 to modulate glutamate and kainate response at GluK1. In the presence of Con A, neither PSD95 alone nor PSD95/NETO2 potentiated kainate-evoked GluK1Q-mediated FLIPR signals (Fig. 5A). Under these conditions, the potency of kainate is similar in cells expressing GluK1Q alone or GluK1Q/PSD95, GluK1Q/NETO2, or GluK1Q/PSD95/NETO2 (Table 3).
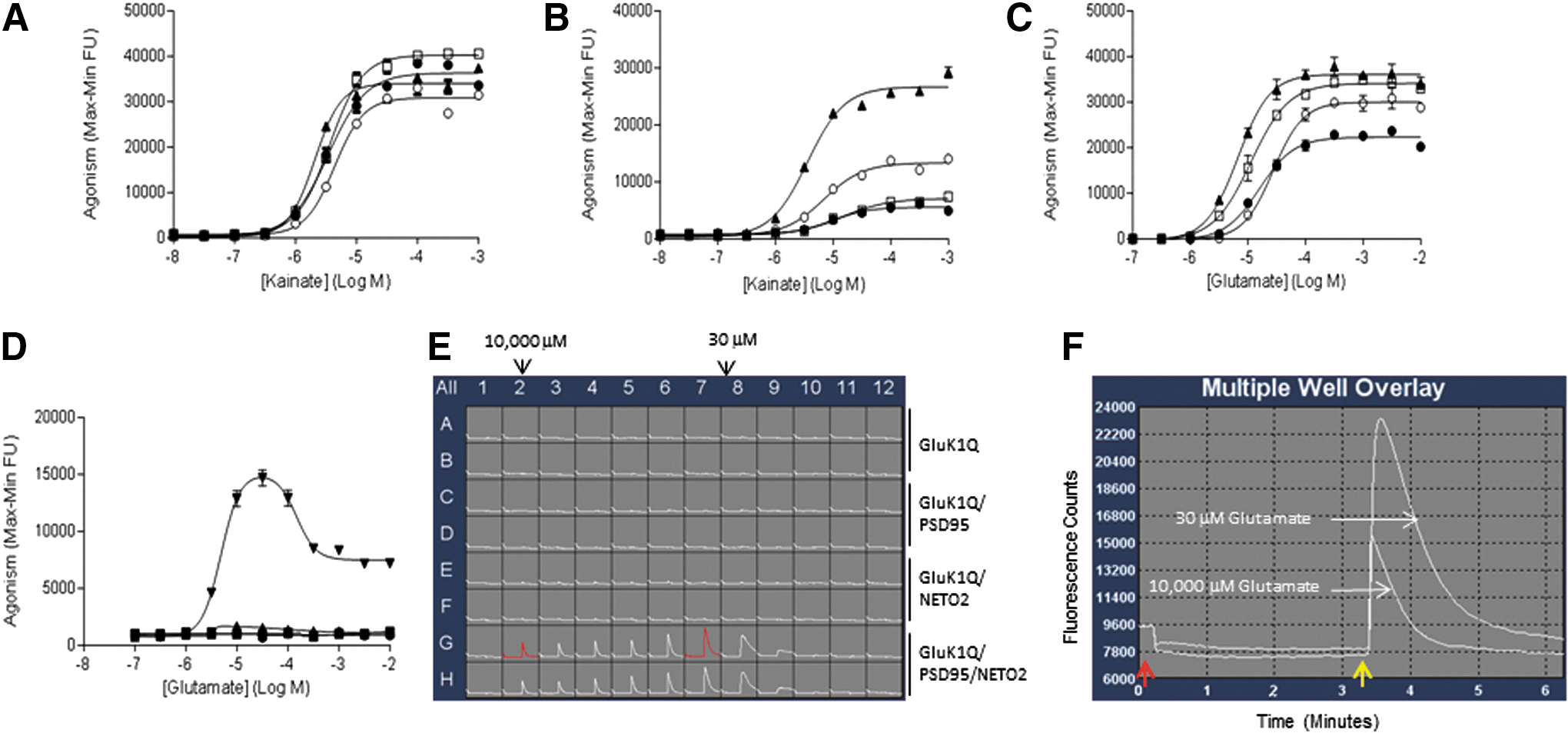
Responses in FLIPR assays to measure modulatory effects of PSD95 and PSD95/NETO2 on kainate- or glutamate-induced response in the presence or absence of Con A in cell lines expressing GluK1Q alone (●), GluK1Q/PSD95 (□), GluK1Q/NETO2 (○), and GluK1Q/PSD95/NETO2 (▲).
