
Abstract
Human glutamate dehydrogenase (GDH) plays an important role in neurological diseases, tumor metabolism, and hyperinsulinism–hyperammonemia syndrome (HHS). However, there are very few inhibitors known for human GDH. Recently, Ebselen was reported to crosslink with Escherichia coli GDH at the active site cysteine residue (Cys321), but the sequence alignment showed that the corresponding residue is Ala329 in human GDH. To investigate whether Ebselen could be an inhibitor for human GDH, we cloned and expressed an N-terminal His-tagged human GDH in E. coli. The recombinant human GDH enzyme showed expected properties such as adenosine diphosphate activation and nicotinamide adenine dinucleotide/nicotinamide adenine dinucleotide phosphate dual recognition. Further, we developed a 2-(3-(2-methoxy-4-nitrophenyl)-2-(4-nitrophenyl)-2H-tetrazol-3-ium-5-yl) benzenesulfonate sodium salt (EZMTT)-based assay for human GDH, which was highly sensitive and is suitable for high-throughput screening for potent GDH inhibitors. In addition, ForteBio binding assays demonstrated that Ebselen is a reversible active site inhibitor for human GDH. Since Ebselen is a multifunctional organoselenium compound in Phase III clinical trials for inflammation, an Ebselen-based GDH inhibitor might be valuable for future drug discovery for HHS patients.
Introduction
Human glutamate dehydrogenase (GDH) is a mitochondrial enzyme that catalyzes the reversible conversion of glutamate to α-ketoglutarate and ammonia by using NAD(P)+ or NAD+ as coenzyme. Glutamate is a major excitatory neurotransmitter and its concentration in the cerebral extracellular fluid has to be kept at low micromolar levels to stimulate rapid signal transduction when millimolar amounts of glutamate in vesicles are released from the neurons. The clearance of glutamate is dependent on efficient cellular uptake and rapid catabolism by several enzymes, including GDH. 1 Dis-regulation in glutamate catabolism, resulting in abnormally high levels of glutamate in the brain, causes neurological problems such as epilepsy. GDH plays an important role in glutaminolysis and in generating α-ketoglutarate to fuel the tricarboxylic acid cycle, which is essential for tumor cell growth. In addition, elevated GDH expression and activity are observed in glioma and colorectal cancer, and high GDH expression predicts poor outcome. 2,3
