
Abstract
Fibrosis is defined by excessive production of type I collagen in various organs. Excessive type I collagen production in fibrosis is stimulated by binding of RNA protein LARP6 to the structural element of collagen mRNAs, the 5′ stem loop (5′SL). The LARP6-dependent regulation is specific for type I collagen and critical for fibrosis development. Inhibitors of LARP6 binding have potential to be specific antifibrotic drugs, as evidenced by the discovery of one such inhibitor. To create technology for phenotypic screening of additional compounds we developed an inverted yeast three hybrid system. The system is based on expression of human LARP6 and a short RNA containing the 5′SL of human collagen α1(I) mRNA in Saccharomyces cerevisiae cells. The cells were engineered in such a way that when LARP6 is bound to 5′SL RNA they fail to grow in a specific synthetic medium. Dissociation of LARP6 from 5′SL RNA permits the cell growth, allowing identification of the inhibitors of LARP6 binding. The assay simply involves measuring optical density of cells growing in multiwall plates and is pertinent for high throughput applications. We describe the specificity of the system and its characteristics for high throughput screening. As a proof of principle, the result of one screen using collection of FDA approved drugs is also presented. This screen demonstrates that using this technology discovery of novel LARP6 inhibitors is possible.
Introduction
Fibrosis is characterized by excessive deposition of extracellular matrix in various organs, including liver, bone marrow, lung, kidney, GI tract, skin, and eye 1 or be associated with trauma, burns, surgical procedures, radiation, and chemotherapy. It has been estimated that fibrotic disorders contribute to about 45% of all-cause mortality in the United States. 2 The most common is liver fibrosis, with prevalence of advanced stage hepatic fibrosis in general population of 2%–4%. 3 The main component of the fibrotic matrix is type I collagen that is a heterotrimeric protein composed of two α1(I) and one α2(I) polypeptides. 4
Recent work has suggested that dramatically increased rate of type I collagen synthesis in fibrosis is not simply an augmentation of the constitutive synthesis, but that activation of an additional mechanism is involved. The core of this mechanism is binding of protein LARP6 to collagen α1(I) and α2(I) mRNAs, which stimulates their translation. 5 –12 Other researchers have corroborated the importance of LARP6 in regulating type I collagen. Insulin-like growth factor-1 (IGF1) increases synthesis of collagen type I by induction of binding of LARP6 to the 5′ stem loop (5′SL). 13 Animals with deficiency of IGF1 receptor downregulated LARP6 and had reduced amount of type I collagen in vascular smooth muscle cells. 14 LARP6 was shown to induce osteogenic differentiation and type I collagen expression in osteoblasts, 15 while irradiation of dermal tissue culture increased LARP6 and type I collagen. 16
In normal tissues type I collagen is one of the most stable proteins with half-life of 4–12 months; its fractional synthesis rate (FSR, defined as % synthesis per day) is about 2% in the skin, 17 while in the liver it is only 0.2%. 18 This is significantly slower than the FSR of proteins in most organs (18%–140% per day). 19 However, at the sites of active fibrilogenesis the synthesis of type I collagen can be increased several 100-fold. 20,21 To accommodate such high synthetic rate a unique bimolecular interaction between a structural element of type I collagen mRNAs, 5′SL, and protein LARP6 is activated. 6 –10,22 –26
Collagen α1(I) mRNA and α2(I) mRNA have an evolutionary conserved secondary structure in their 5′ UTR, termed 5′SL. 5′SL is not found in any other mRNA, except the mRNA that encodes type III collagen. 27 Type III collagen is related to type I, but is only a minor component of the extracellular matrix. 5′SL binds RNA binding protein LARP6 with high affinity and specificity and 5′SL is the only known high affinity target of LARP6. 23,26,28 LARP6 serves as an adapter protein that recruits accessory translational factors to increase translational competency of collagen mRNAs and to couple translation of α1(I) polypeptide to that of α2(I) polypeptide. 6,7 The coordinated synthesis of α1(I) and α2(I) polypeptides increases their local concentration in the endoplasmic reticulum, what accelerates their folding into type I collagen and enables rapid excretion of the heterotrimer into the extracellular matrix.
The clear distinction between profibrotic and constitutive regulation of type I collagen synthesis came from creation of 5′SL knock in mice. 29 In 5′SL knock in mice a mutation was introduced into collagen α1(I) gene, which changed the nucleotides encoding the 5′SL. The mutation did not change the coding region of the gene nor the expression level of the mRNA. Thus, the homozygous knock in mice express collagen α1(I) polypeptide from the mRNA that is not subjected to LARP6-dependent regulation. The 5′SL knock in mice develop normally and have no abnormalities, proving that constitutive collagen synthesis is not compromised. However, when liver fibrosis was induced in these animals it was significantly milder than the fibrosis in control mice, 29 suggesting the importance of LARP6/5′SL mechanism for fibrosis development. The fact that LARP6 binding was not required for constitutive type I collagen expression suggests that expression of type I collagen in fibrosis can be specifically targeted by inhibitors of LARP6 binding.
To find inhibitors of LARP6 binding to 5′SL we have recently performed one high throughput screen that was based on measurement of fluorescence polarization (FP). 30 This screen discovered an inhibitor that was active in suppressing hepatic fibrosis in rats at dose of 1 mg/kg. 31 The success of this effort prompts us to look for additional compounds, hypothesizing that using different screening methods may discover different compounds. In the last decade most new drugs were discovered using phenotypic screens; 32,33 therefore, we decided to develop a phenotypic screening technology for finding LARP6 inhibitors. This work describes development, testing, and application of a technology based on inverted three hybrid system in yeast.
Materials and Methods
Construction of Three Hybrid System Genes
To express fusion protein of Gal4 DNA binding domain and MS2 COAT protein two tandemly arranged copies of bacteriophage MS2 COAT protein were cloned in frame with Gal4 DNA binding domain with myc tag in pGBKT7 vector. This hybrid was designated as MS2COAT-BD. The segment of this vector containing ADH1 promoter, MS2COAT-BD hybrid gene, and TRP1 gene conferring tryptophan auxotrophy was amplified by PCR with primers containing BamHI and XmaI sites and the PCR product was cloned into BamHiI and XmaI sites of pRS314 vector. The pRS314 vector was maintained as episomal in yeast cells used in all experiments by growing cells in medium lacking tryptophan (-TRP).
