
Abstract
Human steroid 5 alpha-reductases (S5αRs) and NADPH irreversibly reduce testosterone to the more potent dihydrotestosterone (DHT). S5αR inhibitors are useful treatments for DHT-dependent diseases, including benign prostatic hyperplasia, androgenic alopecia and hair growth, and acne. There are three S5αR isozymes, and there is a need for safer and more isozyme selective inhibitors than finasteride and dutasteride currently licensed. In this study, we review the methods used to screen for S5αR inhibitory activity and describe studies that characterize the ability of herbal preparations and their constituents to inhibit S5αRs. We identified enormous variations between studies in IC50s for finasteride and dutasteride used as standards. Accordingly, we make several recommendations: Stable isozyme specific transfection systems need creating a standardized enzyme/microsome preparation and all three isozymes, as well as androgen receptor binding, should be tested; agreed reaction conditions, especially the substrate concentrations, and separation/quantitation method optimized for high throughput screening; systematic screening of herbal compounds and most extensive use of leads to develop more potent and isozyme specific inhibitors.
Introduction
Androgens, especially testosterone, play a crucial role in secondary sex characteristic development of both sexes. Many tissue specific and localized testosterone actions are through dihydrotestosterone (DHT) that is produced within target cells by testosterone by a reduction reaction using steroid 5α-reductases (S5αRs). The inhibition of these enzymes provides a way of selectively blocking some androgen actions while leaving others intact, including androgenic and androstatic actions on hair growth and acne. Many studies have searched for herbal compounds that could tap into the growing market for topically applied medicines to treat these conditions.
In this review, we aimed to summarize our current understanding of the biochemistry involving steroid 5 alpha-reductases (S5αRs) and their role in physiological signaling. We then review the various protocols to quantify the actions of S5αR inhibitors, describe the actions herbal compounds have on these enzymes, and assess their clinical potential. We identify a lack of standardization in these protocols and suggest ways to address problems in the development of herbals into clinically useful treatments.
Physiology and Pathologies Associated with Androgens
Testicular interstitial (Leydig) cells are the main source of circulating testosterone and are regulated by the hypothalamic anterior pituitary axis and luteinizing hormone release. These are under androgen feedback regulation. Total blood testosterone is ∼15 nM of which ∼98% is bound to sex hormone binding protein and albumin 1 but only free testosterone (∼0.2 nM) is likely to be physiologically active. Total blood DHT is 0.5–3 nM, and its free concentration correspondingly lowered by protein binding.
Testosterone (T) is pivotal in male differentiation and maintenance of secondary sexual characteristics. Classical testosterone targets are the genitalia, hypothalamus, bone, skeletal muscle, brain, and omental fat acting through the androgen receptor (AR). 2 In male embryonic stage, testosterone stimulates the differentiation of the Wolffian duct into male internal genitalia such as epididymis, vas deferens, and seminal vesicles. At puberty, testosterone modifies the external genitalia, increases muscle mass, develops vocal cords, develops male sexual behavior, libido, penis, hair, prostate, scrotum, and initiates spermatogenesis. 2,3 In addition, androgens improve immunity and tissue repair, reduce nonomental fat accumulation, improve insulin sensitivity, and improved vascular function (contrary to the popular notion). 2
Many androgenic cellular actions are fulfilled by S5αR reducing testosterone to more biologically active androgen, DHT, within target cells. 4 In some tissues (prostate, skin, hair follicles, etc.) cytosolic DHT is 3–10-fold higher than total blood levels. There is no evidence of active DHT sequestration so these higher concentrations must arise from DHT production in the target cells, mainly from extracellular testosterone. Compared with testosterone, DHT has 2–5 greater affinity for ligand binding site, thereby prolonging AR-mediated DHT signaling. 2,5 DHT is also essential for fetal sexual differentiation and growth of the prostate gland, male external genitalia, and pubertal growth of facial and body hair. 2
Classical androgen signaling is through cytosolic ARs leading to nuclear translocation and further complexation into fully competent activators of the genes 4,6 bearing androgen response elements. These AR-regulated genes promote cell differentiation and maturation. 7,8 ARs are coded by a single gene expressing 19+ splice variants of which only 4 are ligand activated. Full-length ARs have features common to other nuclear receptors. Some variants are constitutively active in hormone-resistant prostrate tumors and selectively activate cell-cycle genes and cell proliferation. 9 Other AR constructs are anchored to the plasma membrane and signal through MAPkinase indirectly acting on nonandrogenic genes. Agents interacting with nonandrogenic binding sites on the AR are undergoing clinical trials. 10 Variants are also expressed in cancerous gynecological tissues and breasts. 8 There appears to be no information on the physiological role of these variants in other tissues. But a minority of patients having nonhormone dependent prostate cancer and who express the AR-V7 construct paradoxically show tumor regression with testosterone treatment. If scalp hair follicles also express this same variant, it would explain the paradoxical action of DHT in balding compared to other hair follicle types.
