
Abstract
Strategies for purging persistent reservoirs in human immunodeficiency virus (HIV)-infected individuals may be enhanced by including agents that specifically kill virus-expressing cells. Anti-HIV envelope immunotoxins (ITs) represent one class of candidate molecules that could fulfill this function. We have previously utilized an anti-gp120 IT in conjunction with various stimulants to kill latently infected T cells ex vivo. Here we show that primary macrophages expressing HIV Env are relatively refractory to killing by IT when used alone. However, including stimulants such as prostratin or granulocyte–macrophage colony-stimulating factor to increase HIV gene expression in infected macrophages enhanced IT-mediated killing. Therefore, “activation–elimination” strategies similar to those proposed for purging the latent HIV reservoir may prove useful in clearing chronically infected macrophages in vivo.
W
A wide range of cytokines, antibodies, and other molecules is capable of stimulating HIV from latency, 11 –15 and it has been suggested that use of anti-HIV envelope (Env) immunotoxins (ITs) may accelerate the elimination of infected cells activated by such agents. 16 ITs are hybrid molecules consisting of a targeting domain derived from a monoclonal antibody linked to a toxic moiety. 17,18 Certain ITs and related targeted toxins have proven clinically useful as anticancer therapies, 19 –21 and in the context of HIV, various ITs have been developed that are effective against HIV-infected T cells and macrophages. 22 –24 We have previously shown that one such anti-gp120 IT can be used to kill latently infected T cells after stimulation ex vivo. 25 However, the effect that such “activation–elimination” strategies would have on other persistent viral reservoirs is uncertain.
In addition to latently infected CD4+ T cells, macrophages may also play a role in HIV persistence during therapy. Notably, in contrast to activated CD4+ T cells, 26 –28 macrophages are relatively resistant to the cytopathic effects of HIV, and can continue producing virus for weeks or months after infection in culture. 29 –31 While the frequency of persistently infected macrophages in patients undergoing HAART is not clear, this capacity for long-term chronic infection potentially allows infected macrophages in vivo to produce virus for months after initiation of HAART. Similarly, any temporal or spatial reductions in drug concentrations during HAART may lead to a “topping up” of the macrophage reservoir that could take months to reverse. Productively infected monocytes have been isolated from patients on HAART with sustained viral loads below 50 copies/ml, 32 –34 and although this observation requires further investigation, it does lend weight to the notion that monocyte/macrophage cells may represent an independent reservoir of replication-competent HIV in some patients. Macrophages are also the principal target cell type in potential anatomical reservoirs such as the central nervous system, 35 –37 where suboptimal drug concentrations may allow continued low-level virus replication with the potential to reseed the lymphoid system upon cessation of therapy.