Interestingly, Escherichia coli GDH has just a catalytic domain, whereas human GDH contains both an active site and an additional antenna domain (Fig. 1A, B), which can be allosterically regulated by a series of regulators such as adenosine diphosphate (ADP) and guanosine triphosphate (GTP). ADP activates GDH, whereas GTP inhibits the enzyme. Mutations at the antenna domain result in loss of GTP inhibition and over-activation of the GDH enzyme, which causes characteristic hyper-insulinemic hypoglycemia with concomitant hyperammonemia in hyperinsulinism–hyperammonemia syndrome (HHS) patients. 4 –6 Currently, most reported GDH inhibitors are various phenol derivatives such as epigallocatechin gallate that hijack the ADP activation site. 7
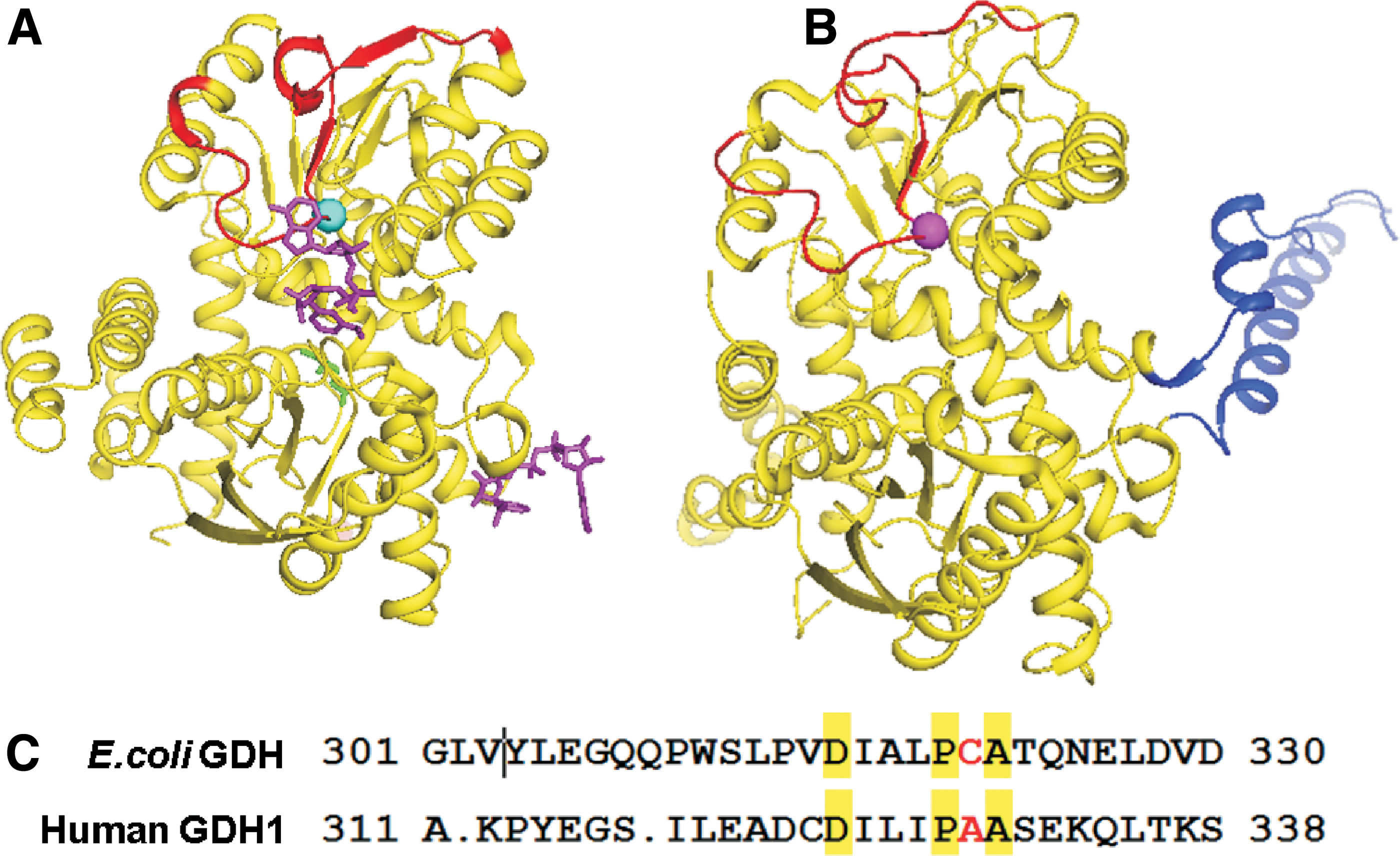
Recently, we discovered that Ebselen could covalently bind to E. coli GDH at the active site cysteine residue (Cys321). 8 However, based on the sequence alignment, the residue corresponding to Cys321 is Ala329 in human GDH (Fig. 1C). To investigate Ebselen inhibition of human GDH, we cloned and expressed an N-terminal His-tagged human GDH in E. coli, and we developed a sensitive 2-(3-(2-methoxy-4-nitrophenyl)-2-(4-nitrophenyl)-2H-tetrazol-3-ium-5-yl) benzenesulfonate sodium salt (EZMTT)-based GDH assay that is highly sensitive and suitable for high-throughput screening for potent GDH inhibitors. In addition, we developed ForteBio-based binding assays to demonstrate that Ebselen is a reversible active site inhibitor for human GDH.
Materials and Methods
Escherichia coli BL21(DE3) strain and E. coli DH 5α strain were purchased from Vazyme (Nanjing, China). pG-KJE8 vector was purchased from YouBio (China). Chemicals were purchased from Aladdin (California). EZMTT detection reagents were from Jennifer Biotech, Inc. (JNFbio; Hangzhou, China). GDH reaction buffer contains 50 mM Tris, 0.003% Brij-35, 0.001% Tween 20, 0.01% bovine serum albumin (BSA), pH 8.0. The UV absorbance was measured by Flexstation 3 (Molecular Device).