S5αR: Biochemical and Pharmacological Properties
The major physiological source of DHT is through NADPH/S5αR hydrogenating testosterone. Human S5αR (hS5αR)(or 3-oxo-steroid-4-ene dehydrogenase, EC 1.3.99.5) is a membrane bound enzyme that irreversibly reduces 4-ene-3-oxosteroids to 5-alpha-3-oxosteroids using NADPH as the reductant (Fig. 1). 6 The proposed chemical mechanism is the direct donation of a hydride (H−) from NADPH to the C-5 position of testosterone (Fig. 1). The enolate system is formed due to the activation of the enone in testosterone by interaction with an electrophilic pocket (Enz+) in the enzyme active site. This leads to a positively polarized species that accepts a hydride from NADPH at C-5 of testosterone. Then, enzyme-mediated tautomerism provides the product DHT and release of NADP+. 11,12
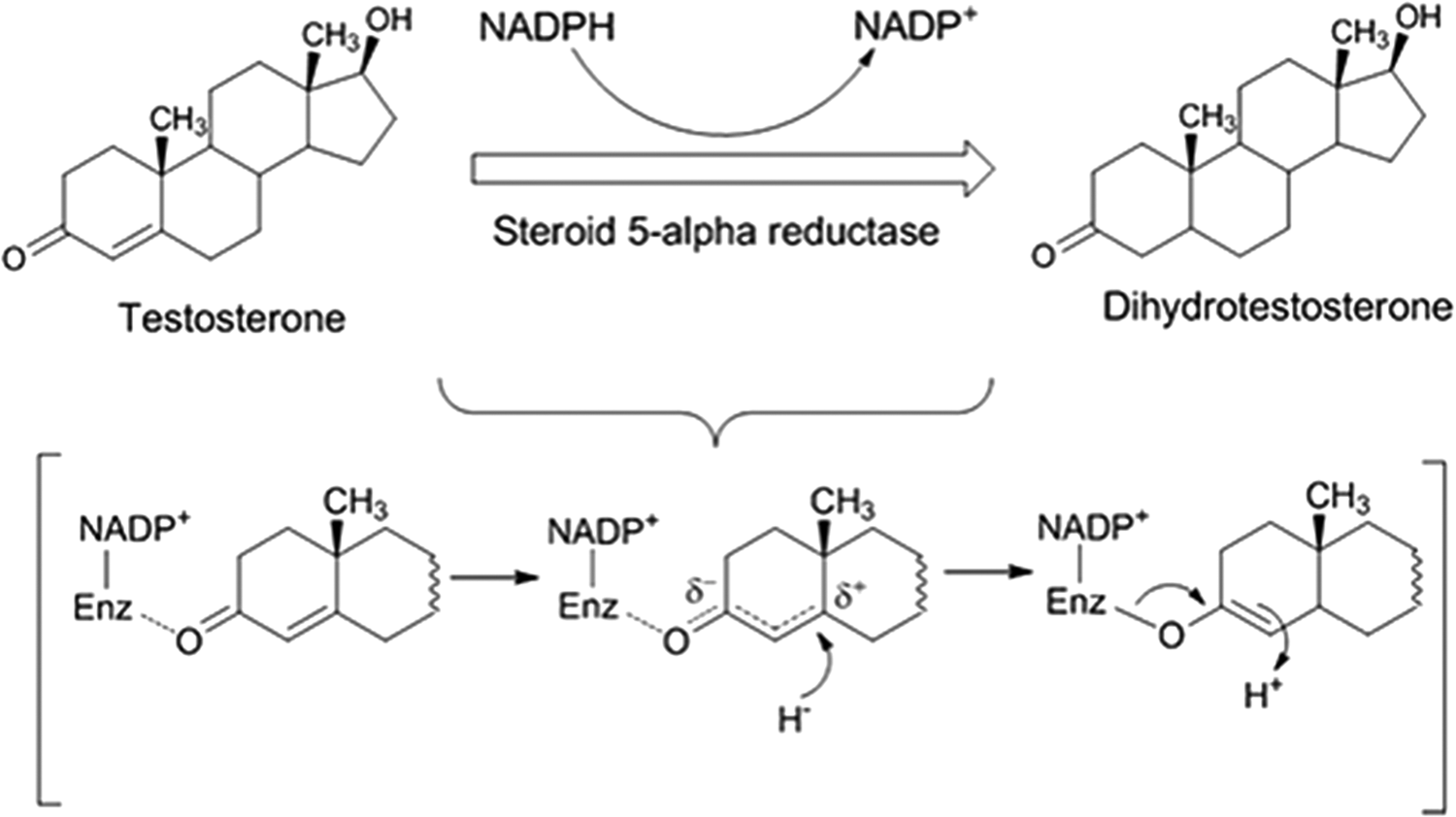
The enzymatic reaction catalyzed by steroid 5α-reductase and the purposed biochemical mechanism.
The prostate also has a limited capacity to produce DHT from dehydroepiandrosterone (DHEA), androstenedione, and other upstream precursors coming from the adrenals 2 also using S5αR. However, aggressive hormone resistant prostatic cancer is able to constituently make DHT independent of testicular steroids. 13,14 Extensive evidence from the prostate indicates that DHT tissue contents are unaffected by high serum concentrations of testosterone and DHT. Male balding is also independent of serum testosterone or DHT over a wide range of concentrations and even seen in hypogonadal men.