To express the fusion protein of Gal4 transcription activating domain and LARP6 the truncated LARP6 containing the minimal sequence required for binding 5′SL RNA (LARP6*) 26 was cloned into pACT2 vector (Clontech) in frame with GAL4 activation domain and hemagglutinin (HA) tag. This hybrid was designated as LARP6*-AD. As control, a mutant of LARP6* that cannot bind 5′SL RNA (RAN mutant, described in Ref. 23 ) was cloned the same way and designated RAN-AD. To co-express LARP6*-AD or RAN-AD with the 5′SL RNA an independent gene containing yeast Pol III promoter driving expression of hybrid RNA with 5′SL sequence of human collagen α1(I) gene fused to two MS2 binding motifs (5′SL-MS2 RNA) was inserted between the two NotI sites of pACT2 vector located 3′ to the cloned LARP6*-AD or RAN-AD gene. These constructs express both; LARP6*-AD and 5′SL/MS2 RNA or RAN-AD and 5′SL-MS2 RNA were designated LARP6*-AD/5′SL-MS2 and RAN-AD/5′SL-MS2.
To integrate the LARP6*-AD, RAN-AD, LARP6*-AD/5′SL-MS2, and RAN-AD/5′SL-MS2 into yeast genome the part of the plasmids containing these genes, together with the LEU2 gene of the vector conferring leucine auxotrophy, was amplified by PCR with primers having ApaI and EcoRV sites and the PCR product was cloned into ApaI and SmaI sites of pRS315 vector. pRS315 vectors were linearized with ClaI, transformed into MaV203ΔPS cells, and the integrants were selected by leucine auxotrophy.
Creation and of MaV203ΔPS Host Cells
Mav203 Saccharomyces cerevisiae strain was described before. 34 The genotype of MaV203 cells: MATα, leu2-3, 112, trp1-901, his3Δ200, ade2-101, gal4Δ, gal80Δ, SPAL10::URA3, GAL1::lacZ, HIS3UAS GAL1::HIS3@LYS2, can1R, cyh2R. To delete two major drug resistance genes, PDR5 35 and SNQ2, 36 hygromycin resistance gene was first integrated into the PDR5 locus by homologous recombination. After selection of hygromyin B-resistant clones, SNQ2 gene was deleted by integration of kanamycin resistance gene into the SNQ2 locus and selection by G418. The PDR5/SNQ2 negative strain was termed MaV203ΔPS.
The pRS314 vector expressing MS2COAT-BD gene was transformed into MaV203ΔPS cells and maintained by selecting for tryptophan auxotrophy. pRS315 vectors encoding for LARP6*-AD, LARP6*-AD/5′SL-MS2, RAN-AD, or RAN-AD/5′SL-MS2 were integrated into the LEU2 locus of MaV203ΔPS cells by homologous recombination. The integrands were verified for expression of both, MS2COAT-BD and LARP6*-AD or RAN-AD by western blot.
Western Blots
Yeast cells were disrupted by vortexing with glass beads and lysates were resolved on 10% SDS-PAGE gel and transferred to nitrocellulose membrane. The membrane was probed with anti-HA antibody (detects LARP6*-AD and RAN-AD), with anti-myc antibody (detects MS2COAT-BD) and with anti-phosphoglycerate kinase antibody (PGK1) as loading control. 37
Growth in Multiwall Plates
MaV203ΔPS cells expressing MS2COAT-BD and LARP6*-AD, LARP6*-AD/5′SL-MS2, RAN-AD, or RAN-AD/5′SL-MS2 were seeded at OD600 of 0.05–0.06 in 40 μL per well of 384-well plates (Cat No. 781091; Greiner Bio-one) or 100 μL per well of 96-well plates (Cat No. 167008; Nunc) in liquid medium lacking tryptophan (-TRP) or lacking tryptophan and uracil (-TRP-URA) or lacking tryptophan and containing 0.2% 5-fluoroorotic acid (-TRP +2x5FOA) and OD600 was read after the seeding (day 0). This reading was subtracted as a blank from the subsequent readings, which were taken after 1, 2, and 3 days of incubation at 30°C. When the inhibitory effect of C9 was tested, the compound 31 was added at 0–10 μM to the cells in -TRP +2x5FOA medium at day 0.
Screening of NIH Clinical Collection Library
NIH clinical collection library NCC-003 containing 448 FDA approved drugs was purchased from BioFocus (San Francisco, CA). The drugs were added at final concentrations of 400 μM to MaV203ΔPS cells expressing MS2COAT-BD and LARP6*-AD/5′SL-MS2 and the cells were seeded into -TRP +2xFOA medium in 96-well plates. OD600 was read after 1, 2, and 3 days of incubation at 30°C and OD600 at day 1 was subtracted from OD600 at day 2 to obtained the net growth for day 2 (ΔOD600 day 2) and OD600 at day 2 was subtracted from OD600 at day 3 to obtained the net growth for day 3 (ΔOD600 day 3). The ΔOD600 at day 2 and 3 were plotted for each drug and compared to ΔOD600 of control cells. Control cells received DMSO (vehicle) and their daily growth was used to calculate the mean and standard deviation (SD) and the cutoff value for scoring hits was set as mean+3SD. A drug that showed increase of ΔOD600 at day 2 or ΔOD600 at day 3 by >mean +3SD was considered a hit.
Counter-screening of hits was performed by adding these drugs to cells growing in -TRP-URA medium. A hit that showed decrease of ΔOD600 at day 2 and ΔOD600 at day 3 of >mean −3SD of the control cells was considered a confirmed hit.
FP Assay
FP assay was described before. 30 Briefly, recombinant LARP6* was added in concentrations of 0–0.8 to 1 nM of fluorescein labeled 5′SL RNA (fl-5′SL) (Dharmacon) in 25 μL of 70 nM NaCl, 5 mM MgCl2, 10 nM Tris pH 7.5, and FP measured in 384-well plate (Brand) with Biotek Synergy H1 plate reader. Blank FP of fl-5′SL in buffer was subtracted from the readings with protein.
For testing of the confirmed hits, 0.375 nM LARP6* was bound to 1 nM fl-5′SL, the hit drugs were added at concentrations of 0–200 μM and FP was read.
Results
Creation of the Inverted Three Hybrid System
Yeast three hybrid system was originally described by Sengupta et al. 38 and it has been used to clone and characterize RNA binding proteins. 39 The system consists of two hybrid proteins and one hybrid RNA. In our modification of the system the hybrid RNA contained 5′SL sequence of human collagen α1(I) mRNA fused to the two MS2 binding sequences (5′SL-MS2 RNA). MS2COAT is RNA binding protein of Escherichia coli bacteriophage that binds MS2 RNA sequence with high affinity. 40 The first hybrid protein contained tandem repeat of MS2COAT protein fused to the GAL4 DNA binding domain of yeast transcription activator GAL4 (this hybrid protein was termed MS2COAT-BD), so that the hybrid molecule can bind simultaneously to the promoter containing GAL4 sequence and to the RNA that has two MS2 motifs.