Escherichia coli GDH was cloned, expressed, purified, and tested in enzymatic assays as previously described.
9
All biosensors were purchased from Pall Fortebio, Inc. Microplates were supplied by the Greiner Bio-One, Inc. Pall Fortebio Octet® K2 was purchased from ForteBio, Inc. (CA), and Flexstation 3 was from Molecular Device (CA). EZMTT detection reagents were from Jennifer Biotech, Inc.
Cloning and Purification of the Human GDH
Human GLUD1 (GDH1) gene (GI: 2746) encoding the full length of human GDH 1 without the signal peptide was PCR amplified in its entire length by using a forward primer 5′-TATAGGATCCATGAGCGAGGCGGTGGCCG and a reverse primer 5′-TATACTCGAGTTATGTGAAGGTCACACCAGCTTC, to introduce BamHI and XhoI restriction sites. The purified PCR product was restriction digested and ligated into the pET 30a expression vector, and it was used to transform the DH 5α strain of E. coli. After DNA sequencing, the plasmid was transformed into an E. coli BL21 (DE3) strain with pG-KJE8 vector. The recombinant N-terminal His6-tagged GDH protein was expressed after 0.5 mM isopropyl β-
GDH Activity Assay
A two-fold dilution series of the purified human GDH (400, 200, 100, 50, 25, 12.5, 6.2, 3.1, 1.6, 0.8, 0.4, and 0 nM) was added to 96-well plates in reaction buffer (50 mM Tris-HCl, 0.01% BSA, 0.003% Brij-35, 0.001% Tween 20, pH 8.0); then, to each well was added a mixture of glutamate (10 mM final), NADP+ (200 μM final), and the EZMTT detection reagent. The reaction was carried out at room temperature, and absorbance measurement at 450 nm was taken every 5 min for 2 h. The utilization of NAD+ as the human GDH1 enzyme cofactor was tested by using the same procedure as described earlier, except that the NADP+ was replaced with NAD+. The ADP activation assay was tested by using the same procedure as described earlier, except that a mixture of glutamate (10 mM final), NADP+ (200 μM final), and the EZMTT detection reagent with or without ADP (400 μM final) was added.
GDH Inhibition Assay
A series of three-fold dilutions of inhibitors (0–13 μM; final) were added in 96-well plates and mixed with E. coli GDH (1 nM) in buffer (50 mM Tris-Cl, 0.01% BSA, 0.001% Tween 20, pH 8). After 0.5h incubation, to each well was added a mixture of glutamate (10 mM; final), NADP+ (150 μM; final), and the EZMTT detection reagent. The reaction was carried out for 1 h, before the absorbance measurement at 450 nm. To exclude assay artifacts, controls with no GDH were also carried out.
The assay for human GDH is performed essentially the same as the E. coli enzyme except that 50 nM human GDH was used in the presence or absence of 200 μM ADP. The z factor was measured by using 50 nM human GDH as a positive control and with no GDH as a negative control.
Chemical Synthesis of Ebselen Derivatives
Figure 3 shows the chemical synthesis of compound 2, a biotinylated ebselen. To a round-bottom flask, biotin (49 mg, 0.2 mmol), 1,1-carbonyldiimidazole (65 mg, 0.4 mmol), and N,N–dimethylformamide (DMF) (2 mL) were added. The mixture was heated and stirred in an oil bath at 60°C for 0.5 h, and then it was cooled on an ice bath. The mixture was then treated with m-phenylenediamine (216 mg, 2 mmol) in DMF (0.5 mL) and stirred for overnight. The solvent was evaporated under reduced pressure, and the crude product was further purified by silica gel column chromatography (eluent DCM–MeOH, 10:1) to obtain a solid compound 1; Yield 63.7 mg (95%).