S5αR exists as three isozymes (S5αR type 1–3) coded by the 5-membered SRD5A gene family of which SRD5A-1 to -3 code for functional S5αRs. There are important differences in biochemical and pharmacological properties, tissue distribution, and chromosomal location while sharing the same substrate preference for types 1 and 2 (Table 1).
Comparison of S5αR Isozymes *
SRD5A3 is widely expressed in humans and animals, including skin as protein and Mrna. 16 While S5αR type 3 may produce DHT from testosterone, other expression systems for SRD5A-3 failed to reduce testosterone, progesterone, androstenedione, or corticosterone. 17 In human abdominal preadipocytes expressing S5αR type 3, hydrogenated androstenedione preferentially produce DHT. 18 However, SRD5A-3 mutations cause no developmental sexual defects, 19 but they do cause ocular and cerebral dysfunction in humans and other defects ascribed to failure to N-glycosylate proteins and similar reactions. These mutations prevent conversion of polyprenol to dolichol by “polyprenol reductase,” a synonym for S5αR type 3. Thus evidence at present suggests that SRD5A-3 may be a pathway specific S5αR or polyprenol reductase in normal tissues depending on conditions and cell type. Notably, the overexpression of S5αR type 3 isozyme is observed in breast cancer, lung adenocarcinoma, testicular seminoma, and castration-recurrent prostate cancer. 16,20,21 There is no evidence that SRD5A-4 and -5 mediate DHT production.
