
Abstract
The CCR5Δ32 mutation does not account for HIV-1 resistance in the majority of persons who are repeatedly exposed to HIV-1 by high-risk activities but remain seronegative and uninfected. Therefore, we investigated the impact of CCR5 59029 A/G and CCL3L1 copy number polymorphism on HIV-1 disease susceptibility and progression among HIV-1-infected and HIV-1-exposed seronegative North Indians. HIV-1-seropositive (HSP, n = 196) patients, stratified on the basis of disease severity (Stages I, II, and III) and HIV-1-exposed seronegative (HES, n = 47) individuals were genotyped for CCR5-59029 A/G polymorphism by PCR-RFLP and CCL3L1 copy number by the real-time TaqMan PCR method. A group of ethnically matched HIV-1-seronegative (HSN, n = 315) healthy volunteers were also genotyped as controls. Statistical analysis was done by SPSS software. The CCR5-59029 AG genotype was significantly higher in the HES compared with the HSP group (57.44% vs. 37.24%, p = 0.014). The CCL3L1 mean copy number of HES was higher compared with the HSP groups (3.148 ± 0.291 vs. 2.795 ± 0.122, p = 0.212), but was not significant when compared with independent samples t test. Possession of CCL3L1 copies ≤2 or >2 was not associated with enhanced or reduced risk of HIV-1 acquisition. Gene–gene interaction studies showed enrichment of the CCR5-59029AG*CCL3L1>2 genotype in the HES group when compared with the HSP group (31.91% vs. 15.81%, p = 0.021, OR = 0.401, CI = 0.194–0.826). The increased frequency of the CCR5-59029AG*CCL3L1>2 genotype among HES individuals led us to conclude that the CCR5-59029 AG genotype and CCL3L1 gene dose appeared to have synergistic or interactive effects and are expected to be involved in the host innate resistance to HIV-1 infection.
Introduction
Genetic susceptibility to HIV infection and AIDS progression is variable among individuals and populations. There are individuals who have been repeatedly exposed to HIV-1 but have remained HIV-1 uninfected. A subset of such HIV-1-exposed seronegative (HES) individuals is HIV-1-seronegative (HSN) spouses of HIV-1-seropositive (HSP) individuals who are sexually exposed to HIV-1 repeatedly, yet remain negative for anti-HIV antibody. Immunologic and genetic studies of these individuals have helped to elucidate protective mechanisms for HIV-1 infection. 1 –4 The homozygosity for a mutation in the CCR5 gene (CCR5Δ32) is presently considered the most relevant against HIV-1 resistance, 5,6 which is rare among North Indians. 7 –9 We have recently reported that in the absence of individuals having the CCR5Δ32 mutation, the DC-SIGNR 5/5 homozygous genotype may a have partial role in conferring HIV-1 resistance among these HES individuals. 10
Among other host-dependent factors, attention has recently been paid to the variability in chemokine receptors and chemokines, which plays a central role in HIV-1 infection and disease pathogenesis. The principal chemokine receptor involved in HIV-1 transmission and progression is CCR5, and its most potent natural ligand is an MIP-1alpha (i.e., CCL3L1) chemokine. 11 An A/G polymorphism in the CCR5 promoter gene at nucleotide position 59029 was identified 12 and the CCR5-59029 GG homozygous genotype was reported to be associated with slower progression to AIDS. 12 –14 The CCR5-59029 G polymorphism has also been linked to decreased surface expression and reduced in vitro infectability. 15 CCR5Δ32/wt in combination with the CCR5-59029 A/G genotype has been reported to have an advantage in resisting sexual HIV-1 transmission in white men. 16
Independent studies have shown that an increased copy number of CCL3L1 results in the enhanced secretion of CCL3L1 by activated leukocytes. 17,18 CCL3L1 gene dosage has been reported to influence HIV-1 infection and pathogenesis. CCL3L1 copy numbers lower than the average for each ethnic group evaluated were associated with unfavorable outcomes, presumably due to lower expression and reduced capacity to block or interfere with HIV-1 binding to the CCR5 coreceptor. 17 Variations in the CCL3L1 copy number have been linked to HIV-1 susceptibility in asymptomatic Japanese patients with hemophilia, 19 while no such association was observed among HIV-1-infected North Americans. 20 The CCR5/CCL3L1 genotype combination also influences cell-mediated immunity (apart from viral burden to include viral entry-independent processes) in both healthy and HIV-1-infected individuals during the course of HIV infection. 21 Recent data also suggest that duplications of CCL3L1 are associated with the HIV-specific CD4 T cell responses that control HIV-1 infection. 22,23
The relationship between CCR5/CCL3L1 and HIV-1 susceptibility and disease progression is in dispute and remains to be confirmed in other populations and settings. Given such evidence, it was reasonable to hypothesize that in the absence of the CCR5Δ32 mutation, the CCR5-59029A/G/CCL3L1 copy number polymorphism may confer a protective effect in HES individuals. Thus, in the absence of any study examining the relationships of CCR5 and CCL3L1 to HIV-1-related outcomes in HIV-1-infected and HIV-1-exposed seronegative North Indian individuals, the present study was undertaken.
