Algar WR, Hildebrandt N, Vogel SS, Medintz IL. FRET as a biomolecular research tool—understanding its potential while avoiding pitfalls. Nat Methods 2019;16:815–829.
Abstract: The applications of Förster resonance energy transfer (FRET) grow with each year. However, different FRET techniques are not applied consistently, nor are results uniformly presented, which makes implementing and reproducing FRET experiments challenging. We discuss important considerations for designing and evaluating ensemble FRET experiments. Alongside a primer on FRET basics, we provide guidelines for making experimental design choices such as the donor–acceptor pair, instrumentation and labeling chemistries; selecting control experiments to unambiguously demonstrate FRET and validate that the experiments provide meaningful data about the biomolecular process in question; analyzing raw data and assessing the results; and reporting data and experimental details in a manner that easily allows for reproducibility. Some considerations are also given for FRET assays and FRET imaging, especially with fluorescent proteins. Our goal is to motivate and empower all biologists to consider FRET for the powerful research tool it can be.
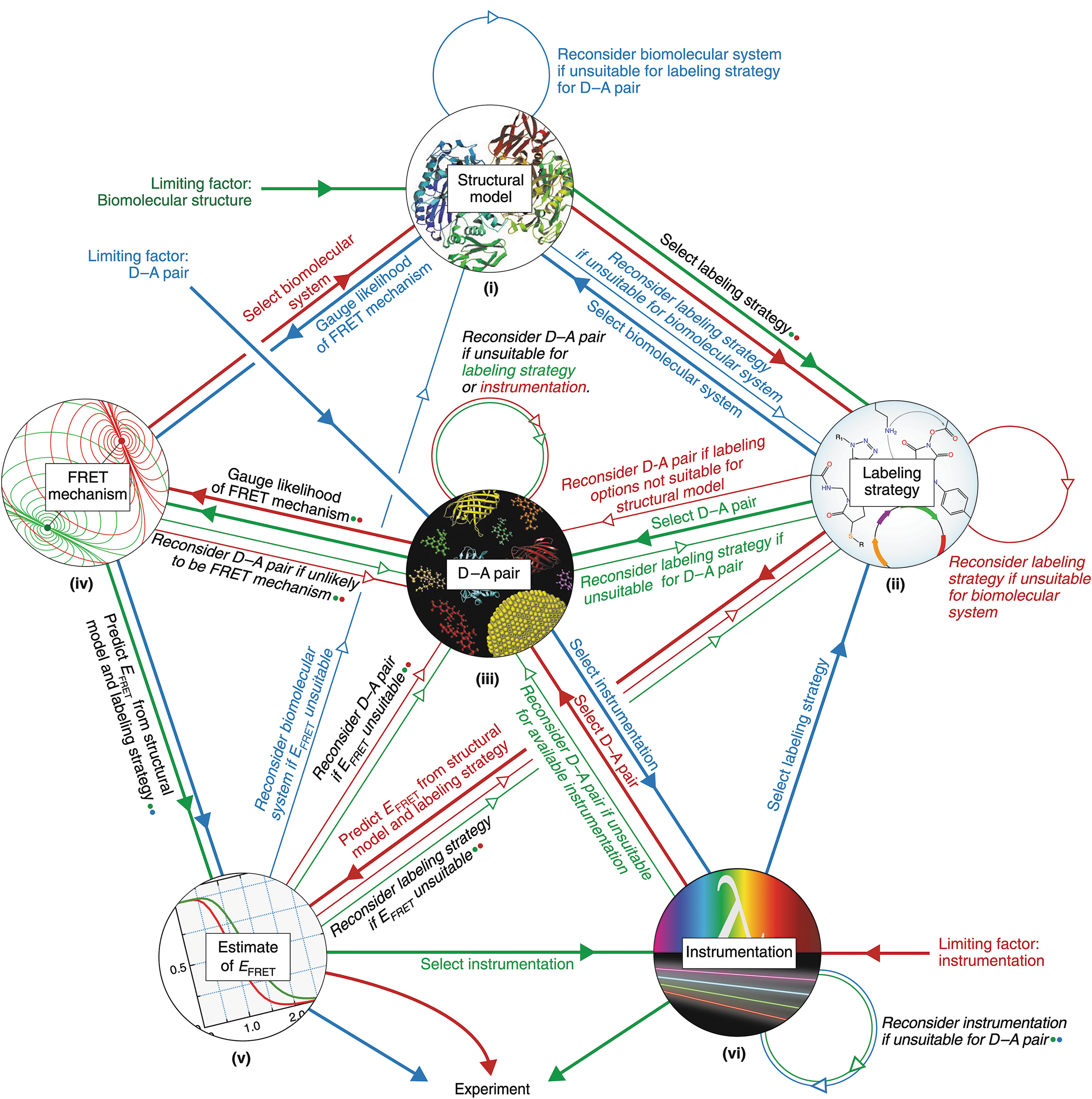
Commentary:One of the most common biochemical assay formats used in compound hit discovery is Förster resonance energy transfer (FRET). In particular, time-resolved FRET (TR-FRET) employing lanthanide donors such as terbium or europium is well utilized. As noted in the paper, the number of publications utilizing FRET to study biology (either biochemical or cellular assays) has risen by more than 10-fold over the last 20 years, with around 2,000 papers noting FRET assays in 2018. This paper provides a comprehensive review and guidance on the development and use of FRET assays for a variety of applications. The paper also lists a good deal of online resources and references to help with choosing suitable FRET pairs for an assay. The supplemental material includes an in-depth description