
Abstract
The lymphatic system is a series of vessels that transport cells and excess fluid from tissues to the blood vascular system. Normally quiescent, the lymphatics can grow or remodel in response to developmental, immunological, or pathological stimuli. Lymphatic vessels comprise lymphatic endothelial cells (LECs) that can respond to external growth factors by undergoing proliferation, migration, adhesion, and tube and lumen formation into new vessel structures, a process known as lymphangiogenesis. To understand the key gene and signaling pathways necessary for lymphangiogenesis and lymphatic vessel remodeling, we have developed a three-dimensional LEC tube formation assay to explore the role of kinase signaling in these processes. The collagen-overlay-based assay was used with primary human adult dermal LECs to investigate a library of 60 tyrosine kinase (TK) and TK-like genes by siRNA-mediated knockdown. Nine candidate genes were identified and characterized for their ability to modify key parameters of lymphatic tube formation, including tube length, area, thickness, branching, and number of blind-ended sacs. Four genes—ZAP70, IRAK4, RIPK1, and RIPK2—were identified as high-confidence hits after tertiary deconvolution screens and demonstrate the utility of the assay to define LEC genes critical for the formation of tube structures. This assay facilitates the identification of potential molecular targets for novel drugs designed to modulate the remodeling of lymphatics that is important for the metastatic spread of cancer and other pathologies.
Introduction
The lymphatic vascular system plays important roles in maintaining tissue fluid balance, absorption of lipids, and immune cell surveillance. 1 Dysfunction of lymphatic vessels is associated with various pathologies, including inflammation, soft tissue swelling caused by accumulation of lymph in the interstitial tissue (lymphedema), and tumor metastasis. 1,2 The growth of lymphatic vessels (lymphangiogenesis) or their remodeling through dilation or hyperplasia is driven through regulated cellular processes such as cell migration, proliferation, adhesion, apoptosis, differentiation, and tube formation, which coordinate the formation of a distinct and regular pattern of lymphatic vessels that is necessary for a fully functioning lymphatic system. 2
Recent focus on the growth (angiogenesis) and remodeling of blood vessels has identified key pathways for the growth, development, and differentiation of blood vascular endothelial cells. 3,4 This understanding has led to the identification of a number of molecular targets for drugs that combat pathological angiogenesis in cancer and eye diseases. A key example is the vascular endothelial growth factor (VEGF) and vascular endothelial growth factor receptor (VEGFR) pathways that control angiogenesis in development and disease. 5
The lymphatic vessels have remained somewhat poorly characterized relative to blood vessels, initially due to the lack of specific molecular markers, the difficultly in distinguishing between blood and lymphatic vessels in histological sections, and the acute importance of the blood vascular circulation. 6 More recent attention on lymphatic vessels has shown their specialized development and the unique role they play in a number of human physiological and pathological processes such as tumor cell dissemination to distant organ sites and lymphedema. Understanding the molecular and cellular bases for these roles is important for identifying ways to target the lymphatics therapeutically. The related process of angiogenesis and blood vascular remodeling may be, in part, controlled by similar molecules and signaling pathways. 1,2,7 * This is important when considering the effects that antiangiogenic therapy, or other drugs directed to the blood vasculature, may have on the lymphatics. Furthermore, therapies directed at molecules controlling the immune system may affect the lymphatic endothelium, which has been shown to play a role in shaping the immune response against some antigens. 8 –12
Vessel formation is a crucial process in lymphangiogenesis. The ability of lymphatic endothelial cells (LECs) to form capillary-like tubes in in vitro models is a key step toward the generation of new lymphatic vessels. The tube formation assay was first demonstrated more than 20 years ago when blood vessel endothelial cells (EC) seeded on a reconstituted extracellular matrix (ECM) were found to form a meshwork of capillary-like structures with tight cell–cell junctions and intercellular or lumen-like spaces as demonstrated by microscopy. 13 During invasion and tube formation, ECs undergo several specific processes such as degradation of their basement membrane and adhesion and migration into the surrounding collagen-rich matrix, which recapitulates multiple steps involved in lymphatic vessel development. Many studies have utilized in vitro LEC tube formation assays to investigate lymphangiogenic properties of different factors. 14,15
Here, we present a study which examines a key group of signaling genes and the role they play in lymphatic endothelial tube formation, an important component of lymphangiogenesis and lymphatic vessel remodeling. Using a functional assay to model tube formation, we have shown that specific kinase-related genes modify key morphological parameters of lymphatic vessel patterning. The screen demonstrates a method to analyze and quantify effects on LEC tube formation and provides a framework for a genome-wide screen to facilitate drug discovery and design for diseases such as cancer and lymphedema.
Materials and Methods
Cell Lines and Cell Culture
Adult human LECs were derived from primary human adult dermal LECs (HMVEC-dLyAd, Lonza, Walkersville, MD), while human blood endothelial cells (BECs) were obtained from pooled primary human umbilical vein endothelial cells (HUVECs) (Cascade Biologics, Portland, OR). Both cell types were maintained in EGM™-2MV complete medium plus supplements as recommended by the manufacturer (Lonza). LECs were cultured on a substrate of human fibronectin (5 μg/ml, BD Biosciences, San Jose, CA). Cells were enzymatically dissociated from tissue culture plates using 0.25% Trypsin-EDTA (Thermo Fisher Scientific, Waltham, MA) and replated at 1:3–1:5 for routine passaging, or at the required density for experiments.
Transfection Optimization
Different transfection reagents were tested for their ability to successfully introduce siRNA into LECs. DharmaFECT® was compared to an amine-based transfection reagent in an Alamar Blue reduction assay (Thermo Fisher Scientific), to assess cellular viability through quantification of the cells' metabolic activity. The dye was added to the cells and fluorescence (excitation 530 nm, emission 590 nm) measured 3–4 h later.
