
Abstract
Human plasmacytoid dendritic cells (PDC) are the major producers of type I interferons (IFN) after stimulation with CpG oligodeoxynucleotides (ODN). HIV-1-infected patients show a deficit in PDC numbers and function with progression of disease. CpG ODN appear to be attractive therapeutics to support the impaired innate immunity in HIV-1 infection. PDC counts, phenotype, and function were analyzed in 23 HIV-infected untreated individuals and 16 controls. Markers for migration (CCR7), activation (CD80), maturation (CD83), and endocytosis (BDCA2) were evaluated at baseline and 20 h after in vitro stimulation with class A, B, C, and P ODN. PDC counts and the expression of BDCA2 on these cells were significantly lower in HIV-1-infected subjects compared to controls (both p < 0.001). After stimulation with CpG ODN, CD80 and CD83 were upregulated to a similar extent in patients and controls, whereas CCR7 was upregulated more efficiently by CpG-P and CpG-C than CpG-A in HIV-1-infected individuals compared to controls. The IFN-α induction significantly differed for the CpG ODN classes (A > P > C > B) in patients and controls (p < 0.05). Functional PDC deficits in IFN-α and TNF-α induction were particularly evident in subjects with less than 500 CD4+ cells/μl. CpG-P ODNs not only induced remarkable IFN-α production in patient PBMCs, but also significantly upregulated the antibacterial and antiviral CXC chemokine IP-10. In conclusion, PDC counts, phenotype, and function are significantly impaired in HIV-1-infected subjects. Optimized P-class ODN may be effective in reversing this innate immune defect, which should be further evaluated in vivo.
Introduction
P
In HIV-1 infection, reduced PDC counts are associated with progressive disease. 10 –19 Similarly, functional PDC deficits have been observed, in particular reduced IFN-α production, 11,13,14,20 elevated expression of costimulatory molecules, 21 reduced allogeneic T cell proliferation, 22 and reduced surface expression of HIV-1 coreceptors CXCR4 and CCR5. 10 In addition, reduced upregulation of the migration marker CCR7 on stimulation with TLR7 agonists has been reported in HIV-1-infected children with immunologic or virologic failure, which may impair homing of PDC to lymphatic tissue and subsequent T cell activation. 23
In a search for molecules that boost innate immune responses, unmethylated CpG oligodeoxynucleotide (ODN) motifs were identified as the major immunostimulatory component of prokaryotic DNA. 24,25 Meanwhile, four classes of CpG ODN can be distinguished according to their immunostimulatory properties: class A activates PDC to high IFN-α production, whereas class B preferentially stimulates B cells and natural killer cells to elicit a Th1-based secretion of proinflammatory cytokines. 26 –29 The class C combines activities of the two former classes in inducing IFN-α production by PDC and stimulating NK and B cell activation. 30,31 Recently, a new class of CpG ODN was described, which contains two palindromic regions and forms higher ordered structures similar to class A but without the poorly defined G tetrad-driven aggregates. These class P ODN stimulate B cells and PDC, but lead to substantial higher IFN-α production compared to class C ODN. 31a
Class A and class B ODN were safely administered in healthy and SIV-infected macaques to boost the immunogenicity of a hepatitis B vaccine. 32 A similar effect was observed in adult HIV-1-infected subjects on HAART, who were vaccinated against hepatitis B in conjunction with a synthetic class B ODN. 33 A study in HIV-1-infected individuals showed reduced numbers of PDC, which, though chronically activated, could be further stimulated by ODN classes A, B, and C to produce IFN-α and IP-10. 34
We investigated the immunostimulatory effects of A, B, C, and P class ODN on the IFN-α production and surface marker expression by PDC of HIV-infected individuals in comparison to a matched control group. Our studies specifically focused on the question of whether any of these ODN may overcome the functional PDC deficit in HIV-1 infection.