The second hybrid protein contained human LARP6 with minimal sequence required to bind 5′SL (LARP6*) or the RAN mutant of LARP6*, which were fused to the GAL4 transcription activation domain (constructs LARP6*-AD and RAN-AD). The RAN mutant cannot bind 5′SL RNA and was used as negative control. 23 To co-express 5′SL-MS2 RNA with LARP6*-AD or RAN-AD a cassette containing the RNA gene was cloned downstream of LARP6*-AD or RAN-AD genes as independent transcription unit. Thus, constructs LARP6*-AD/5′SL-MS2 and RAN-AD/5′SL-MS2 were created, which express both, the protein and RNA (Fig. 1). When LARP6*-AD, MS2COAT-BD, and 5′SL-MS2 RNA are co-expressed, MS2COAT-BD, having DNA binding domain, is brought to proximity of LARP6*-AD, having transcription activation domain, via the 5′SL-MS2 RNA bridge (Fig. 1A).
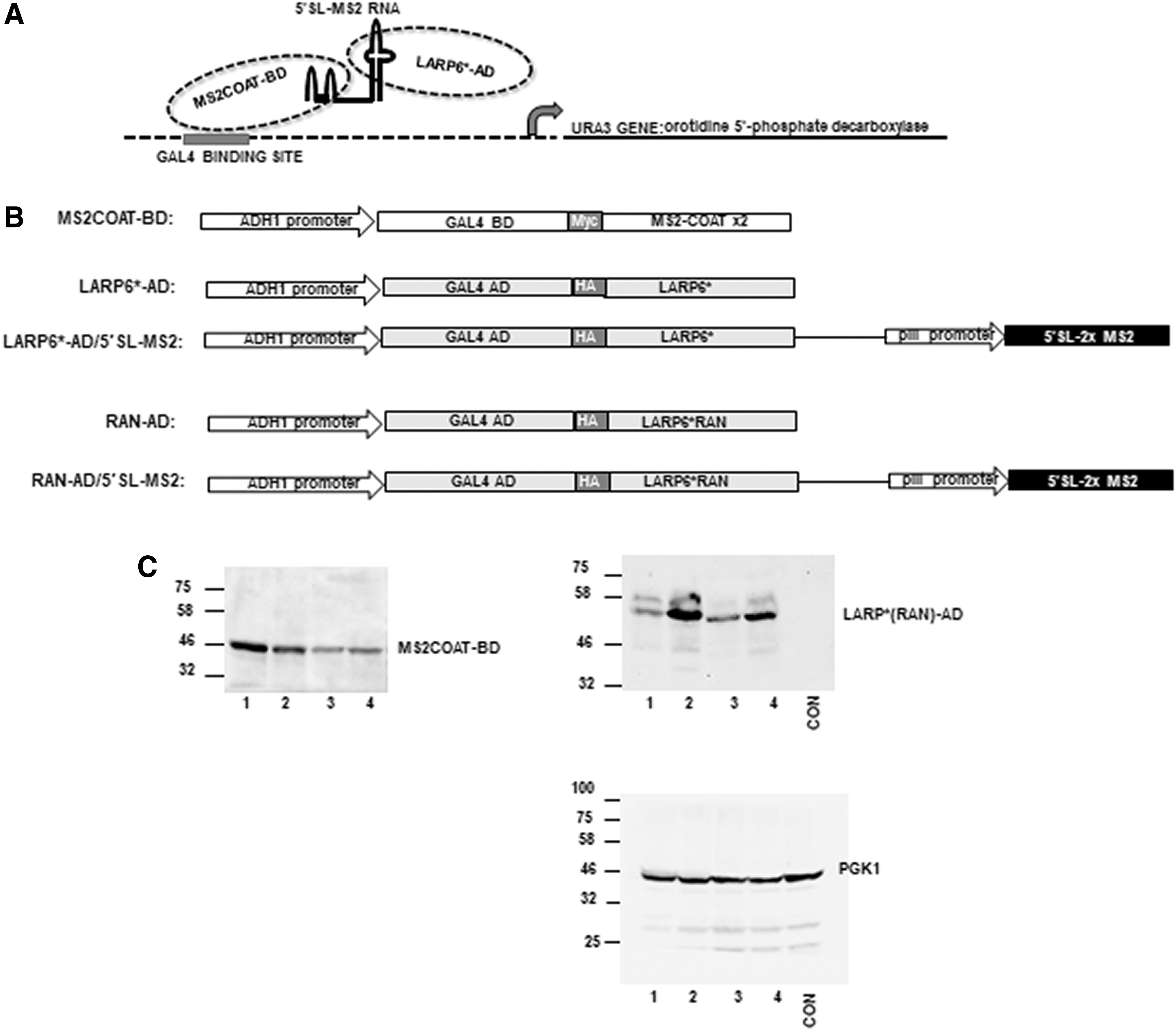
Principle and components of inverted yeast three hybrid system.
This complex can recognize promoter that contains GAL4 recognition sequence and activate transcription of the downstream gene. In our case the downstream gene was ura3, which encodes for orotidine 5′-phosphate decarboxylase. This enzyme allows yeast cells to grow in synthetic medium lacking uracil and inhibits the growth in synthetic medium containing 5-fluoroorotic acid (5FOA). Thus, formation of the 3-hybrid complex can be detected by the growth in -URA medium and lack of growth in 2x5FOA medium. If the interaction between LARP6* and 5′SL is blocked, the three hybrid bridge cannot form and ura3 gene is not transcribed. Without orotidine 5′-phosphate decarboxylase cells cannot grow in the medium lacking uracil, but will grow in medium containing 5-FOA. Thus, the system can be used to positively select for inhibitors of 5′SL/LARP6 interaction by growing cells in the medium containing 5-FOA and negatively select by growing in medium without uracil. In preliminary experiments we have established that 0.2% 5FOA is the optimal concentration for the growth selection.
The constructs expressing the components of the system are shown in Figure 1B. MS2COAT-BD construct was maintained in yeast as an episomal plasmid conferring tryptophan auxothrophy. LARP6*-AD, RAN-AD, LARP6*-AD/5′SL-MS2, and RAN-AD/5′SL-MS2 constructs were integrated as single copy genes into Leu2 genomic locus of yeast cells harboring the MS2COAT-BD episomal plasmid. Thus, cells expressing four combinations of genes were created: (1) cells expressing MS2COAT-BD+LARP6*-AD/5′SL-MS2; in these cells the three hybrid complex forms and URA3 gene is active, (2) cells expressing MS2COAT-BD+LARP6*-AD; in these cells there is no RNA and URA3 gene is silent, (3) cells expressing MS2COAT-BD+RAN-AD/5′SL-MS2; in these cells there is no binding of RAN-AD to 5′SL-MS2 RNA because the mutant cannot recognize 5′SL and URA3 gene is silent, (4) cells expressing MS2COAT-BD+RAN-AD; in these cells mutant protein is expressed and there is also no RNA, so URA3 gene is silent. The latter three combinations served as negative controls to test the specificity and tight regulation of the system.