To confirm efficient uptake of siRNA, LECs were transfected with different concentrations of a control Alexa Fluor® 546-labeled siRNA and subsequently analyzed by brightfield and fluorescence microscopy and flow cytometry. For flow cytometry, cells from each condition were trypsinized as above, collected by centrifugation and washed twice in phosphate-buffered saline (PBS) with 0.5% BSA and 1 mM EDTA. Cell samples were then resuspended in 200 μl of the same buffer supplemented with 0.4 μg/ml propidium iodide to label and exclude dead cells, and analyzed by flow cytometry (FACSCalibur, BD Biosciences). Flow cytometry data were analyzed using CellQuest Pro software (BD Biosciences).
Reverse Transfection of EC with Pooled and Deconvoluted siRNAs
All pooled (siGENOME SMARTpool) and deconvoluted siRNAs (a set of four siGENOME siRNAs) and non-targeting siRNA (siNT; ON-TARGETplus) were purchased from Dharmacon (Thermo Fisher Scientific). The siRNA sequences are shown in Supplementary Table S1 (Supplementary Data are available online at
Collagen Overlay Assay
Cells were prepared for the collagen assay by preplating cells at 104 cells/well in a 96-well plate (with or without siRNA) and incubating at 37°C and 5% (v/v) CO2 for 72 h. To prepare a 10 mL overlay collagen mix (1 mg/mL collagen), 50 μL 1 N sodium hydroxide (Sigma-Aldrich), 333 μL of 10× PBS (Invitrogen, Thermo Fisher Scientific), 950 μL nuclease-free water (Ambion, Thermo Fisher Scientific), 2 mL of bovine collagen I (5 mg/mL) (Gibco), and 6,670 μL EGM-2MV complete medium were mixed on ice. One hundred microliters of the collagen mix was carefully overlaid on top of the cell monolayer to avoid bubble formation. The plates were incubated at 37°C and 5% (v/v) CO2 and the collagen was left for 15–30 min to polymerize and form a gel. Cells were incubated for 8 h, after which they were fixed by aliquoting 100 μL of freshly prepared 4% paraformaldehyde (PFA) in PBS onto each well and left on a shaker overnight at RT.
Total RNA Extraction
For RNA extraction from cell monolayers, the RNeasy® Micro Kit (Qiagen, Hilden, Germany) was used to isolate RNA following manufacturer's instructions. For RNA extraction from collagen gels in 96-well plate, TRIzol® reagent (Ambion) and RNeasy Mini Kit (Qiagen) were used with modification. Briefly, 600 μL TRIzol was added to every 10 wells × 100 μL collagen gels and transferred into a 2 mL Eppendorf tube. The sample was vortexed for 20 s and allowed to sit for 5 min at RT. Four hundred microliters of chloroform was added to each sample and the tube was shaken vigorously for 15 s and let to sit at RT for 2 min before centrifuging the samples at 12,000 g for 15 min, at 4°C. The upper aqueous phase was transferred to a new 1.5 mL Eppendorf tube filled with 500 μL 70% ethanol, and mixed by pipetting. Afterward, 700 μL of the ethanol-RNA solution was transferred into the RNeasy Mini Spin column and centrifuged at 8,000 g for 20 s, at 4°C. The supernatant was discarded and this process was repeated for the remaining ethanol-RNA solution from two samples (total from 20 wells × 100 μL collagen gel). The samples then underwent a series of washing steps following the manufacturer's instructions, and the RNA was resuspended in 30 μL RNAse-free water. RNA concentration and purity were assessed using Nanodrop 2000 (Thermo Scientific).
Quantitative Reverse Transcription Polymerase Chain Reaction
Total RNA was used to generate cDNA using the High Capacity cDNA Reverse Transcription Kit with RNase inhibitor (Applied Biosystems, Foster City, CA) following the manufacturer's instructions. Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed using the StepOnePlus™ 96-well Real-Time PCR system to quantify RNA transcripts of the target gene. The PCR was prepared using TaqMan® Fast Universal PCR Master Mix (2 × ), No Amperase® UNG (Applied Biosystems), and TaqMan Gene Expression Assays (Applied Biosystems) as per manufacturer's instruction. Relative expression was normalized to ACTB (β-actin).
Intracellular Staining of Collagen Cultures
After overnight fixation, the fixative solution was drained from the collagen gels by inversion of the plates onto paper towels, and 100 μL of permeabilization solution [0.2% (v/v) Triton X-100, 1% bovine serum albumin in PBS] was added into each well. The plates were covered with aluminum foil to protect from light and left on a shaker for 3 h at RT. Collagen gels were stained with 100 μL of staining solution prepared from Hoechst 33342 dye (1:500 dilution) and Phalloidin Alexa Fluor 488 (1:100 dilution) in PBS for 3 h. Wells were washed with PBS twice before imaging.
Imaging and Analysis
The plates were heat-sealed and imaged using Cellomics® (Thermo Scientific) with ArrayScan ® VTI HCS Reader software or on some occasions, a BD pathway 435 (BD Biosciences). At 5 × objective, nine fields (each with 1.3 × 1.3 mm field of view) were imaged per well and were stitched together to create a montage. Images were processed using advanced digital imaging software MetaMorph™ (version 7.7.8, Molecular Devices, San Diego, CA) as follows: the tubes were analyzed using the in-built Tube formation assay module to attain the following parameters: length and breadth of tubes; number of tubes; number of nodes; number of branches; and number of connected sets. The output mask from the Tube formation assay module was then used for further processing as follows: loops (the area between tubes) were extracted and their area measured. To qualify as a loop, the extracted area had to have a shape factor of greater than 0.45 (1 equals a perfect circle). The node and tube parts of the mask were used to extract the nuclei from each of their respective classification areas and these were then counted. This provided a read out of proliferation versus growth—many small cells or fewer large cells. Data were expressed as a fraction normalized to the negative control. Cell count and viability were obtained using Cellomics vHCS View ver. 1.6.2. Some images were autocontrast adjusted using Adobe Photoshop CS4 for presentation.
Statistical Analysis
Each well was normalized to the mean of the mock-transfected control. The average of technical replicates was taken and z-scores were calculated and identified using robust z-score >|2| and binned as “enhancing” tube parameters (z-score >2) or “impairing” tube parameters (z-score < −2). Data were analyzed by Student's t-test. P values were significant at P < 0.05 using PRISM software (GraphPad Software, Inc., San Diego, CA). One-way ANOVA with Dunnett's post hoc test was used to compare expression levels for the time course experiment.