To a round-bottom flask, compound 1 (50 mg, 0.15 mmol), Et3N (20 mg, 0.2 mmol), and dry DMF (1.0 mL) were added. To the mixture, a solution of the selenium chloride reagent (38 mg, 0.15 mmol) in DCM (2 mL) was added dropwise at 0°C. The reaction was warmed to room temperature and stirred for 0.5 h. The solvent was evaporated in vacuum. The crude product was purified by flash column chromatography to obtain compound 2 (eluent DCM–MeOH, 20:1). Yield 67 mg (87%), pale-yellow solid. 1 H NMR (500 MHz, DMSO-d6) δ 10.90 (s, 1H), 10.19 (s, 1H), 8.60 (d, J = 8.0 Hz, 1H), 7.88 (t, J = 2.0 Hz, 1H), 7.84 (dd, J = 7.5, 1.0 Hz, 1H), 7.58 (t, J = 7.5 Hz, 1H), 7.53 (d, J = 8.5 Hz, 1H), 7.41 (t, J = 7.0 Hz, 1H), 7.36 (d, J = 9.0 Hz, 1H), 7.29 (t, J = 8.0 Hz, 1H), 6.46 (s, 1H), 6.39 (s, 1H), 4.38–4.12 (m, 1H), 3.14 (t, J = 9.5 Hz, 1H), 2.83 (dd, J = 12.5, 5.0 Hz, 1H), 2.59 (t, J = 6.0 Hz, 1H), 2.34 (m, 2H), 1.73–1.31 (m, 6H). MS: ESI (MeOH), [M + H]+ m/z: 517.1, [M + Na]+ m/z: 539.1.
Biomolecular Interaction Assays
The streptavidine biosensors were loaded with the biotinylated Ebselen (100 μM) or biotin control by dipping into the compound solution in PBST buffer (phosphate-buffered saline, 0.00015% Tween 20, 0.02% BSA buffer) for 10 min. The binding of compound 2 to human GDH1 and E. coli GDH proteins was tested by dipping the individual sensor into a dilution series of GDH proteins (0, 0.06, 0.2 μM) for 100 s to measure the binding on-rate (k on) and then to blank buffer PBST to measure the off-rate (k off). The regeneration buffer used is 10 mM Glycine buffer pH 2. In addition, competition assays were performed by dipping the compound sensor into a mixture of 200 nM GDH protein and its substrates (0–5 mM Glu, NADP, or Ebselen) to get the k on and then into the blank buffer PBST to measure the k off rate.
Gel Shift Assays
A series of dilutions (0, 1, 10, and 100 μM) of the fluorescent CPD3 (Fig. 2) or Ebselen were incubated with proteins (final concentration 0.3 mg/mL; GDH) in buffer A (50 mM Tris, 0.003% Brij-35, 0.001% Tween 20, pH 8.0; 20 μL) at room temperature for overnight. Then, the samples were analyzed by both the native gel and the denaturing SDS-PAGE gel assays. The 10% Tris-glycine native gels were analyzed by fluorescent imaging (Ex 340 nm; Em 520 nm), followed by Coomassie stain to observe the crosslink and co-migration of the fluorescent-labeled Ebselen derivatives with the protein complexes.
Chemical structures of four reported GDH inhibitors.
Results and Discussion
Human GDH is an important enzyme playing a critical role in several disease states. However, efficient expression and purification methods for human GDH are not readily available. The reported expression systems were mainly in insect sf9 or sf21 cells. 10 –12 The expression in E. coli was also achieved, but the protein purification required multiple steps of chromatography, including Q-Sepharose Fast Flow anion-exchange column, ω-aminopentyl hydrophobic-interaction column, and GTP-agarose affinity resin column. 13 In some cases, protein denature and re-nature procedures are required for protein purification.
In addition, traditionally GDH enzymatic activity was followed by monitoring the change in NADPH absorbance at 340 nm. 14,15 However, the NADPH method lacks sensitivity and utilizes a large amount of human GDH enzyme that is not readily available.