The host cells were yeast strain MAV203, which has GAL4 promoter inserted upstream of the ura3 gene. 34 To make these cells suitable for drug screening we deleted two genes encoding for drug exporters pdr5 35 and snq2, 36 creating the MAV203ΔPS strain. Thus, MAV203ΔPS strain accumulates chemical compounds to high levels intracellularly.
Expression of three hybrid proteins in MAV203ΔPS is shown in Figure 1C. MS2COAT-BD was expressed from episomal plasmid (left panel) and LARP6*-AD and RAN-AD proteins were expressed from single copy genes integrated into genomic DNA of MAV203ΔPS cells harboring the MS2COAT-BD plasmid (right panel).
Specificity of the Inverted Three Hybrid System
Figure 2 shows the growth of MAV203ΔPS cells expressing different combinations of three hybrid components when plated on various selection plates. Tryptophan was omitted in all plates to assure retention of the MS2COAT-BD plasmid. Ten-fold serial dilutions of cells were spotted on a plate lacking tryptophan (-TRP, left panel), a plate lacking tryptophan and uracil (-TRP-URA, middle panel), and a plate lacking tryptophan and containing 0.2% 5FOA (-TRP +2xFOA, right panel). The cells grew equally well when there was no selection for the formation of three hybrid complex (-TRP), however, when plated on -TRP-URA plates, only the functional complex (MS2COAT-BD+LARP6*-AD/5′SL-MS2) supported cell growth. On -TRP +2x5FOA plates the MS2COAT-BD+LARP6*-AD/5′SL-MS2 complex suppressed the growth, while all other combinations allowed for growth. This result verified that the phenotypes associated with productive assembly of the three hybrid complex are growth in -URA medium and lack of growth in 2x5FOA medium.

Growth of MAV203ΔPS strain expressing different combinations of three hybrid components on various selective plates. Tryptophane (TRP) was omitted from all plates to assure maintenance of MS2COAT-BD episomal vector. Ten-fold serial dilutions of MAV203ΔPS cells expressing the indicated genes were plated on -TRP plate, on -TRP-URA plate and on -TRP +2x5FOA plate. Incubation on -TRP and on -TRP-URA plate was for 2 days and on -TRP +2x5FOA plate for 3 days.
Inverted Three Hybrid System as Technology for High Throughput Screening
The screening for inhibitors of LARP6 binding to 5′SL RNA is based on ability of chemical compounds to disrupt the three hybrid complex by dissociating LARP6* from the 5′SL-MS2 RNA. As a result, the cells will gain ability to grow in 2x5FOA medium (positive selection) and loose ability to grow in -URA medium (negative selection). To demonstrate the performance of the system in a high throughput setting we measured the growth rates as OD600 after seeding MAV203ΔPS cells into nonselective or selective liquid media in 384-well plates (Fig. 3). The measurements were taken for three consecutive days, incubations longer than 3 days resulted in evaporative loss of the medium. At day 0 the cells were seeded at OD600 of ∼0.06 and the OD600 of each well was measured. The OD600 at day 0 served as a blank and was subtracted from the readings obtained on the subsequent days. In nonselective medium (-TRP, Fig. 3A) all cells grew equally well, reaching saturation after 1 day.
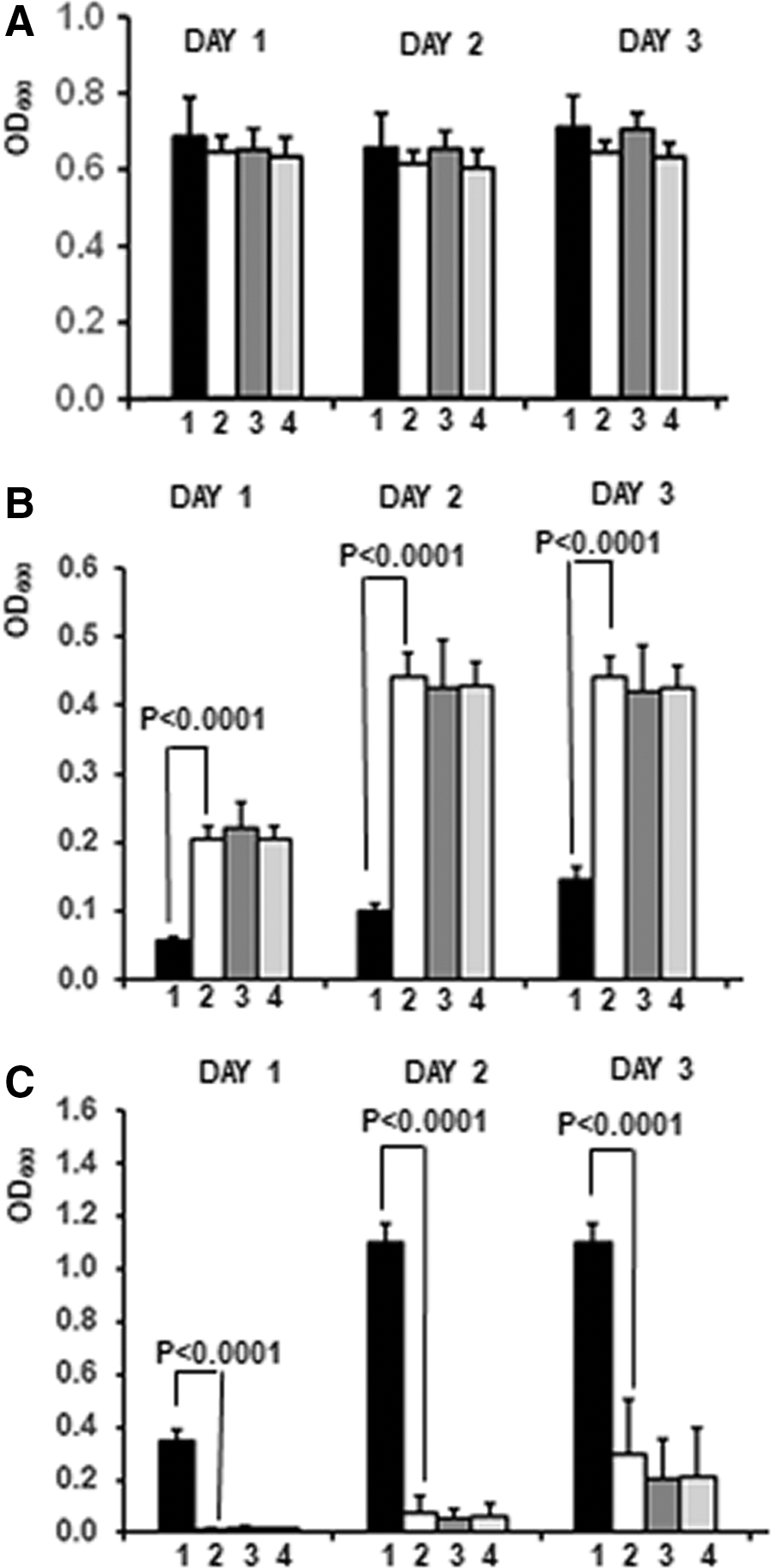
Growth of MAV203ΔPS strain expressing different combinations of three hybrid genes in liquid media.