Results
In vitro LEC Tube Formation Assay Establishment
An in vitro tube formation assay that relied on type I collagen as ECM, to allow monolayer-seeded LECs to invade the ECM and form tube- or vessel-like structures, 16 was developed using primary human adult microvascular LECs. This is a surrogate assay to identify genes involved in the formation of tube structures seen in lymphangiogenesis and lymphatic vessel remodeling, in a high-throughput format (Table 1). Substantial experiments were performed to set up the assay and validate the approach. This included optimization of the transfection reagents and input siRNA concentration (Supplementary Fig. S1), confirmation of siRNA delivery, cell viability, and selection of control siRNA (Supplementary Figs. S2–S4), assessing the dynamic range of the assay and the quantification of tube formation by a unique image analysis protocol (Supplementary Fig. S5).
Three-Dimensional Lymphatic Endothelial Cell Tube Formation Assay Protocol
1. Different primary endothelial cells obtained from commercial suppliers and passaged in EGM™-2MV complete medium (Lonza) with growth supplements according to the manufacturer's instructions.
2. Cells are coated onto fibronectin-coated tissue culture plastic.
3. Aspiration of media performed by multichannel Finnpipette.
4. A 10 mL collagen overlay solution is prepared for each 96-well plate on ice and should have a pinkish hue. This is then layered on the top of the cell monolayer.
5. Incubations can vary depending on the style of experiment. Eight hours for a standard experiment, but time courses could recover samples every 2 h (e.g., assay completed at 2, 4, 6, and 8 h).
6. Plates are left on the shaker overnight with 100 μl of fixative.
7. Used TRIzol® based RNAeasy micro kit (Qiagen).
8. Used high-capacity cDNA Reverse Transcription Kit with RNase inhibitor according to the manufacturer's instructions (Applied Biosystems).
9. qRT-PCR performed using StepOnePlus™ 96-well real-time PCR system.
10. Fixed in 0.2% Triton X-100, 1% bovine serum albumin/PBS with Hoechst dye or Phalloidin Alexa Fluor 488.
11. Images taken with Cellomics ArrayScan® (Thermo Scientific).
BEC, blood endothelial cell; LEC, lymphatic endothelial cell; PBS phosphate-buffered saline; PFA, paraformaldehyde; qRT-PCR, quantitative reverse transcription polymerase chain reaction.
A Primary siRNA Screen of LECs During In Vitro Lymphangiogenesis Reveals the Involvement of Novel Kinase Genes
In search of potential new therapeutic targets for lymphatic vessels during disease states such as cancer metastasis by lymphatics and lymphedema, we focused on a class of tyrosine kinases (TK) and serine/threonine kinases (STK) known to mediate key cellular responses, including cell proliferation, migration, survival, and cell differentiation, some of which are known regulators of lymphangiogenesis. 17,18 As part of the experimental workflow, we interrogated a boutique library of pooled siRNAs targeting 60 TK/STK genes, including several receptor tyrosine kinases (RTKs), using our modified version of the collagen overlay assay to induce three-dimensional (3D) LEC tube formation. The resulting tubes were fluorescently stained to allow for their visualization, followed by high-content imaging and analysis, which revealed morphological differences in the tube patterning of LECs transfected with siRNA compared with the nontargeting control siRNA when overlaid with the collagen gel (Fig. 1A). We have previously developed a computer-based system to assess lymphatic vessel patterning in situ based on several key morphological and patterning characteristics that take into account structurally important features of the lymphatic vasculature, such as intervessel distance, branching, blind-ended sacs, vessel loops, and vessel width and length that are fundamental to the proper functioning of the lymphatic system. 19 To quantify the knockdown effects of various TK/STK genes by the siRNA pools, we utilized the same parameters in vitro, generating a nonbinary readout (Fig. 1B, C). This added to the complexity of the screen, with the readout generating information about the specific role some TK/STKs may play in vessel formation such as fine-tuning branching as opposed to those that may regulate tube length (Fig. 1B, C).

Overview of the screening platform using the 3D collagen assay to identify novel genes important for lymphatic vessel formation.
To assign a gene as a positive hit in the primary screen, we choose those genes affecting two or more vessel parameters with a robust z-score of >|2|. Based on these selection criteria, primary screening identified nine candidate TK/STK genes (∼16% of total genes tested) that may play a role in LEC tube formation. The genes EPHA7, IRAK4, KIAA1804, MAP3K11, MERTK, MAP3K9, RIPK1, RIPK2, and ZAP70 were subjected to further analysis.
Gene Profiling of Candidate TK/STK Genes During In Vitro Lymphangiogenesis
To validate expression of the candidate TK/STK genes identified by our primary screening of LECs, we examined their relative expression by qPCR. As a baseline, we compared relative expression profiles of resting LECs that would be reminiscent of the quiescent state of the adult lymphatic vasculature. All nine candidate genes were expressed at variable levels during the resting state (Fig. 2A). RIPK2 showed highest level of expression compared to IRAK4, MAP3K11, RIPK1, and MERTK, while ZAP70, MAP3K9, KIAA1804, and EPHA7 had lower expression levels (Fig. 2A). To determine whether these genes are specifically expressed by LECs and therefore exclusive to lymphangiogenesis, we compared levels to those in BECs. While all of the candidate TK/STK genes were expressed by BECs at various levels, expression was more prevalent in LECs, especially RIPK2 (Fig. 2A).