Materials and Methods
Patient characteristics
All participants were monitored at an outpatient center specialized in the treatment and care of HIV-1-infected individuals, localized in Nürnberg, Germany (Table 1) None of these patients had received highly active antiretroviral therapy prior to study participation. A total of 11 individuals were asymptomatic, two patients were coinfected with hepatitis C, four subjects suffered from chronic bronchitis and allergic coryza, one subject from skin mycosis, one from diabetes mellitus, and another one from dizziness and fatigue with suspected HIV-encephalopathy. Three patients had evidence of sexually transmitted diseases. Two individuals were known to be infected for more than 10 years. Controls were age-matched healthy volunteers at the Institute of Clinical and Molecular Virology, University Hospital Erlangen, Germany (patients vs. controls, median 35 vs. 31 years, range 27–62 vs. 23–62 years, p = 0.53, not significant, n.s.). This study was approved by the ethical committee of the Medical Faculty, University of Erlangen-Nürnberg (Ref. no. 3375), and informed consent was obtained from all participants.
Isolation and stimulation of PBMCs
All blood samples obtained from patients or controls were processed within a few hours. Peripheral blood mononuclear cells (PBMCs) were isolated from EDTA-anticoagulated blood using standard Ficoll density centrifugation (Biochrom AG, Berlin), and plated at a density of 106 cells/500 μl in 24-well flat bottom plates (Greiner, Frickenhausen). Viability of isolated cells was checked by Trypan blue staining. PBMCs were cultivated in RPMI 1640 media (Gibco, Eggenstein) supplemented with 10% heat-inactivated (56°C, 60 min) fetal calf serum (FCS) (Cambrex, Verviers, Belgium), 50 mg/ml glutamine, 200 U/ml penicillin, and 90 U/ml streptomycin. The cells were stimulated in triplicate immediately after isolation using different concentrations and classes of CpG ODN (see reagents) for 20 h unless indicated otherwise. Supernatants were harvested and stored at −20°C for subsequent IFN-α determination.
Determination of cytokine production
The cell culture supernatants were analyzed for the production of IFN-α 2a/2b using an ELISA module set (Bender MedSystems, Vienna, Austria) according to the manufacturer's recommendations. Samples with values above the linear range were diluted as appropriate. The production of tumor necrosis factor (TNF)-α, IP-10, and interleukin (IL)-6 was assessed using the respective human FlowCytomix Simplex Kits with the Human Basic Kit (Bender MedSystems).
FACS analysis
Determination of cell counts was performed on EDTA-containing blood as described previously. 35 In brief, 2 ml of blood was washed once with phosphate-buffered saline (PBS) supplemented with 1% FCS and 0.5 mM EDTA (Sigma-Aldrich) and subsequently incubated with 100 μl of FcR-blocking reagent (Miltenyi Biotec, Bergisch-Gladbach) at 4°C for 10 min to reduce unspecific binding of staining antibodies. Cells were then incubated with a cocktail of FITC-conjugated lineage (lin) antibodies (CD3, CD14, CD16, and CD20; Immunotools, Friesoythe), PE-conjugated CD4 (Becton Dickinson, Heidelberg), and PE-Cy5-conjugated CD11c (Becton Dickinson); BDCA2-FITC and BDCA4-PE (Miltenyi-Biotec); CD3-FITC and CD4-PE/CD8-PE (all Becton Dickinson). Respective mouse IgG antibodies served as isotype controls. After staining at 4°C for 20 min, red blood cells were lysed in 155 mM NH4Cl, 10 mM KHCO3, and 0.1 mM EDTA. Cells were washed and fixed in 4% paraformaldehyde (Sigma-Aldrich, Taufkirchen) and 400,000 events were collected using a four-color FACSCalibur with CellQuest 3.3 software (BD Biosciences). Absolute differential white cell counts were provided by the Department of Transfusion Medicine and Hemostaseology (University Hospital, Erlangen). Cell counts were either expressed as percentage of PBMCs or absolute numbers (cells/μl) by multiplying percentages with absolute counts for mononuclear cells. Flow cytometry was also performed on PBMCs stimulated with CpG ODN to determine the expression of CCR7 (R & D Systems, Wiesbaden), CD80 (Immunotools), CD83 (Becton Dickinson), and BDCA2 on the surface of PDC, which were identified as positive for BDCA4 and negative for CD14 (Immunotools).