In medium containing 2xFOA the cells expressing MS2COAT-BD+LARP6*-AD/5′SL-MS2 showed minimal growth (Fig. 3B, black bars), while the cells expressing MS2COAT-BD+LARP6*, MS2COAT-BD+RAN-AD/5′SL-MS2, and MS2COAT-BD+RAN-AD grew much faster and reached saturation by day 2 (open and gray bars). In contrast, when the cells were incubated in liquid medium lacking uracil only cells expressing MS2COAT-BD+LARP6-AD*/5′SL-MS2 showed robust growth, while cells with other gene combination grew poorly (Fig. 3C).
These results indicate that a positive phenotypic screen for LARP6 inhibitors can be performed using MAV203ΔPS cells expressing MS2COAT-BD+LARP6*-AD/5′SL-MS2 growing in 0.2% 5FOA medium, while a counter-screen can be performed by measuring inhibition of growth in -URA medium.
Z′-Factor of the Assay
Z′-factor is a measure of how well an assay will perform in high throughput screenings. To calculate Z′-factor positive and negative controls and SDs of their readings are needed. In our system, the growth rate of cells expressing MS2COAT-BD+LARP6*-AD in 2x5FOA medium is used as the positive control. In these cells there is no expression of 5′SL-MS2 RNA and the interaction of LARP6 and 5′SL is nonexistent, therefore, they reflect the maximal effect that can be achieved by an inhibitor. The cells expressing MS2COAT-BD+LARP6*-AD/5′SL-MS2 serve as the negative control, because there is unrestricted binding of LARP6 in these cells. By measuring the growth rate of the positive and negative control in 2x5FOA medium in 384-well plates we calculated the Z′-factor. 41 Table 1 shows four independent determinations of the Z′-factor, which was calculated at day 1, 2, or 3 in culture. Average Z′-factor of 0.6 was obtained at day 2 and of 0.535 at day 3. These Z′-factors suggest that the assay performance is satisfactory for high throughput applications and that the relevant measurements should be taken at day 2 and 3. These data also point to a good day to day reproducibility of the assay.
Z-values of the Assay Calculated in Four Independent Experiments by Measuring OD600 at Day 1, 2, and 3
Validation of the Technology Using Previously Discovered LARP6 Inhibitor
We have previously described discovery and antifibrotic activity of one LARP6 inhibitor. 31 This inhibitor, C9, prevented progression of hepatic fibrosis when administered at 1 mg/kg into experimental animals, while in vitro it reduced binding of LARP6 to 5′SL with IC50 of 13 μM. Therefore, we used this compound to test the inverted three hybrid system. We added C9 to the MAV203ΔPS expressing MS2COAT-BD+LARP6*-AD/5′SL-MS2 and measured the effect on growth in nonselective medium and in 2x5FOA medium. Figure 4A shows that C9 did not have significant effect on growth of cells in nonselective medium in concentrations up to 10 μM. However, in 2x5FOA medium the compound increased the growth rate of cells in concentrations of 1–10 μM. Significant and concentration-dependent increase in cell density was measured at day 1, 2, and 3 (Fig. 4B), suggesting that the system can detect effect of an inhibitor at concentrations lower than its IC50 in vitro.
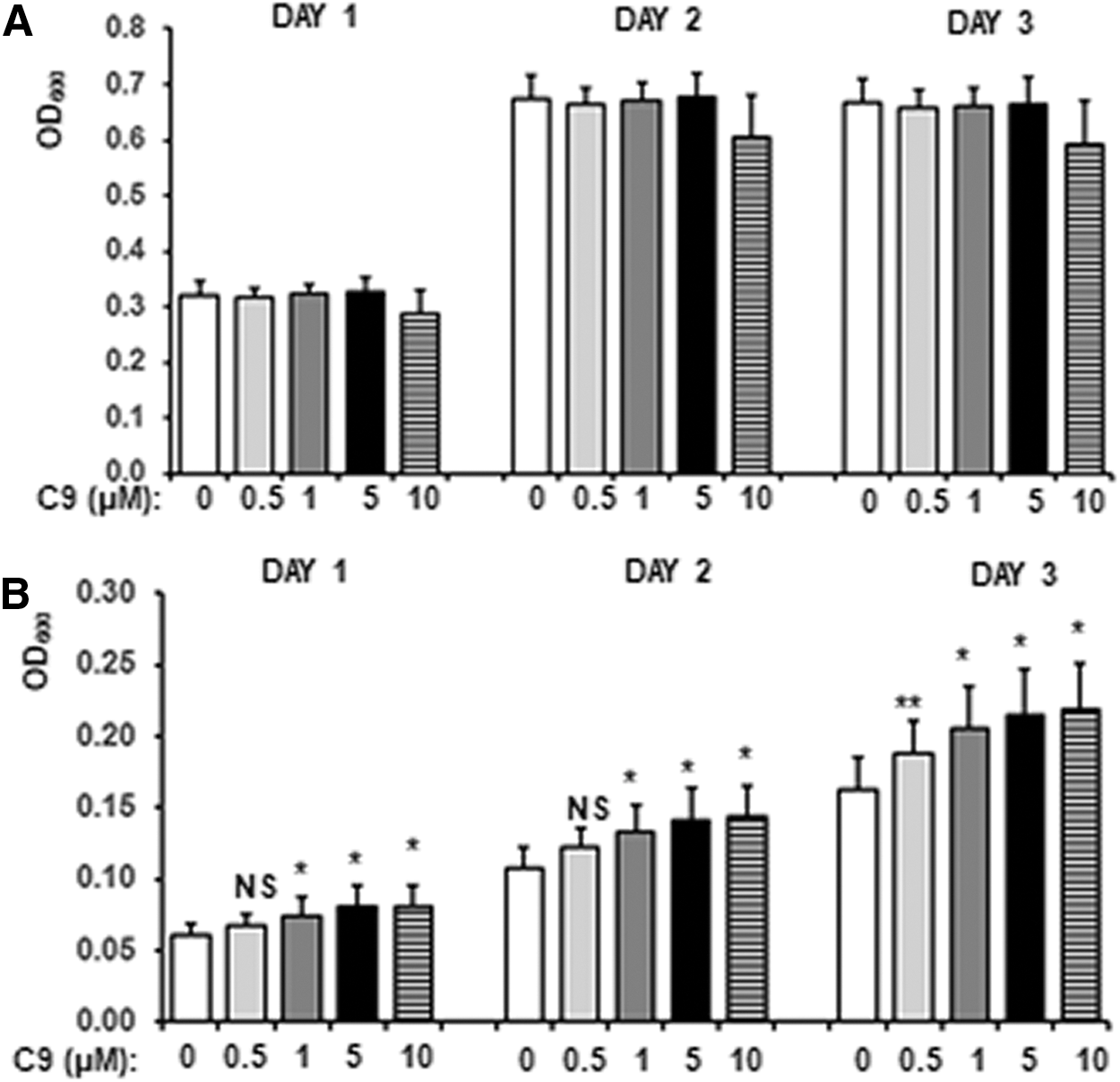
Effect of previously characterized LARP6 inhibitor, C9.