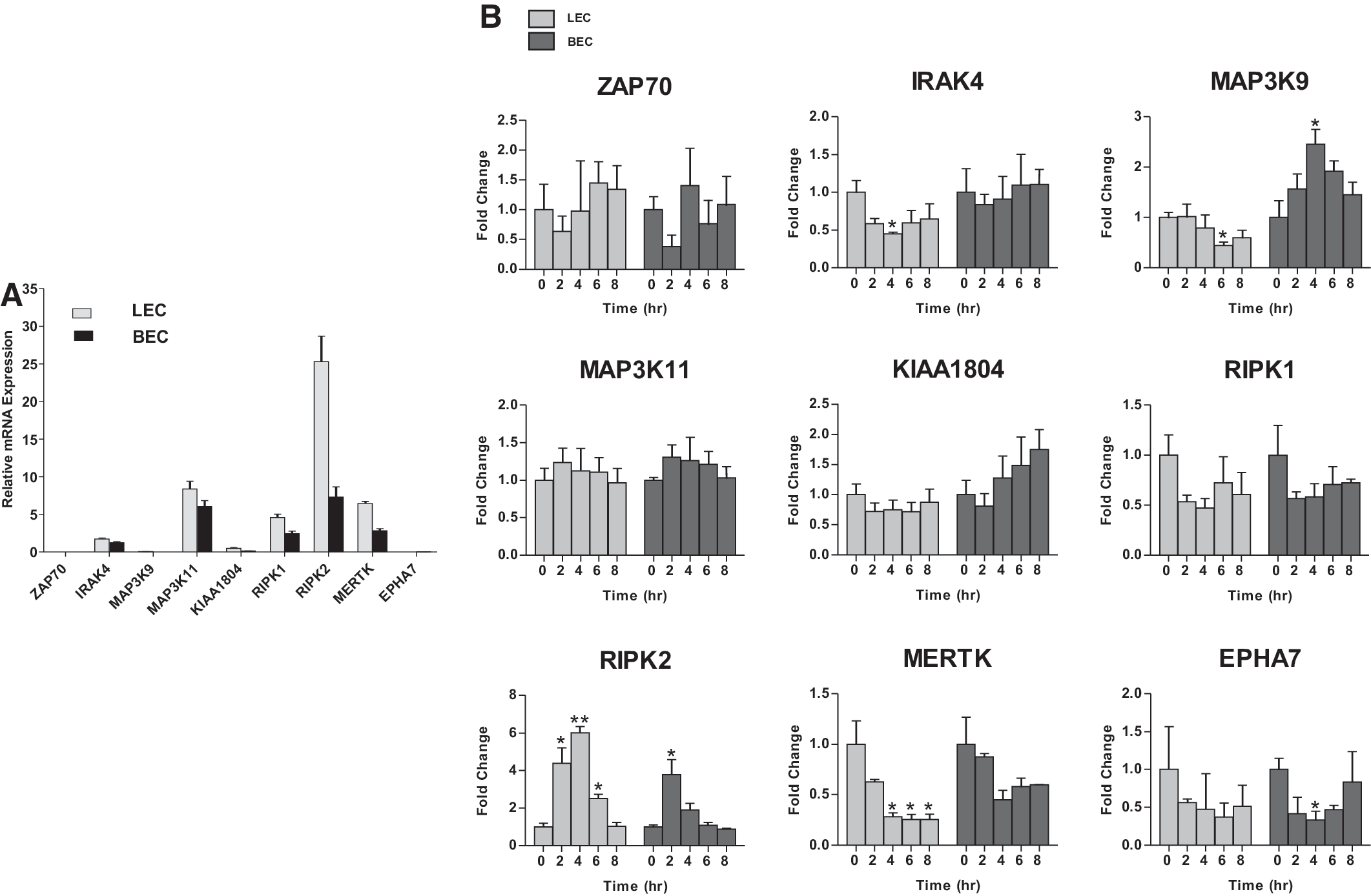
Expression of candidate genes in primary LECs during quiescence and in vitro lymphangiogenesis.
We next assessed expression of the candidate TK/STK genes in LECs by developing an RNA extraction method from the collagen gels that would allow the analysis of genes during the course of in vitro lymphangiogenesis and lymphatic vessel remodeling. In addition, we applied the collagen overlay assay to BECs to assess the applicability of this assay to other related ECs. qRT-PCR was performed at various intervals during the course of the collagen overlay assay on both LECs and BECs. Interestingly, while most of the TK/STK genes remained consistently expressed during the course of our in vitro lymphangiogenesis assay, there was a pattern of differential expression of some of the genes. For example, RIPK2 expression was significantly upregulated in LECs undergoing tube formation, peaking at 4 h before returning to baseline, whereas IRAK and MERTK expression were downregulated over the course of the assay that was not evident in BECs undergoing equivalent in vitro angiogenesis (Fig. 2B). Overall, these data suggest that the candidate TK/STK genes identified by the primary screen are expressed by ECs and may be important during active tube formation.
Validating Candidate TK/STK Genes in a Secondary Screening of LECs
We sought to validate these targets by performing a secondary RNAi screen in LECs using our collagen overlay assay. We utilized a pool of four different siRNAs (SMARTpool) targeting the same gene transcripts to ensure effective gene knockdown that was quantified by qRT-PCR. All nine genes showed significant knockdown efficiency of the target gene compared to the mock control in both LECs and BECs, with the exception of ZAP70 (Fig. 3A, B), which may have been attributed to very low expression levels, making accurate knockdown efficiency measurement difficult.