These reductases are also involved in four other steroid transformations, including other pathways contributing to DHT production through the classic pathway using testosterone, the “back door route” using progesterone intermediates, and the third by the androstanedione route. 22 These alternative pathways have practical importance in castration resistant prostate cancer where the cells continue to elaborate DHT despite low serum testosterone. Alternative pathways might explain the limited inhibitory action of our herbal S5αR inhibitors in only partially depressing axillary hair growth. 23,24 For castration resistant prostate cancer, there is a search for inhibitors of 17, 20 lyase 25 to supplement S5αR inhibitors and block this alternative DHT production. Expression of human S5αR isozymes in specific tissues/cells has been reported using different techniques such as ELISA, protein expression, western blot, northern blot, mRNA, and enzymatic activity, which have shown different S5αR expression in different tissue/cell of enzyme origins (cited in Ref. 26 ). Tissue localization of the three isozymes in human is summarized in Table 1. Type 1 isozyme is mainly located in the liver, nongenital skin, and hair follicles, while Type 2 isozyme is predominantly expressed in the genital area, skin, prostate, and hair follicles of the scalp. Type 3 is mainly expressed in various benign and malignant tissues. Types 1 and 2 share moderate sequence homology of ∼50%. However, they differ with respect to their biochemical properties. For example, Type 2 is active over a narrow acidic pH range (4.5–5.5), whereas Type 1 has the most favorable pH at broad neutral to basic (6.5–8.5). 6,26,27 Optimal pH of isozyme 3 is 6.9. 20,26 The optimal pH information is useful for specific isozyme S5αR inhibitory assay development. Under optimal conditions, the type 2 isozyme has a higher V m/K m indicating a higher S5αR activity than the type 1 isozyme. 28 –30 Isozyme 1 has a higher turnover number, as pointed by its K cat value and a lower substrate affinity for testosterone, K m ∼2 μM. S5αR type 2 has a lower turnover number (K cat) and a higher substrate affinity, as indicated by K m ∼0.2 μM for testosterone substrate. The apparent dissociation constant of NADP+ for both isoenzymes is similar (3–10 μM). 6,29 Previous studies have suggested that the N-terminal hydrophobic region of types 1 and 2 could interact with the aliphatic and aromatic side chains of substrates and that some of the C-terminal pocket, such as H232, could play critical roles in the catalytic activity and K m of the testosterone substrate. 31,32 Crystal structures of S5αR isozymes by X-ray diffraction have not been established yet due to their instability during purification.
Pathologies Targeted by S5αR Inhibitors
S5αR overexpression elevates cellular DHT 6 but not necessarily causes 2 some androgen-dependent disorders, including benign prostatic hyperplasia (BPH), progression to prostate cancer, and male androgenic alopecia (AGA) or baldness, all of which increase with age. 33,34
Prostate cancer is the most common neoplasm in men and thus attracted intense study, including its treatment by S5αR inhibitors. However, neither finasteride nor dutasteride are recommended as prophylactics nor for inhibiting tumor progression. These are effective in prostate cancer because of increased AR sensitivity and increased DHT production by prostate cancer cells from upstream precursor steroids. 26 Thus, the therapeutic role of S5αR inhibitors is likely to be limited to one component of a polypharmaceutical treatment.
Both S5αR type 1 and 2 are found in the skin but their apparent localization appears to depend on skin type, location, species, and protocol used. 26 The folliculosebaceous unit mainly expresses both S5αR type 1 and 2.