Comparison of EC50 Values of Kainate and Glutamate on Gluk1Q and Gluk2Q With or Without PSD95 or PSD95/NETO2
Data are reported as mean ± SEM (n = 3).
No EC50 reported; no concentration-dependent curves observed.
In contrast, in the absence of Con A, PSD95 alone could not potentiate the GluK1Q response to kainate stimulation (Fig. 5B), but could robustly enhance NETO2's potentiating effect on the kainate response at GluK1Q (Fig. 5B). Also, in the absence of Con A, kainate potency is highest in cells expressing GluK1Q/PSD95/NETO2 compared to peer cells (Table 3). Overall, when compared to the same cells expressing GluK1Q alone or GluK1Q/PSD95, GluK1Q/NETO2, or GluK1Q/PSD95/NETO2, kainate was less potent in the absence of Con A than in the presence of Con A (Table 3).
We further explored the potential modulating effects of PSD95 and PSD95/NETO2 on GluK1Q in response to glutamate stimulation. In the presence of Con A, in contrast to results obtained with kainate stimulation, PSD95 alone potentiated glutamate-evoked GluK1Q-mediated FLIPR signals (Fig. 5C). Coexpression of PSD95/NETO2 with GluK1Q had more robust potentiating effects on glutamate stimulation of GluK1Q than either PSD95 alone or NETO2 alone (PSD95/NETO2 > PSD95 > NETO2 > GluK1 alone). Interestingly, the potency of glutamate on GluK1Q had the opposite order to the potentiation order above (PSD95/NETO2 < PSD95 < NETO2 < GluK1 alone).
Strikingly, we found that in the absence of Con A, PSD95 robustly enhanced the potentiation effects of NETO2 on GluK1Q in response to glutamate stimulation, also with a “bell-shaped” response (Fig. 5D).
The bell-shaped response to glutamate stimulation in the absence of Con A observed with GluK1Q/PSD95/NETO2 was about sevenfold larger than results obtained from the combinations of either GluK1Q/NETO1 or GluK1Q/NETO2. A representative FLIPR trace of glutamate concentration responses in various GluK1Q cell lines is shown in Figure 5E. The highest glutamate response was observed at 30 μM (Fig. 5E) with >2.5-fold signal/noise (maximum/minimum) ratio (Fig. 5F).
Potentiating Effects of PSD95 and PSD95/NETO2 on GluK2Q in Response to Kainate or Glutamate Stimulation in the Presence or Absence of Con A
The potentiating effects of PSD95 and PSD95/NETO2 on GluK2-mediated calcium influx in response to kainate or glutamate stimulation were also explored in this study. Similar to GluK1, in the presence of Con A, neither PSD95 nor PSD95/NETO2 potentiated kainate-evoked GluK2Q-mediated FLIPR signals (Fig. 6A). In the absence of Con A, the potency of kainate was similar in cells expressing GluK2Q alone, GluK2Q/PSD95, GluK2Q/NETO2, or GluK2Q/PSD95/NETO2 (Table 3). Also, in the absence of Con A, neither PSD95 alone nor NETO2 alone was able to potentiate the effect of kainate at GluK2Q (Fig. 6B). However, PSD95 and NETO2 together robustly enhanced kainate-evoked GluK2Q-mediated FLIPR signals (Fig. 6B). Overall, when compared to cells expressing GluK2Q alone, GluK2Q/PSD95, GluK2Q/NETO2, or GluK2Q/PSD95/NETO2, kainate was less potent in the absence of Con A than in the presence of Con A (Table 3).