Application of the Technology in Screening of Chemical Libraries
As a proof of principle that the technology can be applied to find novel LARP6 inhibitors we performed a screen of NIH Clinical Collection library NCC-003 containing 448 FDA approved drugs. This screen was performed in 96-well plates, so we first assessed whether the growth in 96-well plates follows the same kinetics as in 384-well plates. Figure 5 shows measurement of OD600 in 96-well plates, where 100 μL of cell suspension was distributed per well. At day 0 the cells were seeded at OD600 of ∼0.06, the OD600 was read and subtracted as a blank from the readings on subsequent days. In nonselective medium the cultures reached saturation after 1 day (Fig. 5A). In 2x5FOA medium the MS2COAT-BD+LARP6*-AD/5′SL-MS2 expressing cells failed to grow, while the cells expressing MS2COAT-BD+LARP6*-AD, MS2COAT-BD+RAN-AD/5′SL-MS2, and MS2COAT-BD+RAN-AD reached saturation by day 3 (Fig. 5B). The Z′-factor estimated in 96-well plates at day 2 and 3 was >0.6. Thus, the system has similar performance regardless of the plate format used.

Growth in 96-well plates.
Because the tested drugs were in DMSO, we first assessed the DMSO concentrations that can be tolerated by MAV203ΔPS cells. DMSO concentrations of up to 5% had no major effect on cell viability (Supplementary Fig. S1), which allowed us to use the drugs from the library at concentration of 400 μM (4% final DMSO concentration). After seeding MS2COAT-BD+LARP6*-AD/5′SL-MS2 expressing cells in -TRP +2x5FOA medium, the drugs were added and OD600 was measured daily. In this screen we calculated the daily gain in cell density by subtracting the OD600 measured at day 1 from the OD600 measured at day 2 (ΔOD600 day 2) and the OD600 at day 2 from the OD600 at day 3 (ΔOD600 day 3). In this way we obtained two values as a readout of the ability of drugs to dissociate LARP6*. The wells where no drugs were added served as controls (16/plate). From these controls we calculated the mean and SD and any drug that showed ΔOD600 increase of >3SD of the controls at either day 2 or 3 was considered a hit.