Acne 35,36 involves excessive DHT dependent secretion of sebum into hair follicles. Again, DHT is produced locally within the secretory cells through S5αR type 1 and 3 (in a cell line). 37 A large number of treatments have been tested, including S5αR inhibitors, antimicrobials, 38 and antibiofilm, 39 many of which are systemically administered and, thus, causing systemic adverse reactions. Thus, topical application would appear safer and application restricted to areas causing most disfigurement. In this study, herbal compounds have some potential since there is a prospect of using a monotherapy that combines antimicrobial, antibiofilm, and S5αR inhibitory actions. Excessive and inappropriate hair growth is another pathology mediated by DHT and has been applied to hirsutism, axillary hair, and AGA. Incidences of hirsutism are 5%–10% in younger women and ∼80% in polycystic ovary syndrome and are also disfiguring. Most pharmacological treatments have been through the oral route thus acting systemically and include S5αR inhibitor (finasteride) and AR antagonists that modestly inhibit hair growth with prolonged (>6 months) treatment. 40,41 However, they all produce site effects where risk versus benefit limits their clinical utility. Finasteride also partially reduced facial hair density (∼30%) and shaft diameter when applied topically. 42,43
Androgen axillary hair is another source of embarrassment for women, and limited evidence suggests that S5αR inhibitors might be partly efficacious. 23
AGA is caused by localized DHT production through S5αR type1 to inhibit hair growth on the scalp through multiple mechanisms, some of which are gender specific. 44 –46 It increases with age suggesting a senesce component where hair transplants or stem cells may be efficacious. 46 There are numerous “hair restorer” preparations acting on many molecular targets and others whose mechanisms of action are obscure. Targets include S5αR inhibitors such as oral finasteride while there are also unapproved herbal S5αR inhibitors (Table 3). Again, these are only partially effective, so supplementing with compounds acting on related molecular targets may have more complete clinical outcomes.
Adverse Effects of S5αR Inhibitors
S5αRs can also reduce other steroids in humans, including progesterone to dihydroprogesterone (precursor of neurosteroids) 47 and cortisol to the less active dihydrocortisol. 48 Accumulating cortisol has the potential of creating a Cushing's type phenotype and insulin resistance. 49,50
Common adverse effects in men on finasteride and dutasteride are sexual dysfunction, infertility, mood disorders, gynecomastia, and raised cardiovascular morbidity/risk factors. 51,52 A persistent adverse side effect of S5αR inhibitors is so called postfinasteride syndrome after stopping finasteride treatment described in its postmarket report. 51 Both finasteride and dutasteride cause erectile, libido, and ejaculatory dysfunction in BPH patients. 53 The psychiatric adverse effects could arise from reduced dihydroprogesterone production. 54 This leads to depressive symptoms and self-harm. 55
While these safety risks may be acceptable in prostrate hyperplasia and cancer, more frivolous uses such as modifying hair growth and acne are more amenable to topical application.
In Vitro Assays for S5αR Inhibition
Table 2 lists assay conditions in studies that mostly assessed inhibition by herbal substances, but the list is not exhaustive. Most studies used free enzyme, as lysates, or subcellular fractions (Table 2). In whole-cell tests, the enzyme is located in either cells related to the target tissue pathology (dermal or prostate) and related cell lines or in expression systems. In this study, inhibitors have to gain access to the cell interior but are impenetrable by many polyphenols. 56 They are also vulnerable to off target or toxic actions and vagaries arising from the metabolic state of the cell. Alternatively, for drug screening, especially inhibitors, cell-free protocols facilitate high throughput screening, mechanistic studies, and more controllable conditions.
Summary of S5αR Inhibitory Assay
1, S5αR type 1; 2, stands for S5αR type 2; DHT, dihydrotestosterone; EIA, enzyme immunoassay; GC, gas chromatography; HPLC, high performance liquid chromatography; MS, mass spectroscopy; NR, not reported; T, testosterone; TLC, thin layer chromatography.
The most common S5αR assay strategy has been to measure declining substrate concentration, commonly radio labelled testosterone, while two studies used NADPH consumption and a few monitored DHT formation which is a more selective and reliable end point 57 –64 (Table 2). For the enzyme, about half the assays used either or both S5αR type 1 and type 2 but not reported the other studies. The origin of the enzyme came from a range of tissues (prostate, skin, liver, dermal papillae, keratinocytes, fibroblasts, or cDNA expression systems) and species (rat or human).