The purpose of the current study was to determine what effect regimens suggested for use in purging the latent reservoir in CD4+ T cells would have upon chronically infected macrophages. Infected macrophages in HAART-treated patients are relatively rare in vivo, and it is difficult to obtain viable tissue samples from HIV-infected patients to directly test these cells for susceptibility to stimulants/immunotoxin. This sampling issue is particularly problematic in central nervous system tissues, where a large number of macrophages are present. Because of limited accessibility to these in vivo cells, we have used peripheral blood mononuclear cell (PBMC)-derived macrophages in this exploratory study. However, the majority of previously published macrophage studies suggest that infected macrophages act as chronic producers of virus, rather than truly latent reservoirs (where virus is not produced until the cell is stimulated). There are few data to suggest that these chronically infected macrophages differ in their response to cytokines or ITs from macrophages that have been infected with HIV for several days. For this reason we utilized peripheral cells infected for 2 days before exposure to stimulants and ITs as surrogates for assessing the effect of these factors on chronically infected macrophages
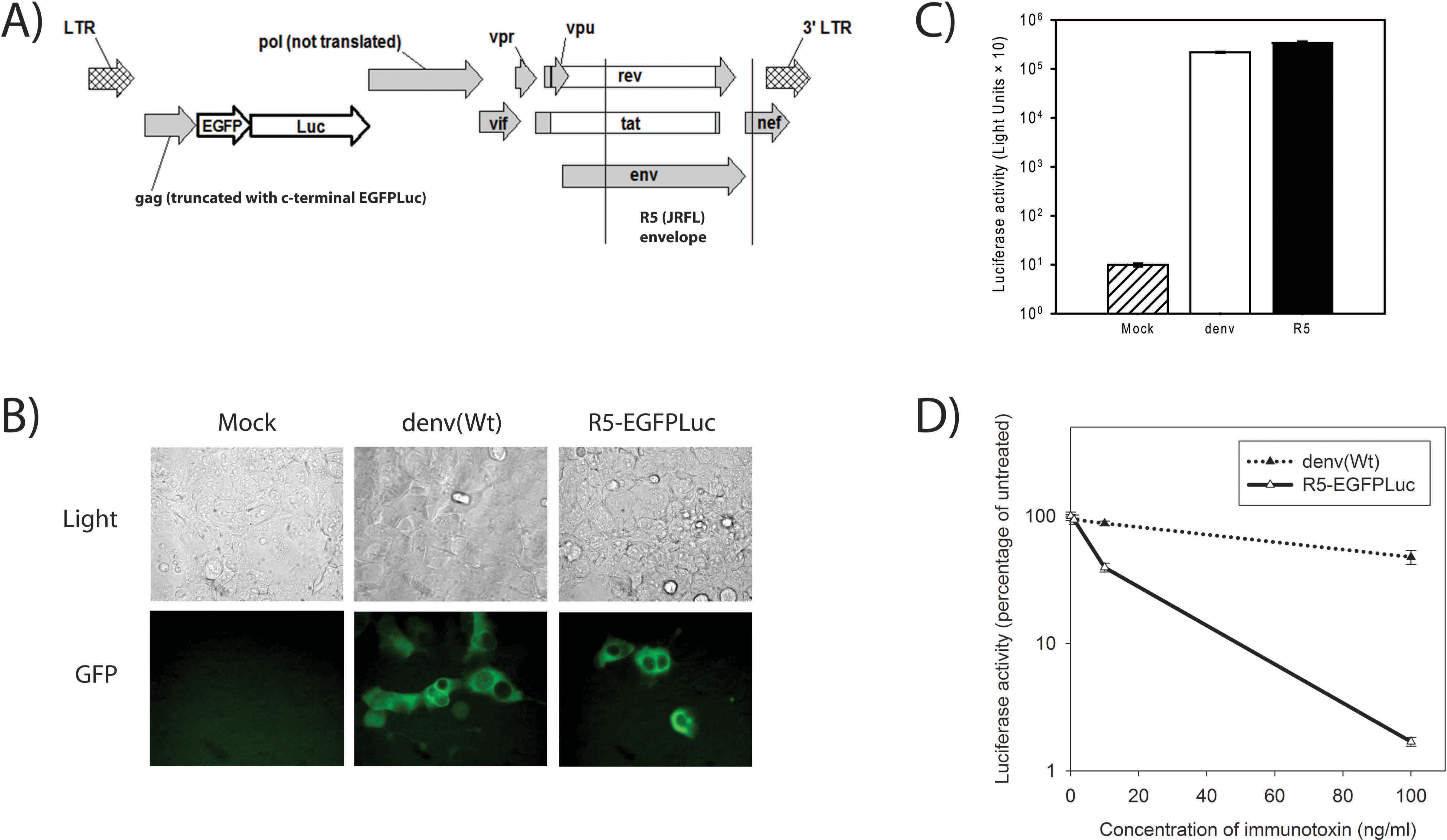
A new HIV-based reporter virus was constructed for this study (Fig. 1A). This virus was generated by first replacing the 1978-bp SpeI-AgeI region of NL4-3
38
with an EGFPLuc reporter gene, which encodes a fusion protein comprising an N-terminal enhanced green fluorescent protein (EGFP)
39,40
and C-terminal firefly luciferase (Luc).
41
The EGFPLuc coding sequence was amplified from plasmid pEGFPLuc (Clontech) by using a PCR with the sense primer 5′-ATTG
Testing of reporter viruses in 293FT cells. (
The new R5-EGFPLuc virus was initially tested in parallel with denv(Wt) by infecting 105 293FT cells at a multiplicity of infection (MOI) of approximately 0.1. At 3 days postinfection, GFP+ cells were visible in the infected cultures (Fig. 1B) and abundant Luc activity was present in infected cell lysates (Fig. 1C). To verify that an anti-HIV gp120 IT was active against cells infected with the R5-EGFPLuc but not against those challenged with the denv (Wt) virus, 293FT cells were infected as in Fig. 1C, then at 1 day post-infection, different concentrations of the IT 3B3:N31H/Q100eY[dsFv]-PE (HY-PE) 23,25,46,47 were added to each culture. At 3 days postinfection the cells were harvested and assayed for luciferase activity. As expected, cells infected with R5-EGFPLuc were susceptible to IT-mediated killing, while those infected with denv(Wt) were not (Fig. 1D).