of FRET theory and experimental design, as well as additional resources such as a table of FRET reagents and their properties. A flow chart to design FRET assays is presented (seefigure), as well as a description of the controls needed to determine if FRET is occurring in either biochemical assays or cell-based imaging experiments. This manuscript should be a great resource for those developing or trouble-shooting FRET assays. Contributed by Doug Auld.
Flowchart for designing a FRET system. Diagram summarizing the stepwise and iterative process of considering the multiple factors that contribute to the viability of a FRET system. It also shows suggested starting points depending on the principal factor limiting the design possibilities. The paths from different starting points are color-coded. Thick lines and solid arrows indicate a step forward after a factor is satisfactorily addressed. Thin lines and open arrows indicate reconsideration of how to address a factor. Alternate paths between the factors are also viable.
Abstract: The pharmaceutical and chemical industries depend on additives to protect enzymes and other proteins against stresses that accompany their manufacture, transport, and storage. Common stresses include vacuum-drying, freeze-thawing, and freeze-drying. The additives include sugars, compatible osmolytes, amino acids, synthetic polymers, and both globular and disordered proteins. Scores of studies have been published on protection, but the data have never been analyzed systematically. To spur efforts to understand the sources of protection and ultimately develop more effective formulations, we review ideas about the mechanisms of protection, survey the literature searching for patterns of protection, and then compare the ideas to the data.
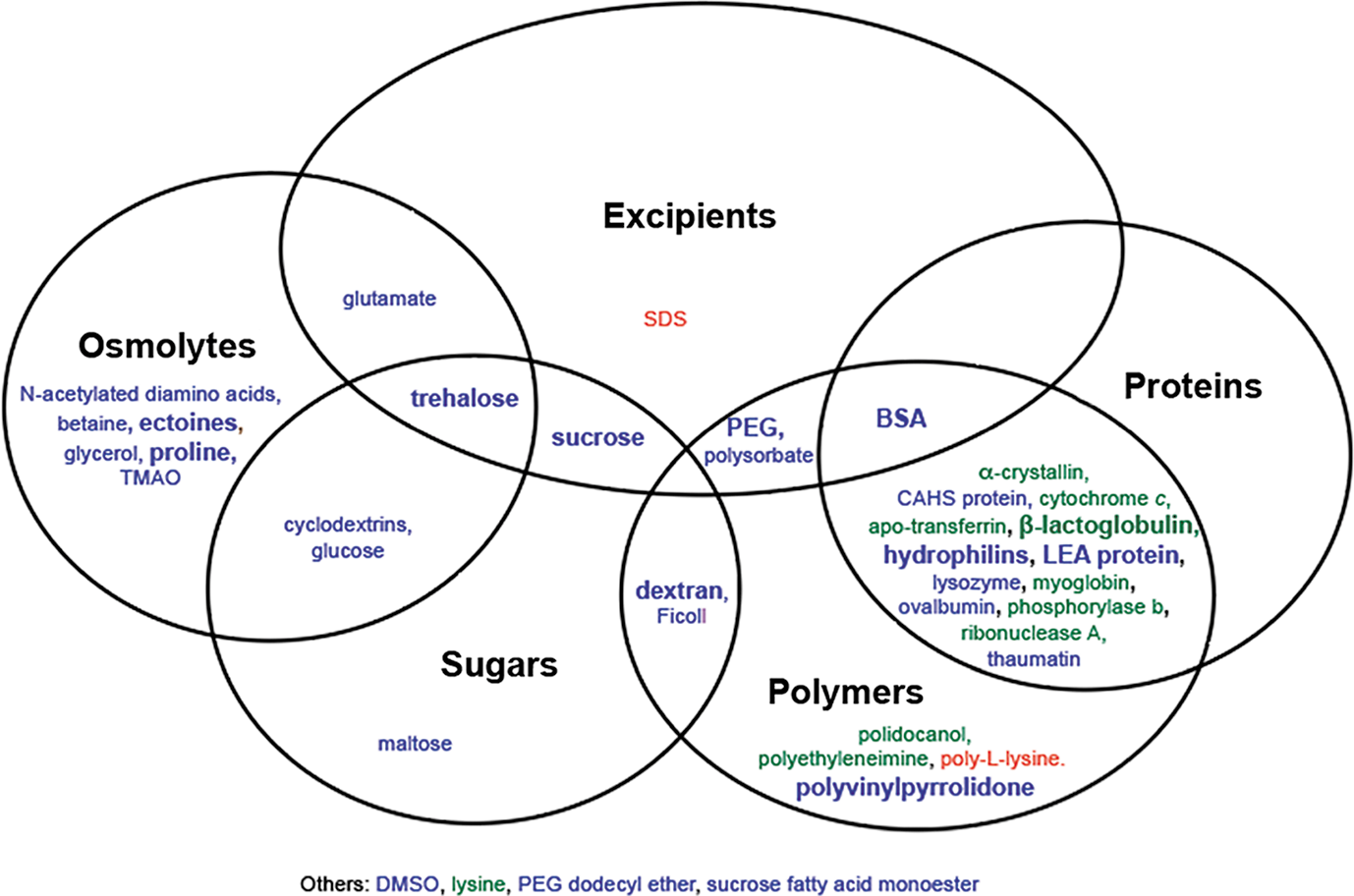