Therefore, we cloned and expressed a mature form of human GDH enzyme with an N-terminal His-tag, and the purified enzyme was used to develop a sensitive EZMTT-based method for human GDH activity measurement under various conditions. The EZMTT-based assay provided linear initial velocity, high reproducibility, and it was suitable for high-throughput screening of GDH inhibitors.
N-Terminal His-Tagged Human GDH1 Is Active
We overexpressed the N-terminal His-tagged human GDH1 in E. coli and quickly purified the enzyme by using Ni-affinity chromatography. Because the EZMTT assay sensitively detected NAD(P)H and showed good stability under the assay condition, we successfully developed a sensitive and reproducible EZMTT-based assay for E. coli GDH. 9 Accordingly, we configured an analogous assay using EZMTT as the detection reagent 9 (Fig. 4A), and we demonstrated linear initial velocity when 50 nM human GDH1 is used (Fig. 4B). The human version of GDH1 can utilize both NADP+ and NAD+, and better activity was observed with NADP+ (Fig. 4C). Also, the addition of ADP significantly increased the human GDH1 catalytic activity by at least 4-fold (Fig. 4D). These properties are in agreement with the literature, indicating that we successfully expressed an active form of an N-terminal His-tagged human GDH1 protein.

Chemical synthesis of biotinylated Ebselen (compound 2).
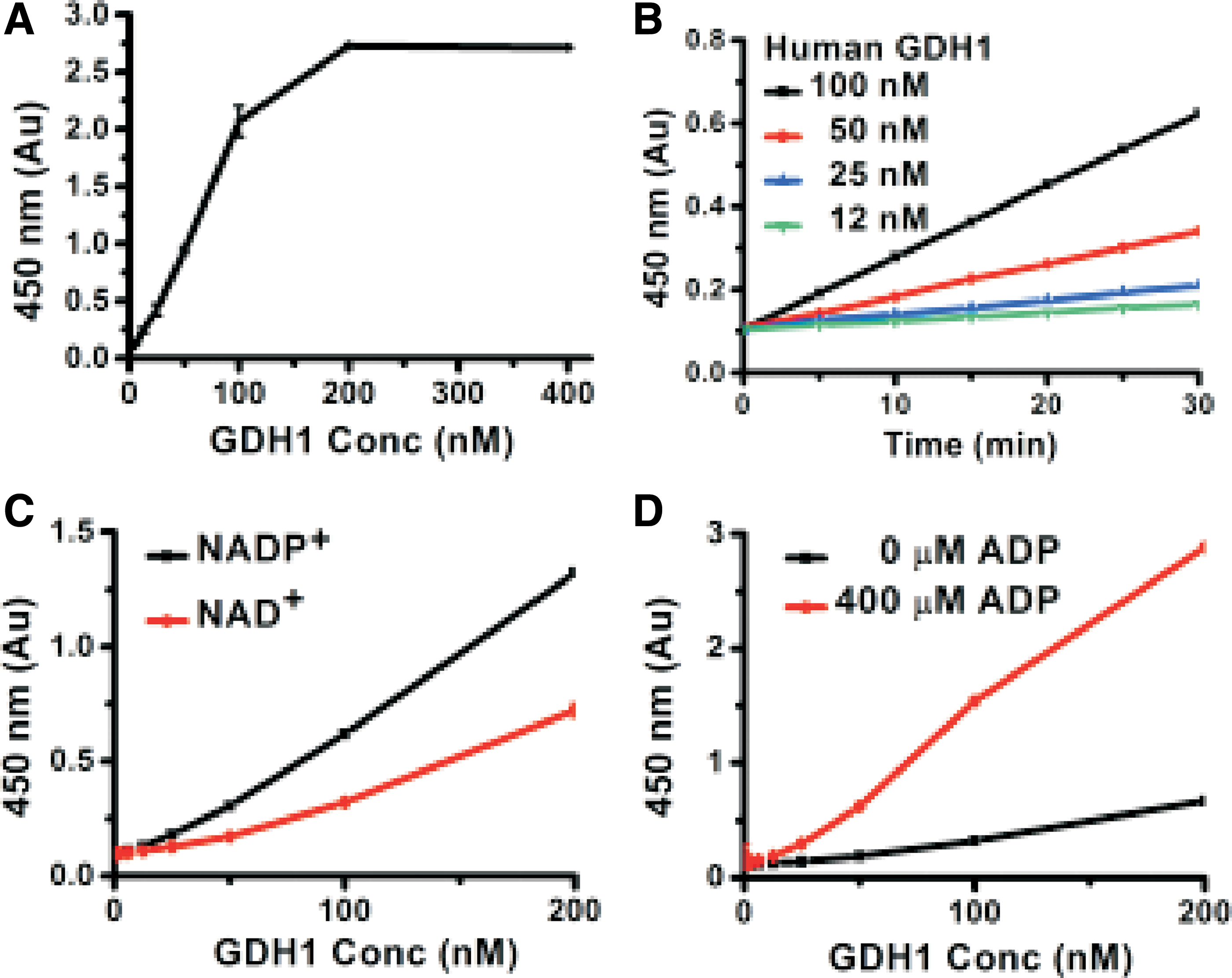
Activity of human GDH1 enzyme.