Materials and Methods
Patient selection
A total of 196 HSP were enrolled from the outpatients attending the clinics of Sanjay Gandhi PostGraduate Institute of Medical Sciences, Lucknow, India from January 2004 to December 2006. The HSP subjects were in different stages of disease progression and had no history or a less than 6 week prior history of antiretroviral therapy. Depending on their CD4+ T cell counts and based on their clinical symptoms, HIV-1 patients were divided into three subgroups, on the basis of disease severity, as per the Centers for Disease Control and Prevention (CDC) criteria. The three groups were as follows: Stage I (n = 86; asymptomatic HIV-1 patients in CDC category A1 to A3), Stage II (n = 61; symptomatic HIV-1 patients without AIDS in CDC category B1 to B3), and Stage III (n = 49; symptomatic HIV-1 patients with AIDS in CDC category C1 to C3). 24,25 Only individuals who had a minimum follow-up time of at least 6 months were included in the study. A total of 315 age-matched normal healthy controls of similar ethnicity with HSN status were recruited for the present study. Forty-seven HSN individuals with a history of repeated sexual intercourse (twice a week) without use of any protection (e.g., condoms) with HIV-1-infected partners for at least 1 year were recruited in the HES group. HIV-1-seronegative status of the HES subjects was confirmed by Western blot at the regular interval of 3 months to 1 year. Demographic profiles of the study groups are given in Table 1. After informed consent, a 5-ml blood sample was taken in EDTA for analysis of DNA.
Demographic and Clinical Characteristics of North Indian Individuals Classified on the Basis of HIV-1 Serological Status and Centers for Disease Control and Prevention Criteria a
HSN, HIV-1 seronegative; HES, HIV-1-exposed seronegative; HSP, HIV-1 seropositive; n, Number of individuals; M/F, male/female; ND, not detected.
These groups were subgroups of the HIV-1-seropositive patient and were classified according to CDC (Centers for Disease Control and Prevention). 23,24 Clinical symptoms in Category A: asymptomatic HIV infection, persistent generalized lymphadenopathy (PGL); Category B: hairy leukoplakia, oropharyngeal candidiasis, vulvovaginal candidiasis, constitutional symptoms such as fever or diarrhea lasting >1 month, herpes zoster; Category C; Pneumocystis carinii pneumonia, Mycobacterium tuberculosis (pulmonary and disseminated), candidiasis of bronchi, trachea, or lungs, extrapulmonary cryptococcosis, cytomegalovirus disease (other than liver, spleen, or nodes), isosporiasis, chronic intestinal (>1 months duration), HIV-related encephalopathy, Kaposi's sarcoma, wasting syndrome due to HIV.
Determination of HIV-1 status and CD4+ T cell counts
All individuals were screened for their HIV-1 status by primarily screening with ELISA (Vironostika, HIV Uni-FormII Ag/Ab, Biomerieux, The Netherlands) and subsequently confirmed with Western blot (LAV Blot I, Bio-Rad, France). CD4+ T cell counts were measured by flow cytometer FACScan (Becton Dickinson Immunocytometry Systems, San Jose, CA) using fresh EDTA-treated whole blood at the time of recruitment.