Commentary:Maintaining high-quality performance of proteins during storage is a primary concern for a variety of applications, including high-throughput screening of target proteins. Determining how tolerant a stock solution of protein is to freeze–thaw cycles is critical in determining the optimal aliquot size and planning to support a program that may last for months. This paper provides a critical review and analysis of additives that have been reported to protect proteins against vacuum-drying (desiccation), the freeze–thaw process, and freeze-drying (lyophilization). The supplemental material includes a table with a list of additives reported to protect from freeze–thaw cycles and their effects. Additives that have been reported to show full, partial, or no protection are described in the paper (seefigure). One point is that certain additives may only show partial protection, and oftentimes reports differ on whether a particular additive shows protection. The contradictory reports on the effect of an additive are likely due to factors such as the sensitivity of the tested protein to stress, the concentration of additive used not optimal, or an influence due to other buffer components that differed between the studies. The paper includes an analysis of sugars such as trehalose, natural products such as ectoline, and intrinsically disorder proteins such as late embryogenesis abundant (LEA) protein. The mechanism of protection is unclear at the moment, and more detailed systematic studies on single agents alone and in combination are needed. This paper will be a good reference for researchers exploring improved methods aimed at maintaining high-quality protein preparations. Contributed by Doug Auld.
Venn diagram for protection from freeze–thaw stress. Additives in blue have at least one report of full protection. Additives in green have no reports of full protection but at least one report of partial protection. Additives in red have no reports of protection. A bold font represents two or more reports. DMSO, dimethyl sulfoxide; SDS, sodium dodecyl sulfate.
From Antibody to a Small Molecule
Melgar K, et al. Overcoming adaptive therapy resistance in AML by targeting immune response pathways. Sci Transl Med 2019;11(508):eaaw8828.