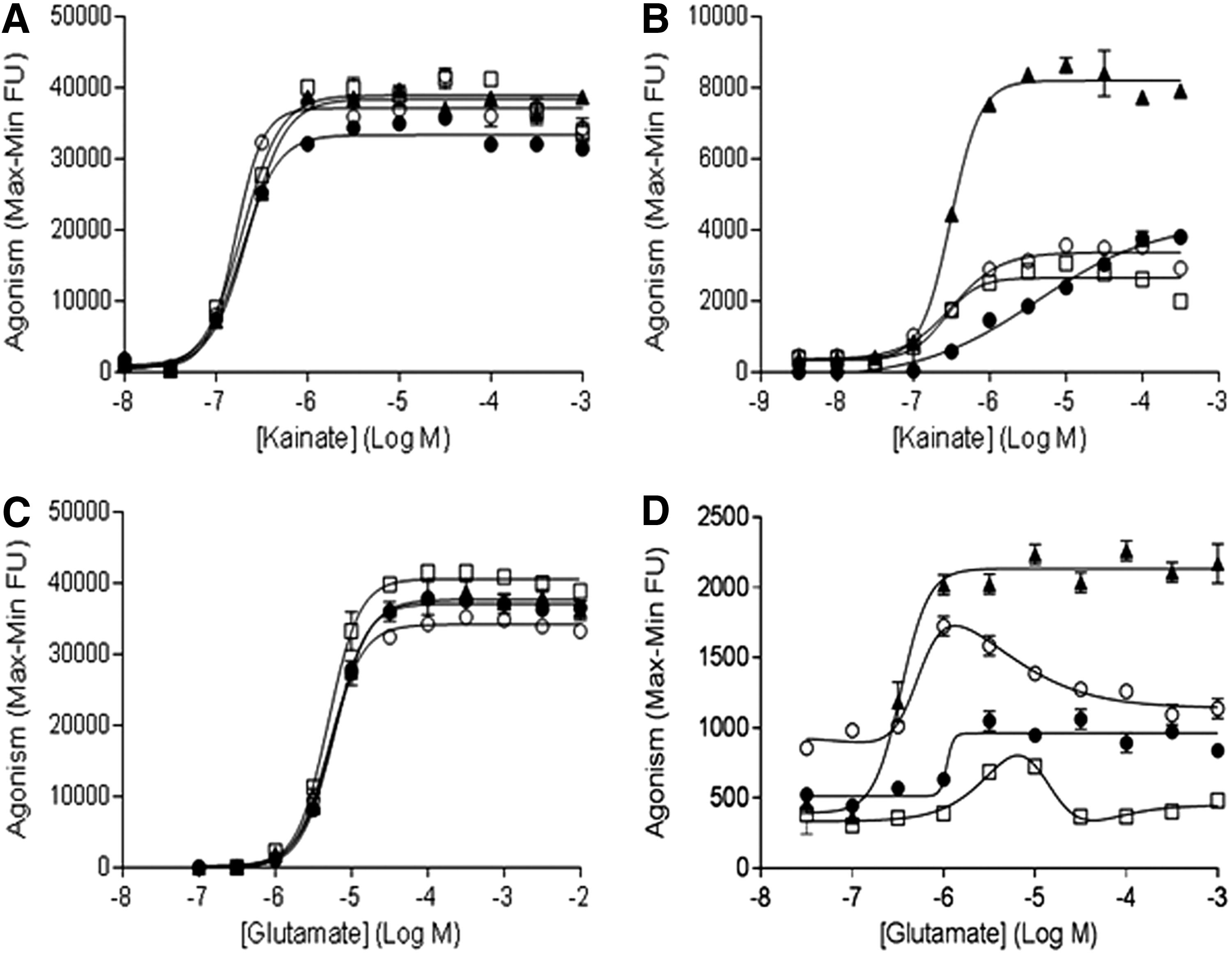
Results from FLIPR assays to measure modulatory effects of PSD95 and PSD95/NETO2 on kainate- or glutamate-induced responses in the presence or absence of Con A in cell lines expressing GluK2Q alone (●), GluK2Q/PSD95 (□), GluK2Q/NETO2 (○), and GluK2Q/PSD95/NETO2 (▲).