Genomic DNA isolation
The genomic DNA samples were obtained from 0.2–0.3 ml of peripheral whole blood using the QIAamp Blood kit (Qiagen, CA) according to the protocols supplied by the manufacturer. Usually ∼0.1 μg genomic DNA was used for the genotyping studies.
Genotyping of CCR5 59029-G/A promoter polymorphism
Genotyping was done by restriction fragment length polymorphism PCR (RFLP-PCR) analysis as previously described. 12 Amplicons including the polymorphic site of interest were amplified from extracted genomic DNA using the following primers: forward 5′-CCC GTG AGC CCA TAG TTA AAA CTC-3′ and reverse 5′-TCA CAG GGC TTT TCA ACA GTA AGG-3′. Of the PCR product 5 μl was digested with 4 units of Bsp1286I (MBI Fermentas) in a 20 (l volume at 37°C for 5 h using the buffer provided and bovine serum albumin. The digested product was loaded onto a 2% agarose gel, electrophoresed in a 1 × TBE buffer, and stained with ethidium bromide.
Genotyping of CCl3L1 copy number by real-time PCR (TaqMan) assays
The CCL3L1 gene copy number was estimated by TaqMan assays using Rotorgene 3000 (Corbet Research, Australia) and primers/probes (Tibmolbio, Berlin, Germany) according to previously described method. 17 The CCL3L1 gene was amplified by using the following primer sequences: forward, 5′-TCT CCA CAG CTT CCT AAC CAA GA3′; reverse, 5′-CTG GAC CCA CTC CTC ACT GG-3′; and probe, 5′-FAM (6-carboxyfluorescein, 6-FAM)–AGG CCG GCA GGT CTG TGC TGA–TAMRA (6-carboxytetramethylrhodamine, TAMRA). The housekeeping gene, β-globin (HBB), served as the internal control and was amplified using primer sequences as follows: forward, 5′-GGC AAC CCT AAG GTG AAG GC-3; reverse, 5′-GGT GAG CCA GGC CAT CAC TA-3′; and probe 5′-VIC-CAT GGC AAG AAA GTG CTC GGT GCCT–TAMRA. Seven serial 1:2 dilutions (50–0.78 ng) of genomic DNA from human A431 cells, known to contain two copies of CCL3L1 per diploid genome (pdg) by Southern blot densitometry, 18 were used on each plate to generate standard curves of C T (threshold cycle) value against the log [DNA] for HBB (also present at two copies) and the CCL3L1 gene. For each test sample, duplicate wells were set up for CCL3L1 and HBB. C T value was determined, and converted into template quantity using the standard curves. Copy number is the ratio of the template quantity for CCL3L1 to the template quantity for HBB, multiplied by two. The amount of test DNA sample added to each PCR reaction was between 2 and 10 ng.
Sequencing
PCR-amplified products were randomly selected to be sequenced according to the manufacturer's protocols (Applied Biosystems/Perkin Elmer, Foster City, CA) using the Dye Terminator cycle sequencing kit in ABI PRISM 310 genetic analyzer.
Statistical analysis
Statistical analysis was done by SPSS software version 11.5 (SPSS, Chicago, IL). The direct gene counting method was used to determine the frequency of genotypes and alleles. The Fisher exact or χ2 test was used to determine differences in allele/genotype frequencies of CCR5-59029 and copy number of CCL3L1 genetic variants. Association between the CCL3L1 mean copy number distributions among different groups was analyzed by independent samples t test. Odds ratios (OR) and its 95% confidence interval (CI) were obtained to describe the strength of association. A p value <0.05 was considered to be statistically significant. The sample size was calculated using the QUANTO ver. 1 program (
Results
CCR5-59029 A/G allele/genotype distribution analysis
Analysis of the genotype frequency data reveals significant deviation (p < 0.05) from HWE for the CCR5-59029 genotypes (Table 2). Deviation from expected values may be due to the excess of heterozygosity observed in CCR5-59029, which may be due to selection pressure. The CCR5-59029 AG genotype was significantly enhanced in the HES group compared to the HSP group (57.44% vs. 37.24%, p = 0.014) (Table 2). Further comparison of the CCR5-59029 A/G allele and genotype frequencies between Stage III and Stages I and II HIV-1-infected patients revealed no significant differences.