Abstract: Targeted inhibitors to oncogenic kinases demonstrate encouraging clinical responses early in the treatment course; however, most patients will relapse because of target-dependent mechanisms that mitigate enzyme-inhibitor binding or through target-independent mechanisms, such as alternate activation of survival and proliferation pathways, known as adaptive resistance. Here, we describe mechanisms of adaptive resistance in FMS-like receptor tyrosine kinase (FLT3)–mutant acute myeloid leukemia (AML) by examining integrative in-cell kinase and gene regulatory network responses after oncogenic signaling blockade by FLT3 inhibitors (FLT3i). We identified activation of innate immune stress response pathways after treatment of FLT3-mutant AML cells with FLT3i and showed that innate immune pathway activation via the interleukin-1 receptor–associated kinase 1 and 4 (IRAK1/4) complex contributes to adaptive resistance in FLT3-mutant AML cells. To overcome this adaptive resistance mechanism, we developed a small molecule that simultaneously inhibits FLT3 and IRAK1/4 kinases. The multikinase FLT3-IRAK1/4 inhibitor eliminated adaptively resistant FLT3-mutant AML cells in vitro and in vivo and displayed superior efficacy as compared to current targeted FLT3 therapies. These findings uncover a polypharmacologic strategy for overcoming adaptive resistance to therapy in AML by targeting immune stress response pathways.
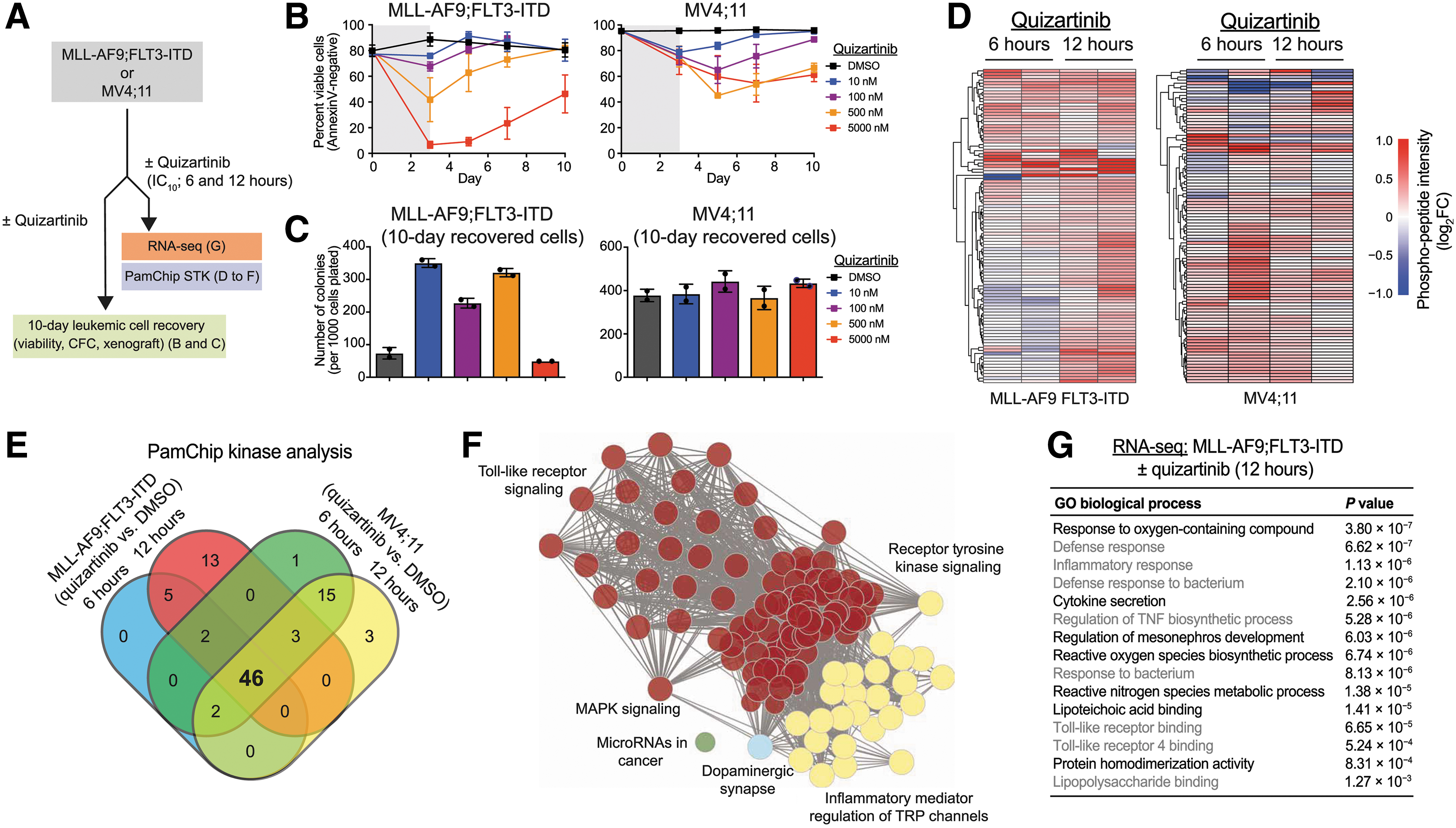