Ebselen and Its Derivatives Inhibit Human GDH1
Under the optimized assay condition, the EZMTT-based human GDH1 assay showed excellent reproducibility with a z factor of 0.78 (Fig. 5A). Under the optimized condition, Ebselen inhibited both E. coli and human GDH1, although it showed more potency toward the E. coli enzyme than the human GDH1 (Fig. 5B). These results may be explained by the fact that Ebselen could crosslink with the Cys321 residue in E. coli, whereas the corresponding residue in human GDH1 is Ala329, which could not be crosslinked. For further characterization of the reversible binding of Ebselen to the human enzyme, we chemically synthesized a biotin-labeled Ebselen (compound 2). Both compound 2 and the fluorescent-labeled Ebselen derivative (CPD3) showed inhibition of both E. coli and human GDH1 (Fig. 5C, D).
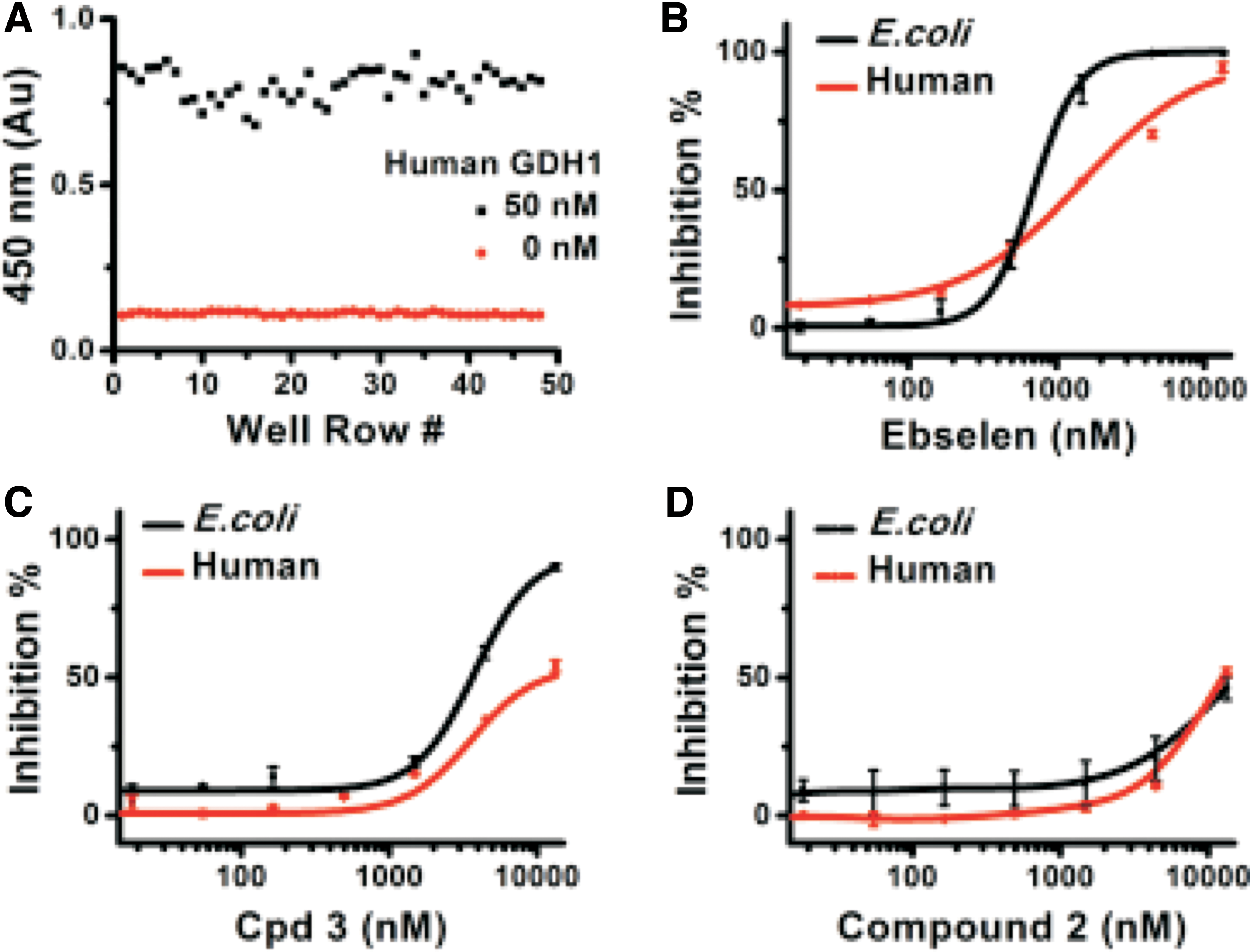
EZMTT dye-based human GDH1 and E. coli GDH inhibition assay.
Ebselen Covalently Binds with E. coli GDH, but Not Human GDH1
Gel shift analysis under native conditions could demonstrate the effect of the compound on protein complex formation and determine whether the fluorescent compound is covalently bound to the proteins. As shown in Figure 6, GDH existed as a hexamer; when CPD3 was added, a dose-dependent increase of fluorescence in a band corresponding to the E. coli GDH was observed. However, under the same conditions, essentially no significant amount of fluorescence co-migrated with the human GDH1, indicating no crosslink. These results further confirmed that because the human GDH1 lacks the corresponding crosslink Cys321, Ebselen could not covalently bind to the human GDH1.

Native gel analysis to identify the covalent linkage between CPD3 and the GDH proteins.
Ebselen Competes Against NADP and Glutamate for Binding to Human GDH1
The Ebselen binding model between E. coli and human GDH1 was investigated by biomolecular interaction assays. As shown in Figure 7, both E. coli GDH and human GDH1 enzymes could bind the biotinylated Ebselen in a dose-dependent manner. Interestingly, the presence of GDH substrates reduced E. coli GDH binding moderately, but it almost completely prevented the human enzyme from binding the immobilized biotinylated Ebselen. These results demonstrated that Ebselen binds reversibly at the active site of the human GDH1 enzyme.