Genotype Frequencies of CCR5-59029 A/G in HIV-1-Seronegative, HIV-1-Exposed Seronegative, and HIV-1-Seropositive North Indians a
Data are no. (%) of subjects. AA, homozygous wild; AG, heterozygous; GG, homozygous mutant; CI, confidence interval; OR, odds ratio; HSN, HIV-1 seronegative; HES, HIV-1 exposed seronegative; HSP, HIV-1 seropositive. Stages I, II, and III are categorized on the basis of their CD4 counts and CDC classification. Statistical analysis: chi-square test by the SPSS statistical package.
HIV-1/AIDS susceptibility and CCL3L1 copy number variations
Estimation of CCL3L1 copy number was done by the quantitative ratio of CCL3L1 and HBB in the template genomic DNA evaluated by the quantitative TaqMan PCR method (Fig. 1). The number of CCL3L1 copies ranged from 1 to 9 per diploid genome in the control cohort with the most common being 2. Thirty-two percent of the North Indian HIV-1-seronegative healthy controls carried two copies of the CCL3L1 gene. No statistically significant differences in distribution of the CCL3L1 copy number between the study groups were found (data not shown), consistent with previous studies 17,19 that reported that the individual copy numbers of the CCL3L1 gene are not associated with HIV/AIDS clinical outcomes. The CCL3L1 mean copy number of HES was, though higher, compared with the HSP groups (3.148 ± 0.291 vs. 2.795 ±0.122, p = 0.212), but was not significant when compared with independent samples t test (Table 3). The median copy number in the HSN group was 2; therefore to test our hypothesis that a CCL3L1 copy number lower or higher than the median copy number influences the risk for HIV-1, subjects were divided into two groups: those with lower or equal than two (≤2) and higher than two (>2) copy numbers. Possession of CCL3L1 copies lower or equal to two (≤2) and higher than two (>2) was not associated with enhanced or reduced risk of HIV-1 acquisition (Table 4).
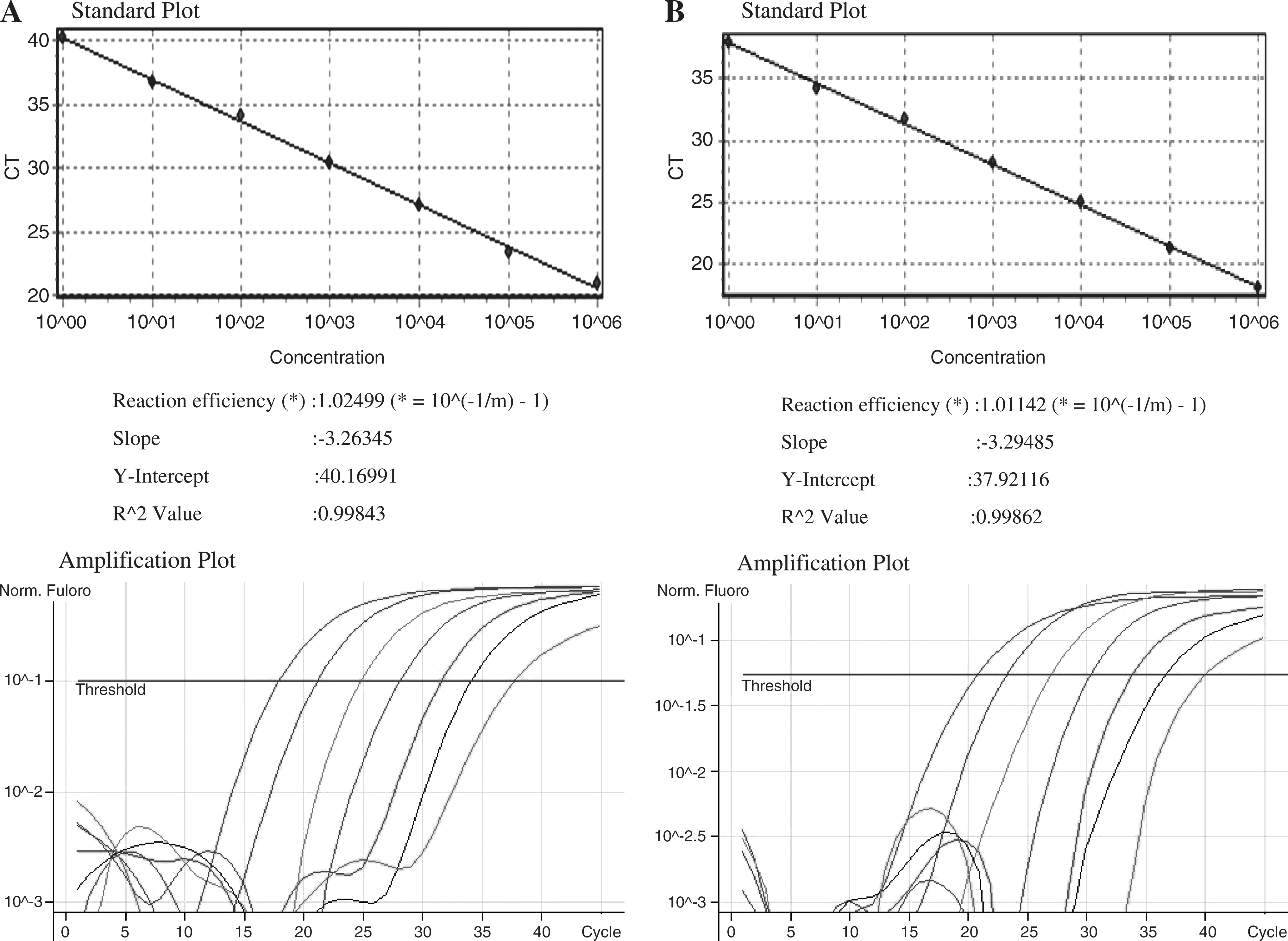