Commentary:Single-target-based therapies in oncology have proliferated during the past few decades, but as they become more widely used, the examples of tumor adaptation and resistance to treatment have also become prominent. As a proof of principle of how deep profiling of cancer resistance development can lead to an effective countermeasure, Melgar et al. studied adaptive resistance in FMS-like receptor tyrosine kinase (FLT3)–mutant acute myeloid leukemia (AML) that develops upon treatment with FLT3 inhibitors. Approximately 25% of patients with AML carry an internal tandem duplication of the FLT3 kinase gene associated with a very poor prognosis, and as such, providing a more effective therapy that is not associated with development of resistance aims to address an urgent medical need. To tackle the problem, here, the team examined the integrative in-cell kinase and gene regulatory network responses in tumors treated with FLT3 inhibitor. Innate immune pathway activation via the interleukin-1 receptor–associated kinase 1 and 4 (IRAK1/4) complex was found to contribute to adaptive resistance in FLT3-mutant AML cells (
first figure
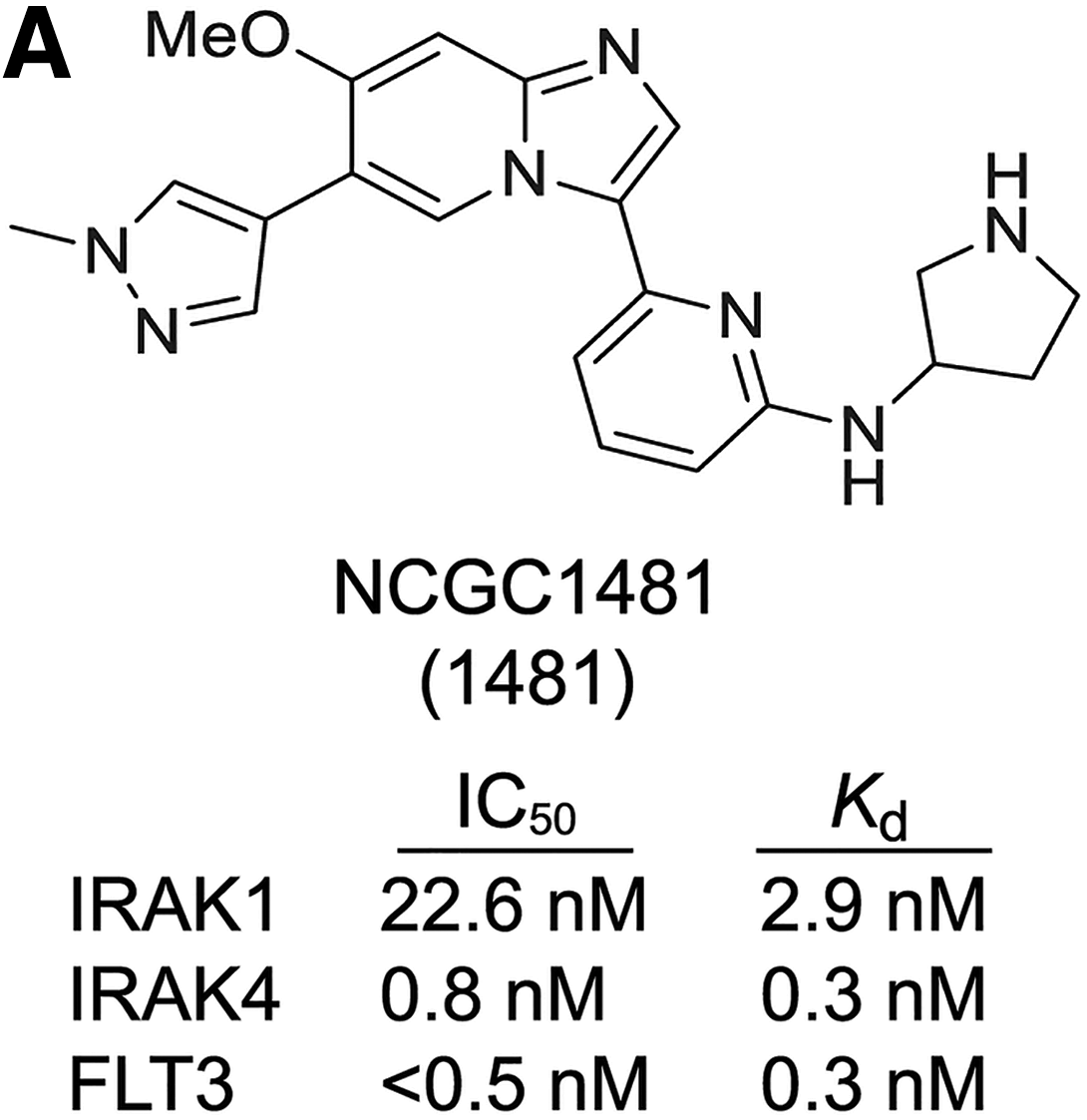
). Through extensive medicinal chemistry effort, the team developed a dual-acting drug candidate, NCGC1481, which simultaneously inhibits FLT3 and IRAK1/4 kinases (
second figure
) with low single-digit nanomolar potencies. The agent eliminated resistant AML cells in vitro and in vivo, effectively preventing the cancer cells from mobilizing that adaptive resistance pathway. The new compound also showed superiority relative to the established therapy in a comparison study of the antileukemic effects of NCGC1481 versus quizartinib using two different FLT3-mutant AML cells from patients with refractory leukemia (AML-174 and AML-019; figure 6 in the paper). Based on this study, it appears that a dual-acting frontline inhibitor of both the primary tumor target and the major resistance pathway presents a superior strategy to anticipate and tackle resistance upfront. Further work in this area will be needed to understand the durability of such treatment. Contributed by Anton Simeonov.