To test infection of mature macrophages with R5-EGFPLuc, monocyte cells were isolated from PBMCs obtained from HIV-seronegative donors, essentially as previously described. 42 At 2 weeks postisolation, cells were scraped off the tissue culture flask and infected in suspension at an MOI of approximately 0.1. After infection, cells were washed and seeded at 12,500 cells per well in a flat-bottomed 96-well plate. Some wells were assessed for Luc activity and GFP expression at day 5 postinfection. Both Luc activity (Fig. 2A) and GFP expression (Fig. 2B) were present in all infected cultures, but absent from mock-infected cultures. Other wells of infected macrophages were maintained for up to 1 month in culture, and GFP+ cells were visible throughout this time (not shown), indicating that chronically infected macrophages can be created using this approach.

Testing of reporter viruses in primary macrophages. (
Cytokines and other potentially stimulatory molecules can have inconsistent or even opposing effects upon different stages of the HIV life cycle, and these effects can vary depending upon the specifics of the experiment and culture system. 48 Much of the prior work relating to the effect of stimulants upon HIV replication in primary macrophages has focused upon virus spread in culture, rather than the specific collection of events between integration and virus production. Since the latter situation is likely more important when developing strategies for purging chronically infected macrophages in vivo (which would be expected to already harbor an integrated provirus), we elected to focus exclusively on this phase of macrophage infection while developing our single-round infection assay. Integration takes approximately 2 days to complete in HIV-infected macrophages, 49 and GFP+ cells were present in the infected cultures at this time after infection with the R5-EGFPLuc virus (not shown). Hence, for testing the effect of stimulants and/or IT upon infected macrophages, these molecules were added at 2 days postinfection; cells were then lysed and assayed for Luc activity (HIV expression) at 5 days postinfection (Fig. 3A).

Effect of different molecules upon postintegration gene expression in HIV-infected macrophages. (
We tested different concentrations of several molecules in this manner, including interleukin (IL)-2, IL-7, and prostratin, each of which has been suggested for use as a component of purging strategies intended to eliminate the latent HIV reservoir within CD4+ T cells. 13,15,50 We also tested macrophage colony-stimulating factor (M-CSF), granulocyte-macrophage-stimulating factor (GM-CSF), and IL-4, which can significantly alter the efficiency of HIV spread in macrophages. 48 Using the current assay system, certain concentrations of both GM-CSF and prostratin resulted in up-regulation of HIV gene expression in the macrophage cultures (Fig. 3B), while the addition of IL-4, IL-7, or M-CSF did not significantly alter HIV expression levels. GM-CSF is FDA approved for treatment of certain neutropenias, 51 and the possibility of using prostratin as a therapeutic to aid in elimination of latent HIV is under evaluation in preclinical trials in nonhuman primates.
Notably, treatment with up to 1 μg/ml of HY-PE also failed to alter HIV gene expression (Fig. 3B), indicating that the IT alone has minimal effect on HIV-infected macrophages in this single-round infection system. One potential explanation for this result is that the majority of HIV budding in macrophages occur intracellularly, rather than directly at the cell surface plasma membrane. 29,30,52,53 This may impair the function of an anti-Env IT by reducing the concentration of Env at the cell surface that is available for recognition by the IT. It was thus possible that upregulating HIV expression could enhance IT-mediated killing by increasing Env expression at the cell surface. Consequently, assays were set up where stimulants and IT were added to infected macrophages, either alone or in combination. Combinatorial use of GM-CSF + IT (Fig. 4A) and prostratin + IT (Fig. 4B) each resulted in a significant reduction in luciferase activity (HIV expression) in cultures infected with R5-EGFPLuc, but not in those infected with denv(Wt). This reduction was less than that observed in either 293FT cells (Fig. 1D) or latently infected CD4+ T cells 25 treated with this same IT, which may reflect the different distribution of Env within these various cell types.