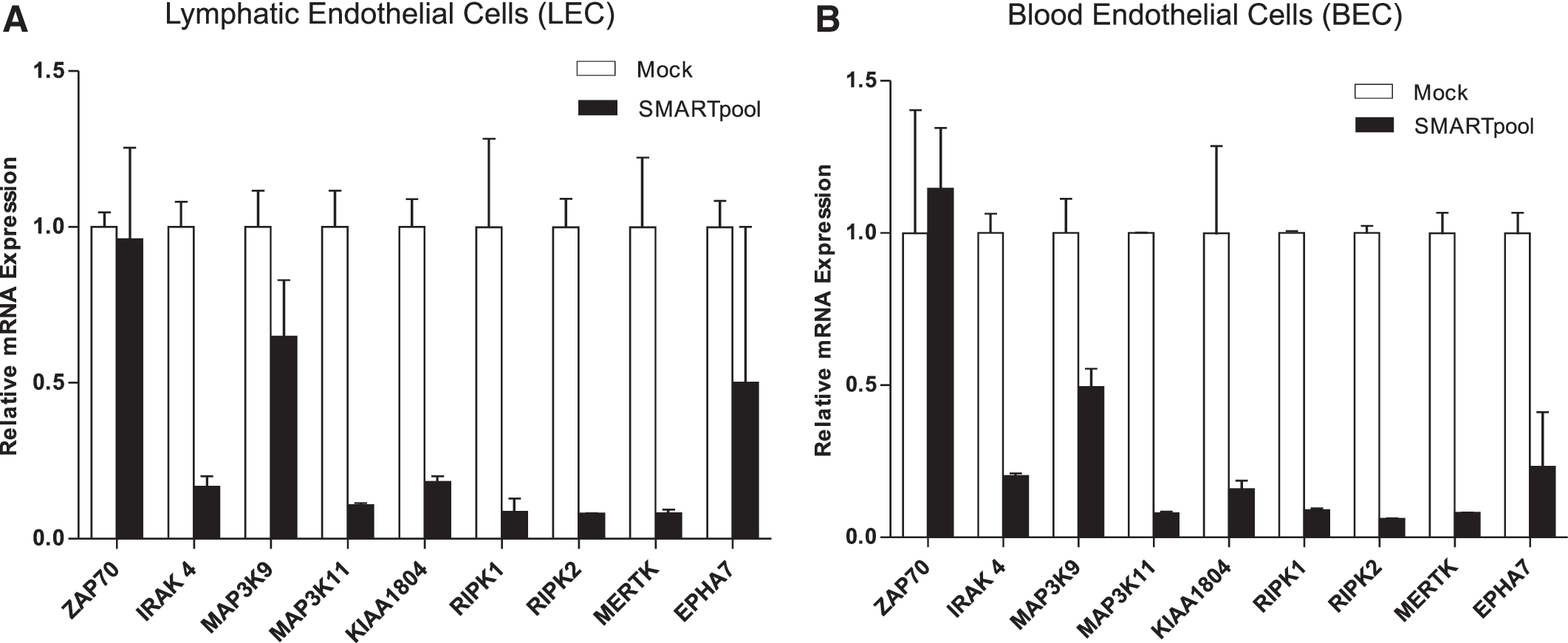
Secondary screening validation in LECs. LECs
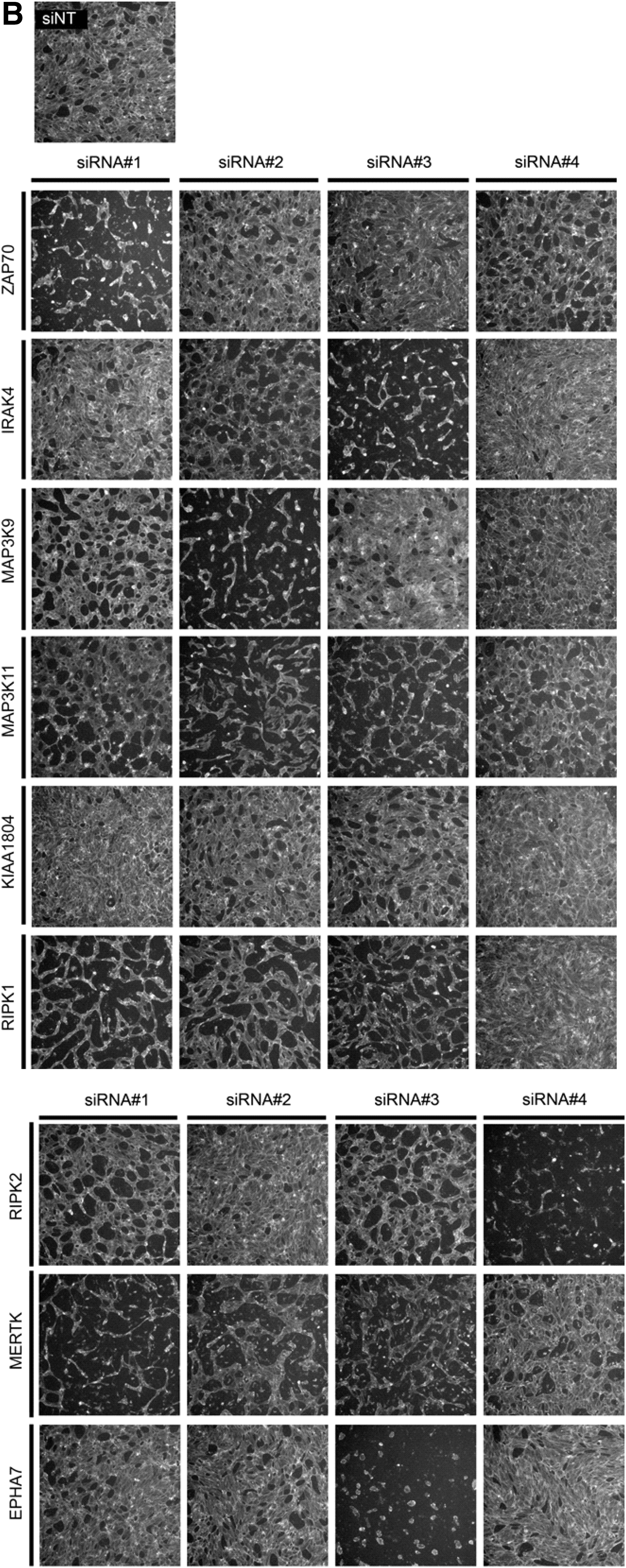
To assess the effects of knockdown of these candidate genes on the ability of LECs to form tubules, we performed collagen overlay assays. Morphological inspection of tubule formation following knockdown of ZAP70, IRAK4, MAP3K9, MAP3K11, KIAA1804, RIPK1, RIPK2, MERTK, and EPHA7 revealed differences in tubule patterning compared to the mock controls (Fig. 4A). Morphological differences were observed in LEC tubules compared to those formed by BECs (Fig. 4B) suggesting that these genes may regulate tube formation differently in LECs and BECs.