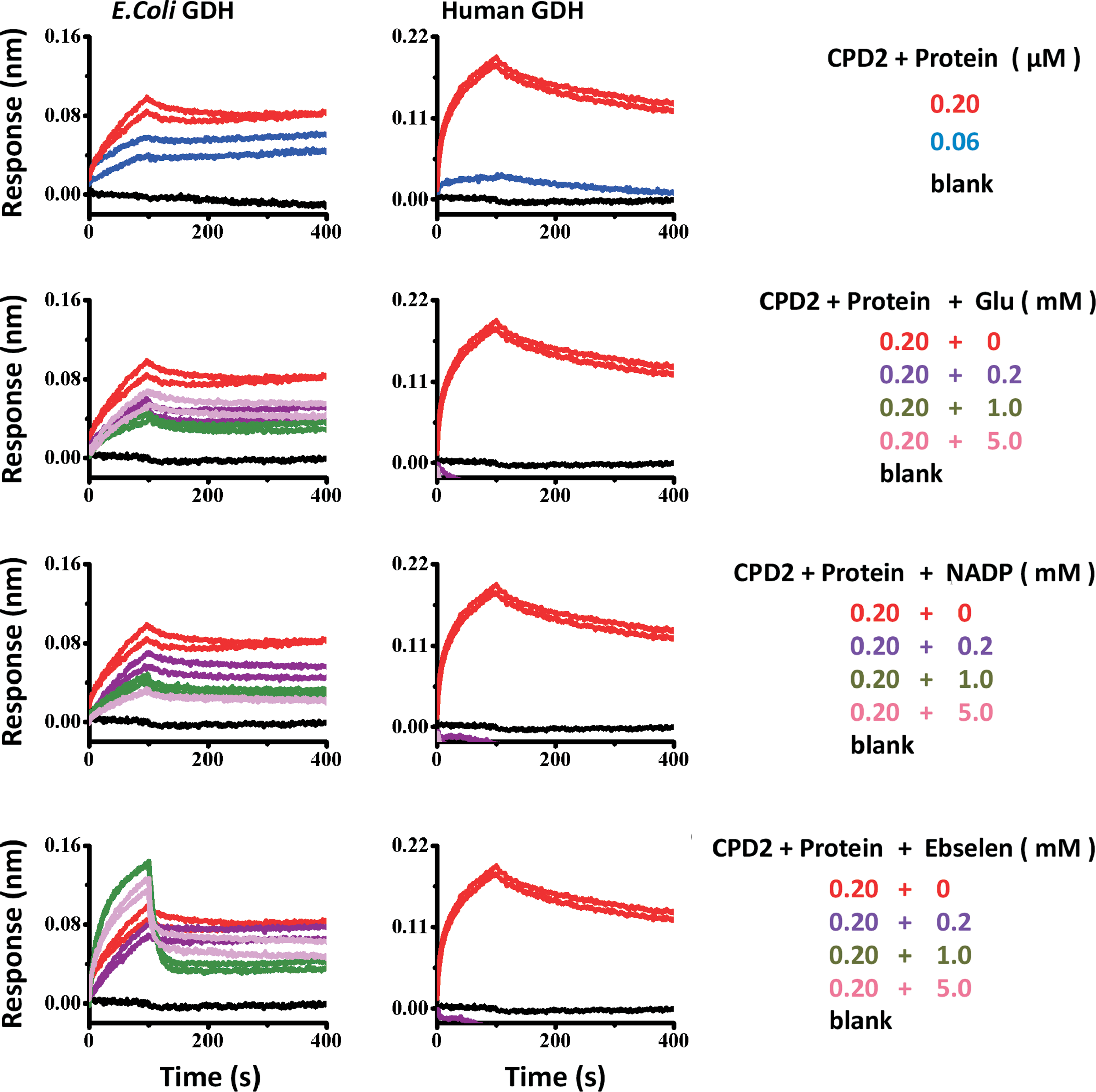
ForteBio analysis of Ebselen binding to human GDH1 and E. coli GDH enzymes in the presence or absence of its substrates (Glu, NADP, Ebselen). Color images available online at
Conclusion
Mutational loss of GTP inhibition causes over-activation of human GDH1 in HHS patients. Our results demonstrated that Ebselen is a reversible active site GDH inhibitor. Since Ebselen 16 –19 is a multifunctional organoselenium compound in Phase III clinical trials for inflammation, a more potent Ebselen-based GDH inhibitor might be valuable for future drug discovery for HHS patients. 4 –6
Footnotes
Author Contributions
B.H.R. designed the experiments and wrote the article; others performed experiments. All authors read and approved the article.
Acknowledgments
The authors appreciate the financial support from QinLan Grant award from Hangzhou innovation foundation (Grant H1160492) and Zhejiang University of Technology (Grant 414800129).
Disclosure Statement
No competing financial interests exist.