Standard curve and amplification plots obtained by Rotor-Gene 6.0.33 software for the human β-globin gene (
Mean Copy Number of CCL3L1 Genes among Different Study Groups a
HSN, HIV-1 seronegative; HES, HIV-1-exposed seronegative; HSP, HIV-1 seropositive; SD, standard deviation; SEM, standard error mean. Stages I, II, and III are categorized on the basis of their CD4 counts and CDC classification. Statistical analysis: independent-samples t test by the SPSS statistical package.
HSP vs. HSN.
HSP vs. HES.
Stage III vs. I.
Stage III vs. II.
Data are no. (%) of subjects. CI, confidence interval; OR, odds ratio; HSN, HIV-1 seronegative; HES, HIV-1-exposed seronegative; HSP, HIV-1 seropositive. Stages I, II, and III are categorized on the basis of their CD4 counts and CDC classification. Statistical analysis: chi-square test by the SPSS statistical package.
HIV-1/AIDS disease progression and CCL3L1 copy number variations
In the absence of number of years of known seroprevalence without antiretroviral therapy and viral load, the HSP group (n = 196) was categorized into Stages I, II, and III on the basis of their CD4+ T cell counts and clinical CDC classification. There was no significant difference in the distribution of the CCL3L1 copy number (data not shown) and its mean among categorized HIV-1-infected patients (Table 3). CCL3L1 copies lower or equal than two (≤2) and higher than two (>2) were also not associated with HIV-1/AIDS disease progression when compared with HIV-1 seropositive patients classified into different stages on the basis of disease severity (Table 4).
Combined effects of CCR5-59029 genotypes and CCL3L1 median copy number
On the basis of genetic association data obtained for CCR5-59029 genotypes and population-specific CCL3L1 median copy number, i.e., 2, the study subjects were divided into six genetic risk groups: (1) CCR5-59029AA*CCL3L1≤2, (2) CCR5-59029AA*CCL3L1>2 (3) CCR5-59029AG*CCL3L1≤2, (4) CCR5-59029AG*CCL3L1>2, (5) CCR5-59029GG*CCL3L1≤2, and (6) CCR5-59029GG*CCL3L1>2.