The results of the screen are shown in Supplementary Figures S2–S7. By these criteria we detected 8 hits at day 2 and 15 additional hits at day 3. Out of 448 compounds tested this represents a hit rate of 5%. This relatively high hit rate required counter-screening, which was done by seeding the cells in -TRP-URA medium; in this medium the hits were validated if they could decrease the daily growth by >mean −3SD of the controls without drugs. Two drugs passed the counter-screen, indicating a hit rate of 0.4%. The two drugs were adenosine receptor antagonist CGS15943 (ARA) and rifaximin (RIF). Figure 6 shows the results of the screen and counter-screen of these two drugs. Black bars show daily growth gain in the 2x5FOA medium compared to the controls, with the cutoff value of the assay (CON MEAN+3SD) and open bars show growth inhibition in -URA medium with the cutoff (CON MEAN−3SD).
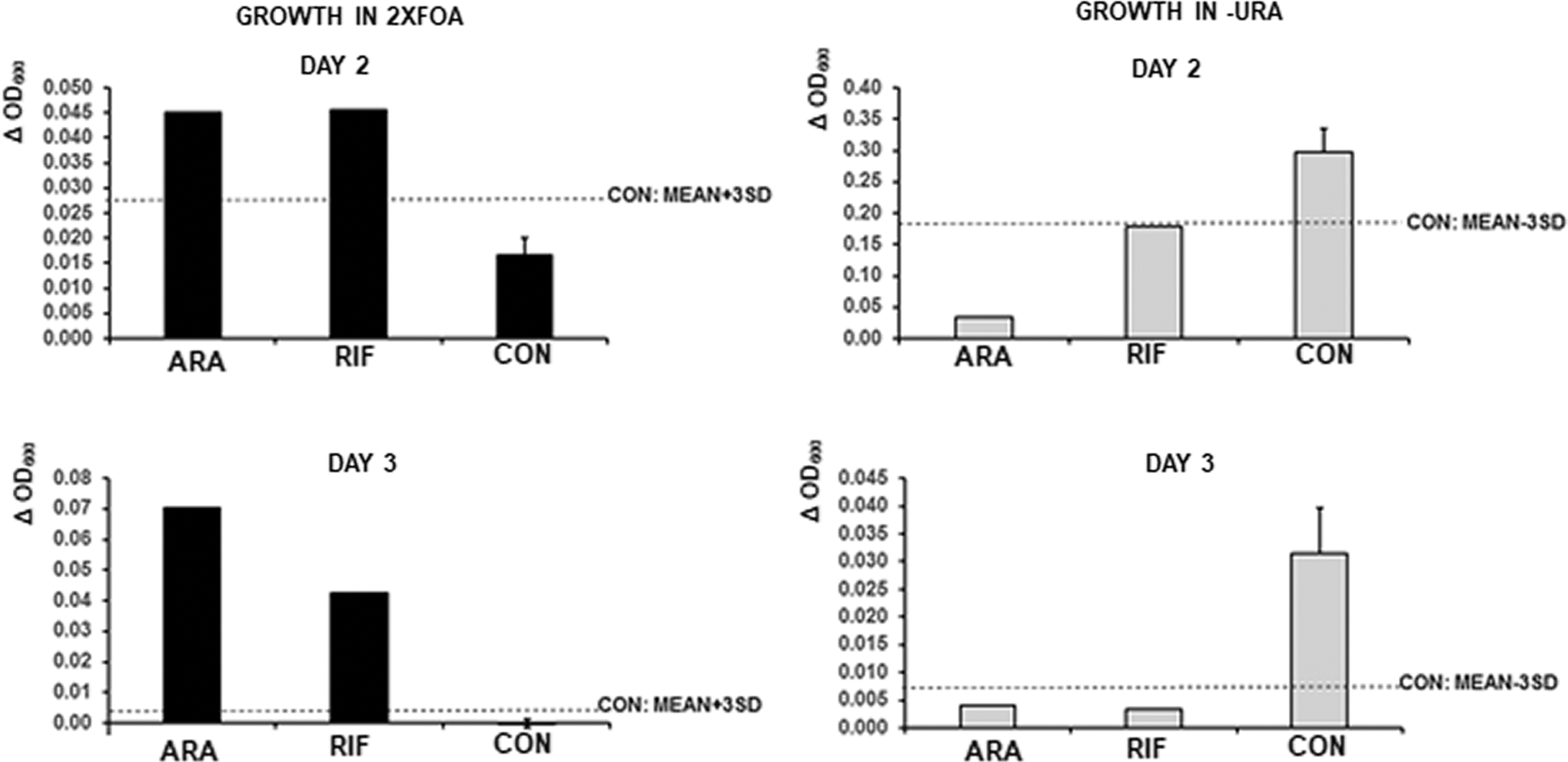
Results of the two positive drugs from the screen of NIH Clinical Collection of FDA approved drugs. Adenosine receptor antagonist CGS15943 (ARA) and rifaximin (RIF) were added at 400 μM to MAV203ΔPC cells expressing MS2COAT-BD+LARP6*-AD/5′SL-MS2 genes and grown in -TRP +2x5FOA medium (black bars) or in -TRP-URA medium (gray bars). OD600 at day 1 was subtracted from OD600 at day 2 (ΔOD600, day 2, upper panels) and OD600 at day 2 was subtracted from OD600 at day 3 (ΔOD600, day 3, lower panels). CON, cells receiving DMSO. Dotted line: mean of controls +3SD (black bars) or mean of controls-3SD (gray bars).
Validation of the Hits by a Biochemical Method
Small number of hits obtained after the counter-screen can be validated using biochemical methods. We have described the FP method to measure LARP6 binding to the 5′SL RNA, 30 which we used for the validation. When increased amounts of recombinant LARP6* are added to a constant amount of fluorescently labeled 5′SL RNA (fl-5′SL) in vitro, the concentration-dependent increase in FP is seen (Fig. 7A). The increase reflects formation of LARP6*/fl-5′SL RNA complex until the saturation is reached. When LARP6* is added at sub-saturating amounts to fl-5′SL RNA and the compounds are added to the reaction, inhibition of LARP6*/fl-5′SL complex formation can be assessed as decrease in FP. So, we added the two hits in increasing concentrations to the LARP6*/fl-5′SL binding reaction and measured the FP. CGS15943 (ARA) showed no effect on binding, however, rifaximin decreased the FP in a concentration-dependent manner (Fig. 7B), suggesting that this drug can inhibit association of LARP6* and fl-5′SL RNA.
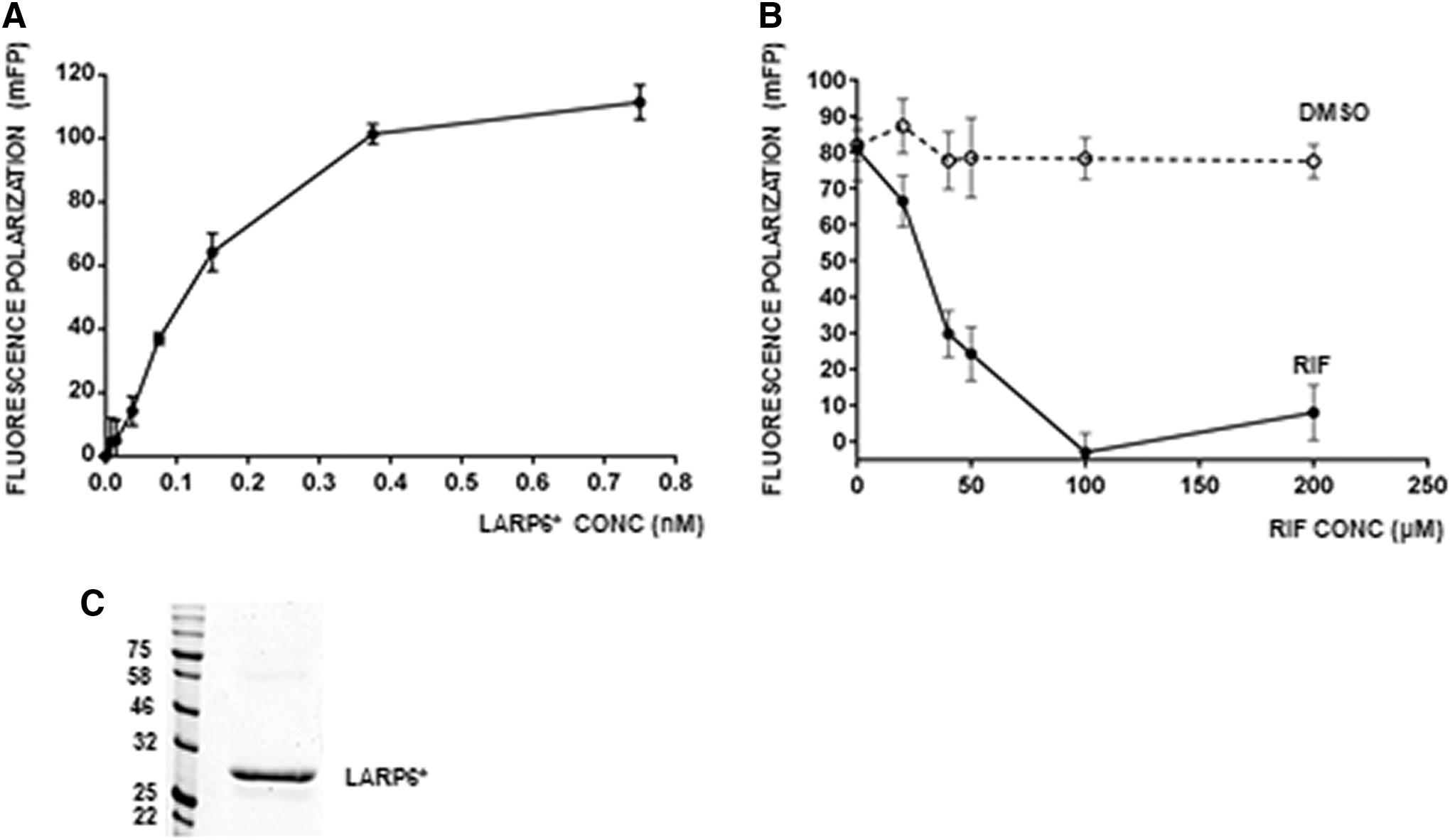
Inhibition of LARP6* binding by rifaximin measured by FP.