days and then replated in fresh medium, and cell viability was measured by AnnexinV staining. Values are expressed as means ± SEM from three biological replicates. (C) After 10 days in liquid culture [from (B)], the remaining viable cells were plated in methylcellulose, and colony formation was determined after 7 days. Values are expressed as means ± SD from two biological replicates. (D) Serine-threonine kinase (STK) PamChip analysis was performed on protein lysates isolated from MLL-AF9;FLT3-ITD and MV4;11 cells treated with quizartinib (0.3 nM) for 6 and 12 hours. Hierarchical clustering analysis was performed on differentially phosphorylated peptides in the indicated groups relative to DMSO (two biological replicates). (E) In-cell active kinases inferred from the phosphorylated peptides (STK PamChip) are shown for each of the indicated conditions. (F) Pathway enrichment of differential in-cell kinase activity in MLL-AF9;FLT3-ITD and MV4;11 cells treated with quizartinib for 6 and 12 hours was determined using Panther. (G) Pathway enrichment of differentially expressed genes (>2-fold, P < 0.05) in MLL-AF9;FLT3-ITD cells treated with quizartinib for 12 hours was determined using ToppGene (n = 3 per group).
NCGC1481 is a potent small-molecule inhibitor of FLT3 and IRAK1/4. (A) Chemical structure of NCGC1481 (1481). The IC50 and equilibrium dissociation constant (Kd) for NCGC1481 with IRAK1, IRAK4, and FLT3 is shown below the structure.