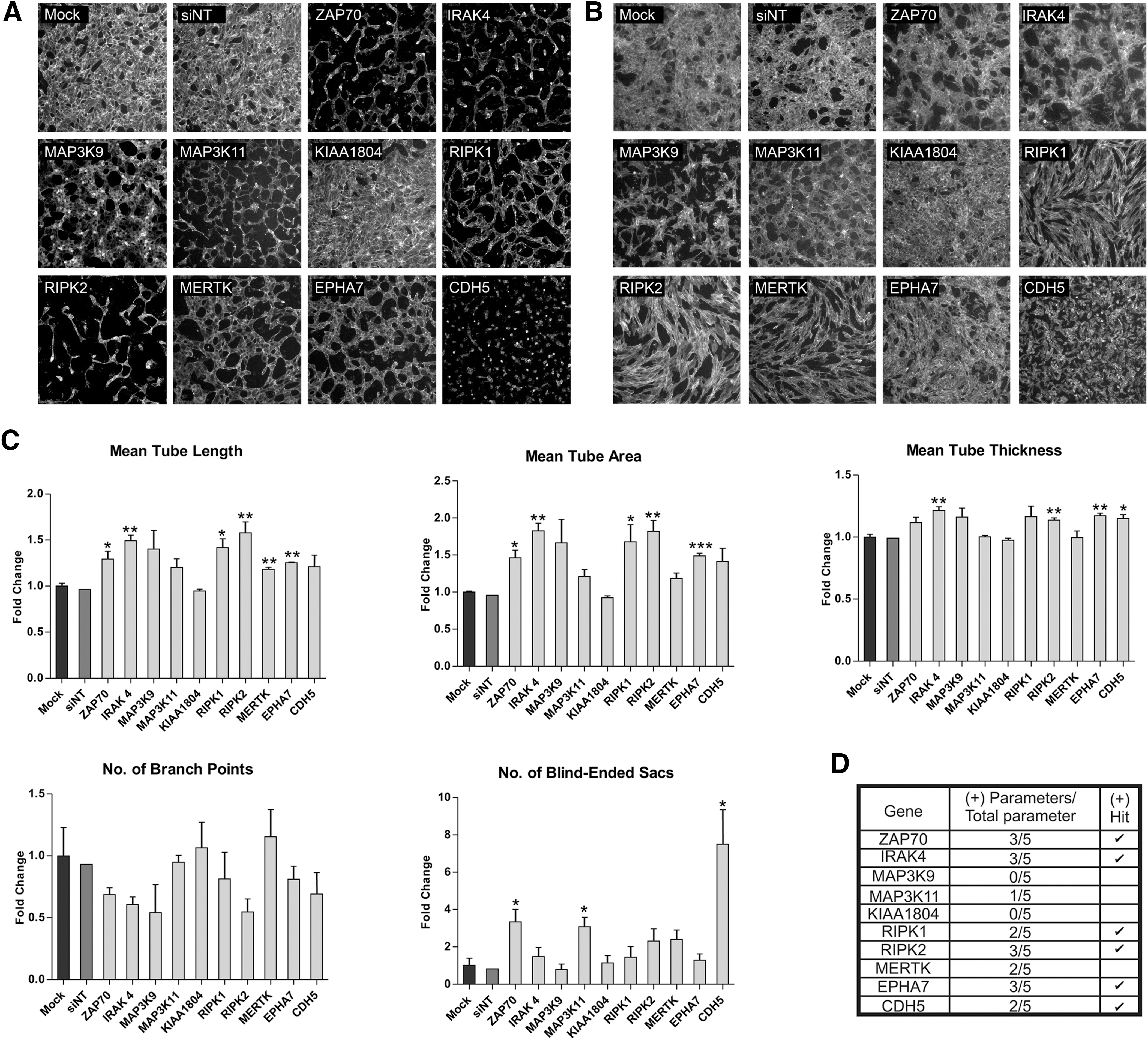
Quantification of tubule morphology following secondary screening of LECs. Collagen overlay assay was performed post-SMARTpool transfection and cells were visualized by Phalloidin staining. Images were captured using the Cellomics ArrayScan at 5× magnification for
To assess patterning, we evaluated vessel parameters including the following: (1) mean tube length, (2) mean tube area, (3) mean tube thickness, (4) number of branch points, and (5) number of blind-ended sacs. We showed that knockdown of the nine candidate genes affected different parameters of tube formation (Fig. 4C). In the secondary screening platform, we defined a positive hit as a gene for which the SMARTpool significantly affected two or more tube parameters compared to the mock control. Based on these criteria, ZAP70, IRAK4, RIPK1, RIPK2, and EPHA7 were selected for tertiary screening and validation. Secondary screening illustrates the variable effects on tubule patterning when specific genes are knocked down, as well as illustrating patterning differences between LEC and BEC tubes during this process. The screening also confirms ZAP70, IRAK4, RIPK1, RIPK2, and EPHA7, derived from the primary screen, as the candidate genes that may be important during lymphangiogenesis (Fig. 4D).
Tertiary Screening of LECs During In Vitro Lymphangiogenesis Reveals ZAP70, IRAK4, RIPK1, and RIPK2 as High Confidence Hits
Although pooled siRNAs increase the probability of gene knockdown, siRNA pools often fail to display a phenotype consistent with the single deconvoluted siRNAs, as well as showing increased false positives or the possibility of false negatives. 20 To confirm that the effect on tube formation was specific to the knockdown, a tertiary screen using different numbers of siRNA duplexes was performed. qRT-PCR revealed knockdown efficiency between 60% and 90% as well as variation in LEC tubule patterning between deconvoluted siRNAs (Fig. 5A, B). The morphological differences observed following tertiary screening were quantified to identify high confidence hits, with those siRNAs significantly affecting >3/5 tubule parameters being considered to be positive (Figs. 5B and 6). From this analysis, genes that were affected by >3/4 deconvoluted siRNAs are considered high confidence hits, 2/4 siRNAs are considered medium confidence, and those affected with <1/4 siRNAs are categorized as low confidence hits. Of the nine TK/STK genes tested, four TK/STK genes were validated as high confidence hits (ZAP70, IRAK4, RIPK1, and RIPK2), three with medium confidence (MAP3K9, MAP3K11, and KIAA1804), and two with low confidence (MERTK and EPHA7) (Table 2). Therefore, the development of a 3D collagen overlay assay as a readout for high-throughput siRNA screening has revealed novel genes that may be important during the process of lymphangiogenesis and lymphatic vessel remodeling.