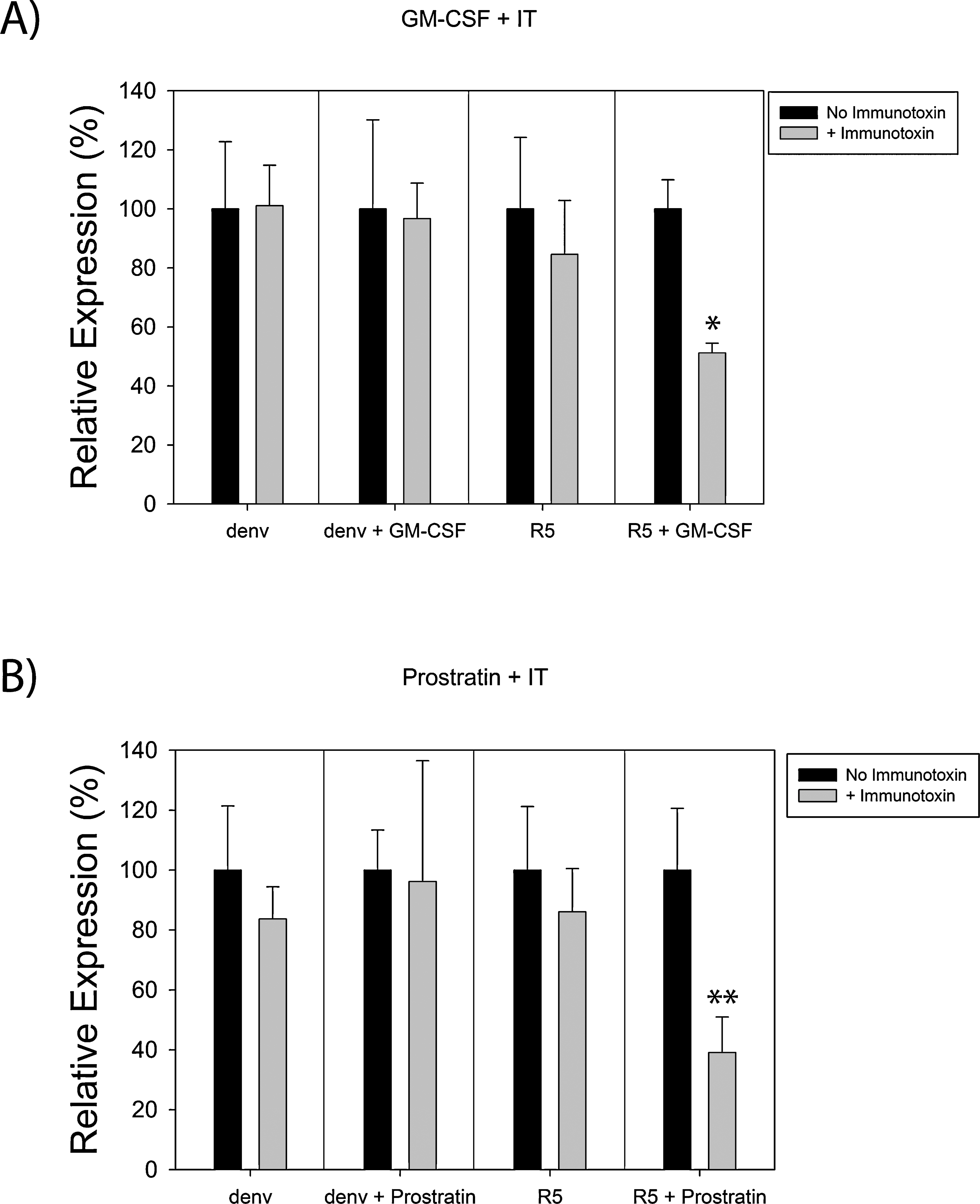
Treatment with a stimulant in conjunction with IT enhances killing of infected macrophages. (
Using stimulants such as prostratin in conjunction with anti-Env ITs represents one approach for purging the latent reservoir of HIV in patients undergoing treatment with HAART. It is likely that multiple ITs with different specificity would be needed to effectively target antigenically diverse clades of HIV. Yet the data described here indicate that suboptimal IT-mediated killing of HIV-infected macrophages can also be improved by stimulation of the host cell. In principle, this type of approach could be used in the future to help compensate for the low affinity that anti-HIV envelope agents, such as anti-gp120 ITs, may have for some of the diverse global isolates of HIV. In summary, activation–elimination strategies designed to eliminate latently infected CD4+ T cells may be adapted to simultaneously target chronically infected macrophages, potentially leading to a broader impact and improved efficacy against the various persistent reservoirs of HIV in HAART-treated patients.
Footnotes
Acknowledgments
This work was supported by NIH Grants AI70010, AI036059, and the UCLA CFAR.
Disclosure Statement
No competing financial interests exist.