Reagents
Endotoxin-free CpG-A [ODN 6016, 5′-T*C-G-A-C-G-T-C-G-T-G-G*G*G*G-3′, where the asterisk (*) stands for phosphorothioate and a hyphen (-) stands for phosphodiester bonds], CpG-B (ODN 10103, T*C*G*T*C*G*T*T*T*T*T*C*G*G*T*C*G*T*T*T*T), CpG-C (ODN 2395, 5′-T*C*G*T*C*G*T*T*T*T*C*G*G*C*G*C*G*C*G*C*C*G-3′; ODN 10109, 5′-T*C*G*T*C-G*T*T*T*T*A*C-G*G*C*G*C*C-G*T*G*C*C*G-3′) and CpG-P (ODN 21798, 5′-T*C-G*T*C-G*A*C-G*A*T*C-G*G*C*G*C-G*C*G*C*C*G-3′) were kindly provided by Coley Pharmaceutical GmbH–A Pfizer Company, Düsseldorf, Germany. The non-CpG ODN 1982 (5′-T*C*C*A*G*G*A*C*T*T*C*T*C*T*C*A*G*G*T*T-3′) served as control. ODN concentrations used for stimulation were 0.75 μM (ODN 6016, ODN 2395, ODN 10109) or 0.25 μM (ODN 10103, ODN 21798), unless otherwise indicated.
Statistics
The Wilcoxon signed-ranks test and the Mann–Whitney U-test were used for comparisons between paired and unpaired data, respectively. Two-sided p values ≤ 0.05 were considered significant.
Results
Altered cell counts in HIV-infected subjects (n = 23) vs. controls (n = 16)
The group of HIV-infected individuals recruited for this study included newly diagnosed (n = 3) and long-term infected subjects (n = 2) (Table 1). One individual had undetectable viral load in the plasma, whereas the median viral load in the other 22 patients was 20,500 copies/ml (range 660–200,000 copies/ml). A total of 2, 9, and 12 HIV-infected subjects had less than 250, 250–500, and more than 500 CD4+ cells/μl, respectively. CD4+ cell counts were significantly lower and CD8+ cell counts significantly higher in the HIV-1-infected subjects compared to the age-matched controls (Table 2), resulting in a significantly altered CD4/CD8 cell ratio (p < 0.001, data not shown). As has been described by other groups in similar studies, percentages (Fig. 1a) and absolute counts of CD4+ lin− CD11c− PDC were significantly lower in patients compared to controls (both p < 0.001) (Table 2). In HIV-infected subjects with 250–500 CD4+ cells/μl, the percentage of CD4+ lin− CD11c− PDC was significantly lower compared to subjects with more than 500 CD4+ cells/μl (median, 0.13 vs. 0.32, IQR 0.11–0.17 vs. 0.20–0.39, respectively; p = 0.01).

Expression of plasmacytoid dendritic cell (PDC) surface markers at baseline in HIV-1-infected subjects (pat.) (n = 23) and control donors (ctrl.) (n = 16). Quantification of the percentages (%) of peripheral PDC as (
Data are presented as median and interquartile ranges. Lineage, lin; not significant, n.s.