In the presence of Con A, neither PSD95 nor NETO2 had significant potentiating effect on glutamate-evoked GluK2Q-mediated FLIPR signals (Fig. 6C). Under these conditions, the potency of glutamate was similar in cells expressing GluK2Q alone, GluK2Q/PSD95, GluK2Q/NETO2, or PSD95/NETO2. In the absence of Con A, PSD95 alone did not potentiate the effect of glutamate on GluK2Q. Yet, coexpression of PSD95/NETO2 with GluK2Q had notable potentiating effects on glutamate-evoked GluK2Q-mediated FLIPR signals (Fig. 6D) compared to PSD95 alone or NETO2 alone.
Furthermore, compared to GluK1Q, the potentiating effects of PSD95/NETO2 on GluK2Q did not yield a “bell-shaped” curve and showed significantly less FLIPR signals (Fig. 6D). Nevertheless, compared to previous FLIPR assays with cells expressing GluK2 alone, which required Con A, utilization of FLIPR assays with cells expressing the complex of GluK2Q/PSD95/NETO2 represents a potential high-throughput assay to characterize the effects of glutamate at GluK2 in the absence of Con A.
Discussion
The discoveries of compounds that modulate the effects of glutamate at KARs are important drug targets for the potential treatment of various CNS disorders. 36,37,41,52 –55 Commonly used high-throughput FLIPR assays utilize recombinant cell lines expressing GluK ion channels only. 56 However, KARs do not appear to function as a single protein in native neuronal cells. 8,27 It has been recognized that recombinant and native KARs often differ in their pharmacological and biophysical properties. 5,8,57
These differences suggest that cell lines with heterologously expressed receptors may lack a potential modulatory component that could influence key characteristics of response at KARs. 8 In addition, artificial potentiators (e.g., Con A) are required to enable high-throughput FLIPR assays for GluK1 and GluK2 because of the fast desensitization after glutamate stimulation. Con A, which is not endogenously present in humans, is a commonly used potentiator in GluK1 and GluK2 FLIPR assays. The utilization of Con A in assays that characterize GluK1 or GluK2 effects in a screening paradigm may not be optimal.
To date, the utilization of native human cells is not logistically possible for high-throughput assays. To address this shortcoming, it would be preferable to conduct high-throughput GluK1 or GluK2 assays with recombinant systems that mimic native cells, where not only GluK1 or GluK2 but also their modulatory proteins are coexpressed. Furthermore, the ideal high-throughput GluK1 and GluK2 assays should be conducted in the absence of exogenous potentiators. In the attempt to address these concerns, we hypothesized that distinct combinations of NETO1, NETO2, and PSD95 with GluK1 or GluK2 would have distinct effects on GluK1 or GluK2 pharmacological activities or roles in synaptic plasticity.
In this study, we have developed a physiologically relevant FLIPR assay system with recombinant cells expressing combinations of GluK1Q, its modulatory subunits and PSD95 highly expressed scaffold protein, in the postsynaptic membrane of neurons. Compared to previous FLIPR assays using recombinant cells expressing only GluK1Q, coexpression of GluK1Q/PSD95/NETO2 may better mimic native function at GluK1. More importantly, this combination enabled robust glutamate-evoked GluK1-mediated FLIPR signals without the necessity of Con A. The peak FLIPR signals were achieved with 30 μM glutamate, while 100 μM glutamate (EC80) was used as an agonist in previous GluK1 FLIPR assays with cells expressing GluK1 only. Apparently, coexpression of PSD95/NETO2 with GluK1 increases glutamate's potency on GluK1.