Thus, our working hypothesis was that a higher CCL3L1 copy number in association with a CCR5-59029 AG protective genotype might confer resistance in HES individuals against HIV-1 infection. Our data support the presence of an interaction between CCR5-59029 and CCL3L1 in the HES cohort only (Table 5). The frequency of CCR5-59029AG*CCL3L1>2 was significantly increased in the HES group when compared with the HSP group (31.91% vs. 15.81%, p = 0.021). Further comparison of CCR5-CCL3L1 genotypes between HIV-1-infected patients classified on the basis of severity of disease (Stages I, II, and III) did not show any significant association (Table 5). Further stratification of the groups by age and gender did not provide evidence of an association between CCR5 promoter alleles/genotypes and CCL3L1 copy number with HIV infection and progression (data not shown).
Comparison of CCR5-59029/CCL3L1 Combined Genotypes between HIV-1-Seropositive and HIV-1-Exposed Seronegative North Indians a
Data are no. (%) of subjects. AA, homozygous wild; AG, heterozygous; GG, homozygous mutant; CI, confidence interval; OR, odds ratio; HES, HIV-1-exposed seronegative; HSP, HIV-1 seropositive. Statistical analysis: chi-square test by the SPSS statistical package.
Discussion
Resistance to HIV-1 infection in some individuals has generated biological and genetic differences in explaining differential susceptibility to HIV-1 infection. Genetic variability in chemokine receptor genes and their ligands plays a major role in providing susceptibility to HIV-1 transmission and disease progression.
In the present study, we have provided evidence that the CCR5-59029 A/G polymorphism influences HIV-1 resistance among HIV-1-exposed seronegative North Indian individuals. The CCR5-59029 AG genotype was significantly enriched in HIV-1-exposed seronegative individuals, consistent with the previous studies in white men. 16 In contrast to previous studies, 12 –14,25 the inability of the present study to find any association between CCR5-59029 A/G polymorphism and delayed disease progression is most likely due to differences in ethnicity and environmental and social conditions of the populations. Our results were consistent with previous studies in a London cohort that failed to find any disease-retarding effect of the G nucleotide. 26 Our results were also consistent with the previous studies on North Indians 7 and white Europeans 27 that failed to show any significant association of the CCR5-59029 A/G polymorphism with disease progression among HIV-1-positive individuals. The present study further elucidated a possible genetic variant other than DC-SIGNR, previously reported in the same cohort, 10 that is operative in conferring resistance to HIV-1 infection among HIV-1-exposed seronegative North Indian individuals.
Results from our study demonstrated that 32% of the North Indian healthy controls have two copies of the CCL3L1 gene. It was lower than New Zealand controls (51%), 28 United Kingdom controls (53%), 28 European Americans (49%), 17 and white U.S. subjects (51%) 18 and higher than African Americans (16%) 20 and Japanese (10%). 19 Gonzalez et al. 17 reported that individuals with <2 copies of the CCL3L1 gene from the median for their population group presented a significantly higher risk for acquiring HIV-1 infection, whereas those with >2 copies above the mean had significantly less risk. Moreover, a gene dose lower than the cohort median was associated with increased risk of progressing more rapidly to AIDS and death. Similar findings by Nakajima et al. 19 also reported that the average copy number of CCL3L1 in hemophilic HIV-1-infected subjects was significantly lower than in controls and subjects possessing two or fewer copies of CCL3L1 had a significantly higher risk of acquiring HIV-1.
Our results, however, do not support this hypothesis, showing equal frequency of CCL3L1 copy number variants in control and HIV-1-infected groups. This is in agreement with other groups, which failed to find any significant association of CCL3L1 copy number with HIV-1 susceptibility/progression. 20 Moreover, Nakajima et al. also reported that CCL3L1 copy number variations had no significant effect on the disease progression among the long-term nonprogressor (LTNP) subjects, when compared with nonprogressors and patients under treatment. 19 Analysis of HIV-infected European individuals also failed to detect any statistically significant association between the distribution of CCL3L1 gene copy number and rate of progression to AIDS. 18 In the absence of finding any association between CCL3L1 copy number and HIV/AIDS susceptibility it can be speculated that expression of CCL3L1 mRNA may be independent of CCL3L1 gene copy number and may be regulated/influenced by other factors. 29 The discrepancy with respect to CCL3L1 gene dose and HIV-1 susceptibility between the present study and previous studies of Gonzalez et al. 17 and Nakajima et al., 19 though not clear, may be due to differences in racial or ethnic background, study design, and prevalence of virus subtype. The absence of number of years of known HIV-1 seropositivity could also be another factor that may influence disease progression in these patients. The integrity of the immune system, the antiretroviral schedules, the compliance with the therapeutic regimen, and the presence of the reactivation of chronic infections in HIV-1-infected patients classified as Stage III could also influence disease progression and could also explain the contradictory results obtained in different studies.