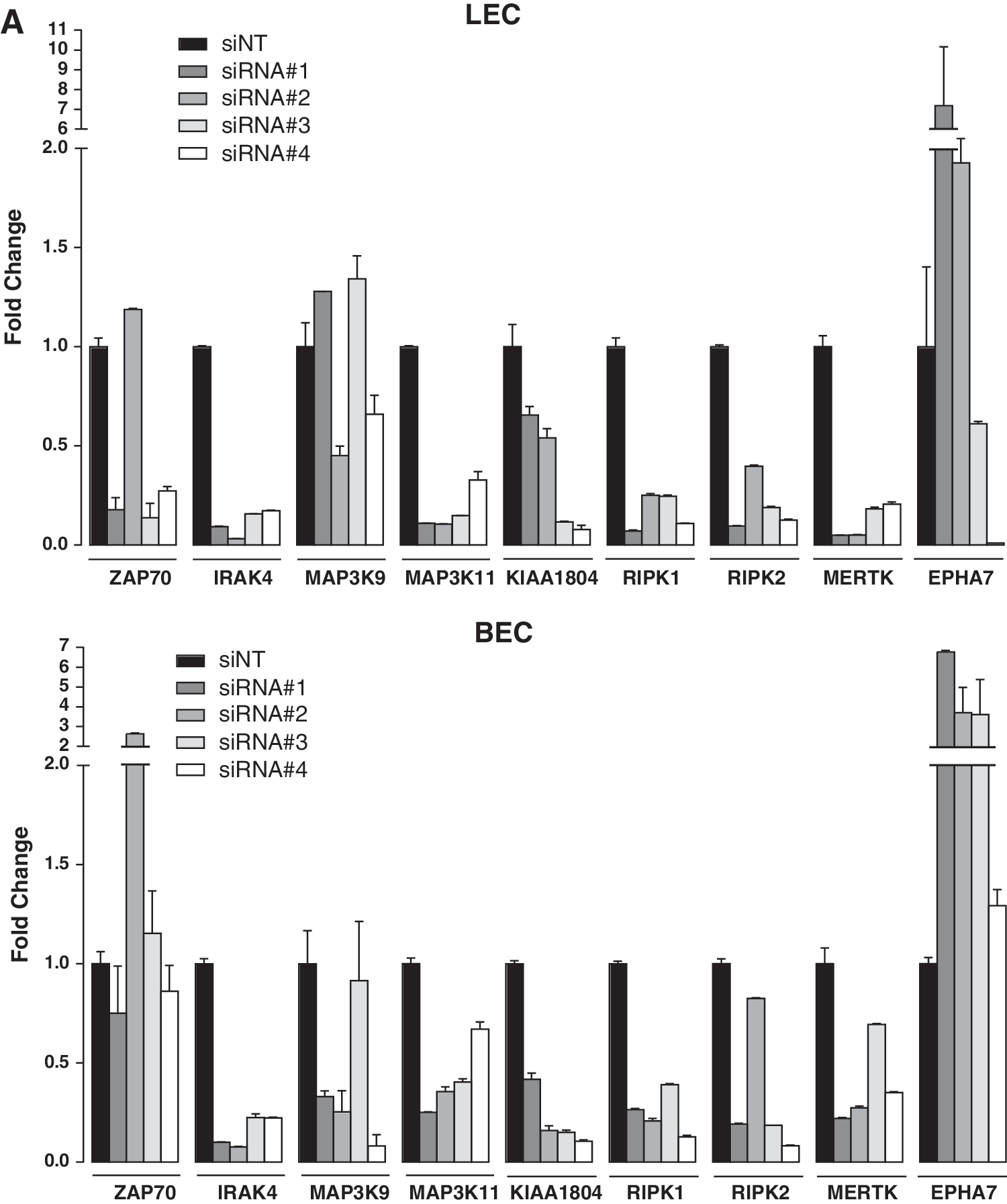
Tertiary screening of LECs.

Quantitation of tubule morphology following tertiary screening of LECs. The tube parameters quantified are as follows: mean tube length, mean tube area, mean tube thickness, number of branch points, and number of blind-ended sacs. Results are normalized to the siNT control and the data are presented as mean ± SD from three biological replicates (n = 3). Student's t-test was used to calculate the P value for each siRNA compared to the siNT control (*P < 0.05, **P < 0.01, ***P < 0.001). The gray shading indicates 2 SD below and above the mean of the siNT control that serves as the cutoff threshold. A statistically significant bar that falls below or above the gray shading was considered a positive hit. The positive control that affects four or more parameters was CDH5 (VE-Cadherin).
Summary of the Expression Analysis and siRNA Screening Using the Pooled and Deconvoluted siRNAs
Discussion
Lymphangiogenesis, the formation of new lymphatic vessels, is a multistep process involving the migration and proliferation of ECs, followed by tube formation and maturation to form a new functional vessel. 2,21 While lymphangiogenesis is believed to be rare in the resting adult, it can be activated during wound healing, inflammation, and cancer. In the context of cancer, targeted therapeutic inhibition of the key steps of lymphangiogenesis could provide a potential antimetastatic benefit. In search of potential new therapeutic targets, we developed a screen, which focused on a class of TK/STK, known to mediate key cellular responses, including cell proliferation, migration, survival, and cell differentiation in cell systems. We describe an assay system to detect genes that affect different parameters of lymphatic vessel morphology, such as vessel length, width, and branch points. The system demonstrated robust performance in two types of ECs and allowed us to identify potentially clinically useful hits from a pharmacologically targeted gene family.