We have previously found that the glutamate concentration utilized in compound screening assays could be critical in finding potent GluK1 antagonist compounds. In this study, lower levels of potentiation of glutamate effects by NETO1 or NETO2 alone on GluK1 were observed in the absence of Con A. We speculate that a stable cell line coexpressing GluK1/PSD95/NETO1 may be valuable to examine the glutamate-dependent calcium flux in the absence of Con A.
In this study, we also found interesting pharmacological properties in cells with different expression combinations of GluK2 with NETO1, NETO2, PSD95, or PSD95/NETO2. For example, cells coexpressing GluK2/PSD95/NETO2 respond to glutamate stimulation with appropriate signal levels in the FLIPR assay in the absence of Con A. Thus, it seems that the combination of PSD95/NETO2 is advantageous to modulate GluK1- and GluK2- mediated FLIPR signals.
In a previous study, it was demonstrated that coexpression of either NETO1 or NETO2 with GluK2Q did not alter the voltage-gated calcium permeability in the steady state of GluK2Q maintained by a weakly desensitizing agonist (domoate). 34 Although assays (electrophysiological recording vs. FLIPR) were different, results from our studies suggest that NETO1 or NETO2 regulated the GluK1Q- or GluK2Q-mediated calcium influx magnitudes when evaluated under certain assay conditions (Figs. 2B–D and 4B). Furthermore, by using a natural KAR agonist (glutamate), the calcium permeability in cells coexpressing GluK1Q, NETO2, and PSD95 was robustly increased compared to cells expressing GluK1 alone (Fig. 5D). The increased calcium permeability in the assay shown in Figure 5D was a key GluK1 channel property allowing us to enable a native-mimicking GluK1 high-throughput compound screening assay.
It was reported previously that the NETO2 protein levels and surface expression are decreased in GluK2 knockout mice, indicating that NETO2 expression and trafficking are regulated by GluK2 in mice. 53 Similar to that observation, expression levels of NETO1 or NETO2 in the recombinant cells used in this study were also regulated by GluK1 or GluK2. GluK2 increased NETO1 expression to a greater level than that observed with GluK1, while GluK1 increased NETO2 expression more than GluK2.
It is controversy in the literature if NETO1 or NETO2 alters the trafficking 25,32,34,58 of GluK2 to cell membranes to function. We observed that the cells coexpressing NETO2 and GluK2Q were more sensitive to glutamate (∼30 nM) compared to cells expressing GluK2 alone (Fig. 4D). Although our result was confirmed with repeated experiments, the observation could be just GluK2/NETO2 FLIPR assay specific. If not, it could indicate that NETO2 may modulate GluK2 channel properties (e.g., increased sensitivity to glutamate or autoactivation) through altering the expression or trafficking of GluK2.
It was reported that neuronal AMPA receptors and recombinant AMPA receptors coexpressed with their auxiliary protein (stargazin) autoinactivate at high concentrations of glutamate with bell-shaped response curves, whereas recombinant AMPA receptors expressed alone do not. 59 We observed similar glutamate response curves in some FLIPR assay conditions in our studies. In the absence of Con A, bell-shaped responses to glutamate stimulation were observed in cells expressing GluK1Q/NETO1, GluK1Q/NETO2, GluK1Q/PSD95/NETO2, GluK2Q/NETO1, and GluK2Q/NETO2, whereas not in cells expressing GluK1 or GluK2 alone.
Bell-shaped CRC were also observed for steady-state currents of native KARs and explained by the different concentration dependence of activation and desensitization. 60 Based on the study of stargazin-mediated bell-shaped glutamate response curves of AMPA receptors, 59 it is possible that the association and disassociation between NETO1 and NETO2 with a different state (resting or open) of KAR may result in the bell-shaped glutamate response patterns observed in our FLIPR assays.