Some assays have no need for sample preparation. Enzyme immunoassay 57,65 relies on antibodies but can be unreliable while UV-microplate format has poor selectivity and sensitivity. 66,67 Separating testosterone from DHT is particularly problematic, and separation of other reactants, cofactor, and contaminants has used chromatography, thin layer chromatography, 61,68 –71 high performance liquid chromatography, 72 –74 and gas chromatography. 75
Quantification of the indicator substance (substrate, cofactor, or product) used radioassay 58,61,70,72 (and more references in Table 2), mass spectroscopy (MS), 75,76 UV spectroscopy, 67,73,74,77 or enzyme immunoassay. 57 The conventional and most sensitive method is radioassay. 64,78 –80 However, increased requirements for dedicated facilities and safety are making radioassay problematic. MS has attracted interest due to its high sensitivity and selectivity. 63,75
No study has considered AR blockade which would also contribute to efficacy while being safe with topical application.
Finasteride is commonly used as a positive control yet its IC50 on allegedly S5αR-2 varies by 300-fold (0.1–32 nM) demonstrating a dire need for standardization of assay conditions for assessing inhibitor potency. Enzyme homogeneity is probably the most troublesome due to: (1) inability to purify active enzyme and different isolation methods, (2) differences between rat and human enzyme, (3) other interfering isozymes (e.g., S5αR-3 is widely distributed and when active, very sensitive to finasteride), and (4) different cell types or expression systems that may express other interfering proteins. There is clear need for a standardized enzyme and most appropriately would come from three cell lines stably incorporating human SRD5A-1, -2, or -3. A fourth line transfected with empty vector should be an enzyme blank. The S5αR activity should be mostly in the microsomal component of the fractionated cell homogenate. Other incubation conditions such as the medium specification and NADPH and substrate concentrations should be agreed upon.
Herbal S5αR Inhibitors
Herbal medications are becoming increasingly popular because of their perceived safety. 103 To satisfy this demand, many natural products have been isolated that display S5αR inhibition. These are summarized in Table 3.
S5αR Inhibitors from Natural Products
Ext, extract; H, human; HL, human cell line; HR, human recombinant; NR, not reported; P, pure compound; R, rats.
Although the list is not exhaustive, it demonstrates S5αR inhibition by a diverse range of mostly plants and/or single chemical constituents thereof. Some studies did not report the isozyme tested, and others used S5αR type 1 or 2, or both, and used sources or constructs from a variety of animals and assay conditions. No study assessed AR blockade. Furthermore, many studies tested herbal compounds already known to have many other biological actions on well characterized molecular targets, for example, biochanin A, daidzein, and genistein. 56,87 These are known to be agonists at estrogen receptors at lower concentrations than used here and act on many other targets so cannot be selective for S5αR. Several studies were prompted by traditional uses on hair and skin (Table 3). However, traditional uses need confirmation by beginning with robust clinical trials. 104 Although the herbals may demonstrate S5αR inhibition, it may have more potent actions on the other multiple signaling pathways that control hair growth, etc. If it explored, such actions could lead to valuable adjuncts to S5αR-based treatments.
IC50s (or equivalents) of herbals ranged 0.3–40,000 μg/mL, while the maximum inhibition was commonly well below 100%; without an I max, the IC50 is an estimate. Many studies did not report IC50. While a few studies were translated to inhibiting mouse or human hair growth (not shown in Table 3), in vitro studies showing low potency (e.g., IC50 > 100 μg/mL) are unlikely to achieve amounts in topical or oral preparations that show efficacy in vivo. No tested substance reached the high potencies of many synthetic compounds of IC50 < 100 nM (∼50 ng/mL). However, many of these highly potent antagonists may preferentially bind to the NADP-enzyme complex in a manner similar to finasteride action. A way around this potency discrepancy was addressed in three studies that used herbal compounds as leads for structure/activity studies. 56,76,82
Efficacy and Safety Trials
Although finasteride is widely used as an S5αR type 2 inhibitor, its oral use is only justifiable for prostate hyperplasia in a polypharmaceutical role. Other treatments are localized treatments of the integument where finasteride is topically active. Most synthesized and herbal S5αR inhibitors are, or appear to be, nonpolar and are likely to penetrate and tend to stay in the stratum corneum of the skin. 110 In this study, the active inhibitor needs formulating into a suitable vehicle to enhance cutaneous application and has sufficient potency to compensate for the low penetration rate. To achieve this, the drug needs applying at high concentration or improving skin penetration by drug delivery system.