Investigating the association between possession of CCR5-59029/CCL3L1 combined genotypes and risk of acquiring HIV and/or disease progression, a higher frequency of CCR5-59029AG*CCL3L1>2 was observed in HIV-1-exposed seronegative compared to HIV-1-seropositive patients. Thus, the CCR5-59029 genotype and CCL3L1 gene dose appeared to have synergistic or interactive effects involved in the host innate resistance to HIV-1 infection. The CCL3L1 gene copy number is not the absolute parameter that confers HIV/AIDS susceptibility 29 ; rather it acts synergistically with other host factors. It is conceivable that the CCR5-59029 A/G and CCL3L1 copy number polymorphism evaluated in this study may potentially exert additive or synergistic influence with other factors, such as the antiviral immune response mediated by the CD8+ T cells and the levels of β-chemokines, the differential coreceptor expression on target cells, human leukocyte antigens (HLA), and cytokine polymorphisms. 30,31 Moreover, cellular factors such as cytosine deaminases APOBEC3G (apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G), protease inhibitors, α1-antitrypsin, and viral factors such as subtypes and the viral load presented either by the sexual partners of the exposed but uninfected individuals or by the infected individuals enrolled in this study could also exert some effects on the risk of acquisition and disease progression, respectively. 32 In addition, there are likely to be as yet unidentified host factors that are important in determining the outcome of HIV-1 infection. The protective effect of the CCR5-59029AG*CCL3L1>2 genotype against HIV-1 infection is further supported by evidence that reported an inverse association between CCL3L1 copy number and CCR5 expression on the T cell surface. 17 Recent studies have shown the presence of β-chemokine-mediated resistance against HIV-1 infection among HIV-1-exposed but -uninfected partners of HIV-1-infected individual. 30
Numerous studies in the past have provided important insight regarding the single nucleotide polymorphisms (SNPs) in host genetic factors influencing the susceptibility and resistance to HIV-1 infection. However, it is very difficult to attribute the effect of a particular gene, which could explain, by itself, the natural resistance to HIV-1 infection among sexually exposed but seronegative individuals and the disease progression in HIV-1-infected individuals. This is due to the fact that much of the genetic variation underlying infectious disease susceptibility is complex, involving the combination of many loci, as is further confounded by SNP variations between different populations and ethnic groups. However, further studies are required to confirm the present findings, correlating the MIP1-alpha concentration in serum samples with CCL3L1 gene copy number, and CCR5 expression on PBMCs with HIV-1 susceptibility in larger and well-established cohorts.
In conclusion, in the absence of an HIV-1 resistance conferring mutation, i.e., CCR5 delta 32, resistance to sexual transmission of HIV-1 infection among HIV-1-exposed seronegative individuals might be conferred by a synergistic effect of the CCR5-59029 AG genotype and CCL3L1 gene. The overall understanding of the molecular mechanisms underlying resistance to viral infection remains elusive and their elucidation may offer valuable clues for the development of HIV therapeutics and vaccines, providing a basis for the prevention of HIV/AIDS.
Footnotes
Acknowledgments
We gratefully thank Mr. S. Pandey and Mr. V.K. Mishra, Department of Microbiology, SGPGIMS, Lucknow for assistance in coordinating the clinical samples and all of the individuals who participated in the study. We also thank Mr. Nitin Semwal, Professional Biotech Pvt. Ltd., New Delhi for his excellent technical assistance. The Senior Research Fellowship provided by the Indian Council of Medical Research (ICMR, New Delhi) to Mr. Anurag Rathore (first author) is greatly acknowledged.
Disclosure Statement
No competing financial interests exist.