It was reported that GluK2/Gluk5 complex binds to PSD95 tightly through the high affinity of GluK5 to the PDZ1 domain of PSD95. 42 Interestingly, we observed that PSD95 robustly enhanced bell-shaped glutamate response in the GluK1/NETO2 FLIPR assay, but abolished the bell-shaped glutamate response in the GluK2/NETO2 assay.
This difference suggests that PSD95 played different roles in modulating the two functional complexes of GluK1/NETO2 and GluK2/NETO2 in responding to glutamate stimulation probably due to its different binding affinities to the two complexes.
Although the underlying mechanism is unclear, these bell-shaped responses seem to be a common phenomenon in the above coexpression setting and may be indicative of a protective mechanism of native neurons to prevent excessive glutamate-induced firing in physiological and pathological states. As such, our findings indicate that the coexpression of GluK1Q/PSD95/NETO2 and GluK2Q/PSD95/NETO2 may better represent native states for GluK1 and GluK2 in recombinant systems for high-throughput assays.
In addition, regulating the kinetic properties of GluK1and GluK2 by targeting NETO1, NETO2, and PSD95, as well as their complexes, can increase the diversity of functional properties of GluK1 and GluK2. These modulatory subunits may represent a starting point for a better understanding of the role played by neuronal GluK1 or GluK2 in pathological conditions. Moreover, exploiting the effective pharmacological agents associated with specific GluK1 and GluK2 complex and their differential expression patterns may provide a suitable platform to develop better and safer therapeutics.
In this report, we present GluK1 and GluK2 FLIPR assays with various subunit compositions of GluK1, GluK2, NETO1, NETO2, and PSD95. For enablement of a FLIPR assay, the signal magnitude and the signal/noise ratio are two key factors considered to determine if an assay can be reliably used to screen compounds. No absolute standard requirements for these two factors were listed in our KAR compound screening practices, but general minimum requirements are suggested to avoid data variation and reliability concerns.
Based on our experience running KAR compound screening FLIPR assays, at signal test step, we optimize assay parameters to yield basal signals between 8,000 and 10,000 fluorescence counts (FU) and then look for ≥16,000–20,000 FU peak signals from EC80 of agonist stimulation. With these general requirements, the assays of GluK1/NETO1, GluK1/NETO2, and GluK1/PSD95/NETO2 are three reliable GluK1 high-throughput assays with either kainate or glutamate as an agonist without the need of Con A.
Of these three assays, the GluK1/PSD95/NETO2 is the best GluK1 high-throughput assay discovered in this report because it uses glutamate as an agonist without the need of Con A. More importantly, the GluK1 receptor properties in cells coexpressing GluK1/PSD95/NETO2 is more close to that observed in native cells such as bell-shaped glutamate response patterns 60 and relatively slowed desensitization. 30,32,58
The limitation of GluK1/NETO1 and GluK1/NETO2 assays is that they need to use kainate as an agonist and kainate does not exist in man. The GluK2/PSD95/NETO2 assay is the only suboptimal GluK2 compound screening assay discovered in this report. It does not need Con A, but still needs kainate as an agonist. Other assays presented in this report are not recommended as GluK1 and GluK2 compound screening assays, although some of them had notable levels of FLIPR signals and signal/noise ratios in the absence of Con A. Further studies are needed to determine if other KAR interaction proteins will help to deliver more reliable GluK1 and GluK2 high-throughput compound screening assays better mimicking the native channel properties of GluK1 and GluK2.
Footnotes
Acknowledgments
The authors would like to thank Dr. Andrew H Ahn for his critical comments on this article and Dr. Michael P. Johnson for his assistance on editing on bell-shaped curves.
Disclosure Statement
All authors are employees of Eli Lilly and Company.