Our primary RNAi screen was performed on a boutique TK/STK library of 60 genes to identify a subset important in LEC tube formation. We were able to compare the importance of these genes in BECs as a way to interrogate the fundamental differences between lymphatic and blood vessel formation. A previous study had identified genes that are differentially expressed between LECs and BECs. 22 It identified MAP3K9 to have a comparable expression between LECs and BECs, and RIPK to have selective expression in LECs, akin to what was observed in our study. Interestingly, our analysis of gene expression during the course of in vitro lymphangiogenesis revealed that RIPK2 is differentially expressed over time, supporting the notion that this candidate is important during the process of lymphatic vessel patterning. The screening also highlighted the characteristically different tube patterning between LECs and BECs, which may be a reflection of the differential or selective expression of such candidate genes. 23
Candidate genes from the primary screening were validated in a secondary screening using pooled siRNAs, and further subjected to tertiary screening using deconvoluted siRNA duplexes to identify high confidence hits and eliminate false positives. To address off-target effects, we set the threshold to a minimum of two siRNAs that yielded an overlapping phenotype for a gene to be considered a medium to high confidence hit. Since specificity is important in RNAi experiments, siRNAs with low off-target effects are preferable, such as the chemically modified ON-TARGETplus siRNA. 24 Phenotypic differences exhibited by some of the deconvoluted siRNAs may be attributed to off-target or toxic effects rather than a knockdown of the target gene itself. 25,26 Since an off-target effect can cause cell toxicity, 27 we monitored cell viability during the tertiary screening process and revealed cases where some siRNAs were toxic. For example, siRNA No. 3 targeting EPHA7 caused a <10% cell viability and was thus excluded from further analysis.
Perhaps the most extensively studied of the RTK family relating to lymphangiogenesis is VEGFR-3, which was the first growth factor receptor TK shown to be specifically expressed by lymphatic endothelium. 28 VEGFR-3 is the key receptor mediating lymphangiogenic signaling in embryonic development and the adult. 29 –33 VEGF-C and VEGF-D are the ligands for VEGFR-3 34,35 and signaling through this pathway has been shown to induce LEC migration, proliferation, and survival. 29 –31 Other characterized RTK signaling pathways that have a role in lymphangiogenesis are as follows: the Ang/Tie signaling pathway has been shown to be important during lymphatic vessel development 36 –38 ; fibroblast growth factor receptor (FGFR)-3 ligands FGF-1 and FGF-2 have been shown to promote proliferation, migration, and survival in cultured LECs, and stimulate lymphatic vessel growth in mouse cornea 39 –41 ; insulin-like growth factor (IGF)-1 and IGF-2 binding to IGF1R also induce lymphangiogenesis by stimulating proliferation and migration of primary LECs 42,43 ; and signaling by the STK activin-like kinase-1 (ALK1), a TGFβ type-I receptor, has also been found to promote both blood and lymphatic vessel development. 44
Our approach has shown the utility of using primary LECs in a collagen overlay assay to screen for additional RTK and other kinase genes associated with lymphangiogenesis. Using a siRNA platform, we have directly targeted TK/STK genes with functional significance during the process of in vitro lymphangiogenesis. This screening approach using a nonbinary readout of five key tubule parameters, such as tube length, thickness, branch points, density, and blind ending sacs, has identified novel TK/STK genes—ZAP70, IRAK4, MAP3K9, MAP3K11, KIAA1804, RIPK1, and RIPK2—as medium to high confidence hits implicated in lymphatic vessel formation and patterning. For most of these candidates, their exact involvement in lymphangiogenesis and lymphatic vessel development will need to be characterized by further studies. For example, ZAP70, along with the related spleen tyrosine kinase (Syk) was found to function redundantly to promote early angio- and lymphangioblast migration in zebrafish. 45 IRAK4 is associated with IL-1β-driven angiogenesis in inflammation, 46 and an in vitro study demonstrated that IRAK4 signaling is required for VEGF expression in macrophages. 47 MAP3K9 acts downstream of Rac1 and Cdc42, pathways which have been implicated in lymphangiogenesis. 48 Deletion of Rac1 in mice resulted in impaired lymphatic vessels and defective angiogenesis and, in vitro, RAC1 regulates tubulogenesis in HUVEC 49 and primary ECs. 50 It would be interesting to investigate whether MAP3K9, a medium confidence hit, can play a role in mediating these effects. MAP3K11 knockdown using short hairpin RNA (shRNA) was shown to inhibit tumor growth and metastasis of a breast cancer line in mice, 51 and the KIAA1804, RIPK1, and RIPK2 genes have been implicated in MAPK signaling in colorectal cancer and apoptosis, respectively, 52,53 yet their role(s) in lymphangiogenesis remains to be explored.
We observed differences in the tubule patterning depending on the specific gene being inhibited. This highlights the individual roles that genes play in a complex process such as tube formation and shows how different morphological changes to vessels that might occur in pathologies are driven from changes to gene expression/function. It also shows how different signaling pathways, through their connection with different cell surface molecules or internal protein structures, can control the interaction of ECs with other ECs and ECM.
This screen could be adapted to a 96-well format genome-wide scale using a randomized plate format that we have used previously for a genome-wide migration screen on LEC. 7 The challenges to overcome would include the application of the collagen overlay and timing correctly the dynamics of the tube formation for optimal evaluation of individual parameters.
This study has revealed a tool for identifying genes that may be important during lymphangiogenesis and lymphatic vessel remodeling. The assay has used a population of highly enriched LECs, defined by specific gene expression and cell surface markers. The method of successfully transfecting the primary LECs means that the direct effect of knocking down genes expressed by the LECs can be monitored in the assay. Previous studies have demonstrated the effect on in vitro lymphangiogenesis when specific genes are knocked down in other cell types (e.g., HEK, U87MG, and MDA-MB-231), which are then cocultured with primary endothelial cells. 14 Our study has further investigated a broad range of structurally related and druggable molecules, the TKs and STKs, to provide a basis for a larger style screen. In addition, this 3D LEC formation assay may be applied to other cell types, such as BECs and can be used to screen small molecule inhibitors and neutralizing antibodies. Positive hits can be further validated by a combination of biochemistry and mouse models to interrogate novel signaling pathways during the process of lymphatic vessel formation, therefore providing a new therapeutic discovery platform for lymphatic vessel disorders.
Footnotes
Acknowledgment
We wish to thank Jason Li, Peter MacCallum Cancer Center for help with bioinformatics and statistical analysis.
Authors' Contributions
All authors have contributed to the article and approved the final version.
Disclosure Statement
S.A.S. and M.G.A. are shareholders in Opthea Limited that develops therapies to vascular targets in eye disease and cancer. For all other authors, no competing financial interests exist.