Rifaximin itself has no fluorescence at the excitation/emission wavelengths used for the FP measurements; therefore, it did not interfere with the readings and the decrease in FP was attributed to the inhibition of LARP6*/fl-5′SL complex formation. From the inhibition curve in Figure 7B it can be seen that 50% inhibition was achieved at ∼40 μM of rifaximin. The purity of LARP6* used in these experiments is shown in Figure 7C. Although rifaximin is not intended to be used as antifibrotic drug, the experiment demonstrated the proof of principle that the screen based on inverted yeast three hybrid technology can identify compounds with the ability to inhibit LARP6 binding to 5′SL RNA.
Discussion
Fibrosis is chronic condition resulting from excessive synthesis of extracellular matrix composed predominantly of type I collagen. Although fibrosis affects millions of people there are no approved antifibrotic drugs. As a chronic disease fibrosis requires long-term treatment, therefore, antifibrotic drugs must have minimal side effects, should specifically target type I collagen, and be affordable. High level of type I collagen biosynthesis in fibrosis is supported by binding of protein LARP6 to the unique sequence element, 5′SL, found in mRNAs encoding for type I collagen. 5 –7,9,22,23,42 This mechanism is specific for regulation of type I collagen expression in fibrosis, but is dispensable for constitutive expression. 29 Therefore, small molecule inhibitors of LARP6 binding can be antifibrotic drugs with desirable properties. We have recently described identification and characterization of one inhibitor of LARP6 binding to 5′SL; this compound was active in suppressing hepatic fibrosis when administered at 1 mg/kg in experimental animals. This increased the interest in finding additional inhibitors, but using different methods.
The first LARP6 inhibitor was discovered by in vitro method based on FP. 30,31 However, in the last decade most new drugs were discovered using phenotypic screens. 33,43,44 This prompts us to devise the phenotypic screen described here, as a technology to discover additional LARP6 inhibitors. We show that (1) the system specifically reports binding of LARP6 in yeast cells, (2) the readout is simple and has good Z′-factor for high throughput screens, (3) the screens can be performed in plates of different formats, (4) the counter-screen can be easily done by switching to a different synthetic medium, (5) as a proof of principle, one FDA approved drug was identified that has ability to prevent LARP6 binding to 5′SL.
To our knowledge this is the first report that yeast three hybrid system can be applied as high throughput drug screening method. The main reason yeast three hybrid system was used as the platform is because the system is tightly regulated. It allows on/off switching of the URA3 gene with sufficient precision to obtain the Z′-factor acceptable for high throughput screening. The other advantage is that the system is easy to manipulate and score. We have just inverted the system so that dissociation of LARP6 from 5′SL RNA can be scored as a positive outcome. The disadvantage of the system is that some drugs may poorly penetrate or accumulate in the yeast cells, what we tried to alleviate by deleting two major drug efflux genes. However, other drug efflux systems also exist, what may limit the performance of the system. Using high drug concentration in the screen may overcome this problem (we used 400 μM) and also eliminate potentially toxic compounds. There is no equivalent system developed for mammalian cells, probably because it is difficult to tightly regulate an effector gene that stimulates cell growth.
The basis for our technology is regulation of ura3 gene expression in S. cerevisiae by binding of LARP6 to 5′SL RNA and formation of the three hybrid complex (Fig. 1A), which trans-activates the URA3 promoter. This allows for positive selection of compounds that can dissociate LARP6 by growth in 5FOA medium. The URA3 silencing phenotype can be counter-screened by negative selection by simply switching the growth to -URA medium (Figs. 3 and 5). In our screen this eliminated the majority of hits identified using the positive selection. To make the yeast cells suitable for drug screening we deleted two genes encoding for drug exporters, pdr5 and snq2. 35,36 These two proteins are responsible for majority of drug efflux from S. cerevisiae cells, so their deletion increases drug accumulation in the cells. 45
We performed one screen using small number of FDA approved drugs as a proof of principle that it is possible to identify chemical compounds that can interfere with LARP6 binding using the technology. We identified one drug, rifaximin, that has the ability to interfere with LARP6 binding to 5′SL in vitro (Fig. 7). Rifaximin is an antibiotic that binds β-subunit of bacterial RNA polymerase and blocks transcription. 46 It is used to treat GI disorders and hepatic encephalopathy. It was active in μM concentrations to dissociate LARP6 in vitro, what may be its unrecognized property. We do not suggest that rifaximin should be pursued as antifibrotic drug, but report this finding only as an example that it is possible to identify novel LARP6 inhibitors using our technology. This is further underscored by the fact that one already discovered LARP6 inhibitor tested positive in the assay (Fig. 4). The initial screen resulted in relatively high number of hits, but the counter-screen dramatically reduced their number and we estimated that it will be >0.5% after the counter-screening. This would allow use of biochemical or other methods for their verification.
There are two obvious sources of false positive hits in our system; the compounds that interfere with MS2COAT-BD binding to MS2 sequence and the compounds that inhibit enzymatic activity of the URA3 gene product, orotidine 5′-phosphate decarboxylase. Such inhibitors would give the same phenotype as the compounds that dissociate LARP6. This is illustrated by identification of CGS15943 (ARA) drug in our screen (Fig. 6), which had no effect on LARP6 binding, but had an effect on the growth phenotype of MAV203ΔPS cells.
In conclusion, the technology has been described that allows for screening of large chemical libraries to find inhibitors of LARP6 binding. These inhibitors can be further evaluated and developed as potential antifibrotic drugs. The method is simple, requiring measuring of OD600 on day 2 and 3, has good Z′ factor, can be adapted to different plate formats, and produces manageable number of hits to be further evaluated.
Footnotes
Disclosure Statement
The authors disclose that they have no conflict of interest and that they are not associated with any commercial entities.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7