Most preclinical studies on herbal medicines are rarely translated to humans due to ambiguous output for translation. Clinical trials are more cost-effective and precise than preclinical studies. They begin with safety evaluation, the 4-h human skin patch test for skin irritation, and challenge test for skin allergy. A trial needs ∼20 participants to whom the intervention is applied with roll-on applicators to conceal any differences between test and placebo formulations randomly applied on opposite sides of the body using full blinding. 23,24 This should be weighed before and after the treatment course to assess adherence and the amount applied. For efficacy trials studying topical application, the treatment should be confined to a skin area on one side of the body and a symmetrically contralateral area for placebo treatment, the two areas being randomly allocated to test and control. Thus participants act as their own controls. A test area should be a circle 1–4 cm in diameter depending on the primary end point and measurements. This would be about 1% of body surface so systemic absorption would have minimal effect on blood concentrations.
The protocol should have adequate powering, safety monitoring, and duration (∼6 months). Designs must incorporate the CONSORT 2010 and herbal checklists and consider the Cochrane risk of bias scale, v2.
Recommendations
Herbal compounds offer enormous scope in treating DHT mediated conditions. However, if candidate compounds are to be treatments, we recommend the following considerations: Screening sources of enzyme from cells that stably express human S5αR-1, -2, or -3, which also enables standardization between laboratories. These cells should be freely available. The additional assay for screening of AR blocking and 17,20-lyase inhibition should also be considered. More information about the relationship of physiological roles to the three S5αRs should be explored. Liquid chromatography (LC) or gas chromatography (GC) offers reliable substrate/product separation in assays. The MS detection is safety-conscious technique in comparison to radio-labeling detection, and it is also amendable to higher throughput. Screen herbal compounds having structures similar to 4-ene-3-oxosteroids as blockers or obstructive substrates like finasteride. Since specific S5αR inhibition is only partially effective, the role of herbal adjuncts acting on related processes involving the target pathology should be explored to improve clinical effectiveness. Studies prompted by traditional medicines must initially demonstrate clinical efficacy. Functional inhibition: screen targets in whole-cell studies to demonstrate that DHT production is actually inhibited. Bioavailability: if treatment is intended to be oral, pharmacokinetics/metabolism must be assessed in humans. Clinical trials: these must have robust design and be free of biases and include detailed safety monitoring.
Conclusion
Development of S5αR inhibitors as cosmetics has been haphazard, and few studies using natural products have translated into clinically efficacious products. A wide array of chemical structures from a diverse range of plants appears to inhibit testosterone reduction using in vitro systems. However, many of the herbal extracts or individual compounds tested had high IC50s, and some have numerous and well-known actions on other biological targets. The methodologies used were diverse as exemplified by the extreme range of comparator drug potencies and prevented comparisons being made between studies.
To address these issues, there are eight recommendations, including creating isozyme specific and stable S5αR expression systems as sources of enzymes and standardized reaction conditions.
Footnotes
Acknowledgments
The funding provided by The Center of Excellence for Innovation in Chemistry (PERCH-CIC), Office of the Higher Education Commission (Ministry of Education); the Thailand Research Fund (the grant nos. DBG608005 and IRN58W0005); and Naresuan University, Thailand are gratefully acknowledged.
Authors' Contributions
J.S., G.M., and C.N.S. wrote the article and K.I. and C.N.S. checked and edited the article.
Disclosure Statement
No competing financial interests exist.