Reduced expression of BDCA2 on the PDC of HIV-infected subjects
The difference in PDC counts between patients and controls was also observed when PDC were characterized as BDCA2+ BDCA4+ cells (Fig. 1b and Table 2) (p < 0.001). No differences were detected between HIV-infected subjects with 250–500 CD4+ cells/μl and subjects with more than 500 CD4+ cells/μl (median, 0.13 vs. 0.18, IQR 0.10–0.16 vs. 0.12–0.32, respectively; p = 0.24, n.s.). This effect was due to downregulation of BDCA2 on the PDC of HIV-1-infected subjects (Fig. 1c), which was significant in comparison to the control donors (p < 0.001) (Fig. 1d). These data were corroborated by the mean fluorescence intensities (MFI) of BDCA2 on the PDC of HIV-infected (median, 24.45; IQR, 14.91–35.80) and control donors (median, 48.83; IQR 35.79–76.56) (p = 0.002) (Fig. 1e), whereas BDCA4 was comparably expressed on the PDC of both groups (median, 55.41 vs. 54.82; IQR, 42.23–76.77 vs. 35.23–89.93) (p = 0.74, n.s.). Thus, HIV-infected patients showed reduced counts of BDCA2+ BDCA4+ PDC and an altered phenotype with downregulation of BDCA2 at the cell surface.
In parallel, alterations of the PDC phenotype in HIV-1 infection were systematically investigated, focusing on surface markers for migration (CCR7), activation (CD80), and maturation (CD83). CCR7 was detected at lower levels in HIV-infected individuals; however, this difference was not significant (p = 0.16, Fig. 1d). CD80 and CD83 were not differently expressed on patient and control PDC (p = 0.66 and p = 0.64, respectively) (Fig. 1d).
Similar IFN-α induction profiles in HIV-infected subjects and controls
The CpG ODN were used at increasing concentrations to stimulate control PBMCs for 20 h. The optimal concentration to induce IFN-α production was 0.75 μM for CpG-C (2395) and CpG-C (10109) and 0.25 μM for CpG-B (10103) and CpG-P (21798). The optimal concentration for CpG-A (6016) was determined to be 0.75 μM, because the amount of IFN-α increased only marginally at a 3-fold higher ODN concentration (Fig. 2a). The IFN-α production significantly differed between the four ODN classes (A > P > C > B). P-class ODN 21798 induced significantly higher amounts of IFN-α compared to the C-class ODN 2395 and 10109 (both p = 0.005), which induced comparable amounts of IFN-α (Fig. 2a). When PBMCs of HIV-1-infected individuals were evaluated in parallel, similar data were obtained for the optimal ODN concentrations used for stimulation (Fig. 2b). Also in these subjects, the IFN-α production was significantly higher after stimulation with the new class P ODN 21798 compared to the CpG-C ODN (both p = 0.05) (Fig. 2b). These data indicate that PDC of HIV-1-infected individuals show similar IFN-α induction profiles compared to uninfected controls.

Induction of interferon (IFN)-α production by class A, B, C, and P CpG oligodeoxynucleotide (ODN) stimulation for 20 h in PBMCs obtained from control donors (n = 10) or HIV-1-infected subjects (n = 15). Determination of the optimal ODN concentration in (
Functional PDC deficits in HIV-infected individuals in response to CpG-A and CpG-P stimulation
Next, the extent of IFN-α induction was compared in patient (n = 23) vs. control (n = 16) PBMCs. Significant differences were observed for the stimulation with CpG-A (6016) (p = 0.01) and CpG-P (21798) (p = 0.02), but not with the other ODN (data not shown). When the HIV-infected subjects were grouped according to their CD4+ cell count, the IFN-α induction by all ODN classes was significantly lower in patients with 250–500 CD4+ cells/μl compared to those with more than 500 CD4+ cells/μl (p < 0.05) (Fig. 3a). In contrast, no significant differences in IFN-α levels were observed between patients with more than 500 CD4+ cells/μl and controls. To separate numerical from functional PDC defects, the IFN-α induction was normalized for percentages of PDC within the PBMCs for each individual donor. After this adjustment, the differences in IFN-α induction remained significant for all classes (p < 0.05), when patients with more or less than 500 CD4+ cells were compared (data not shown). Similar data for class B, class A, and class C ODN were recently reported for HCV-infected patients. 36 Altogether, our data indicate that PDC of HIV-infected subjects with less than 500 CD4+ cells/μl are functionally impaired in responding to CpG ODN stimulation.
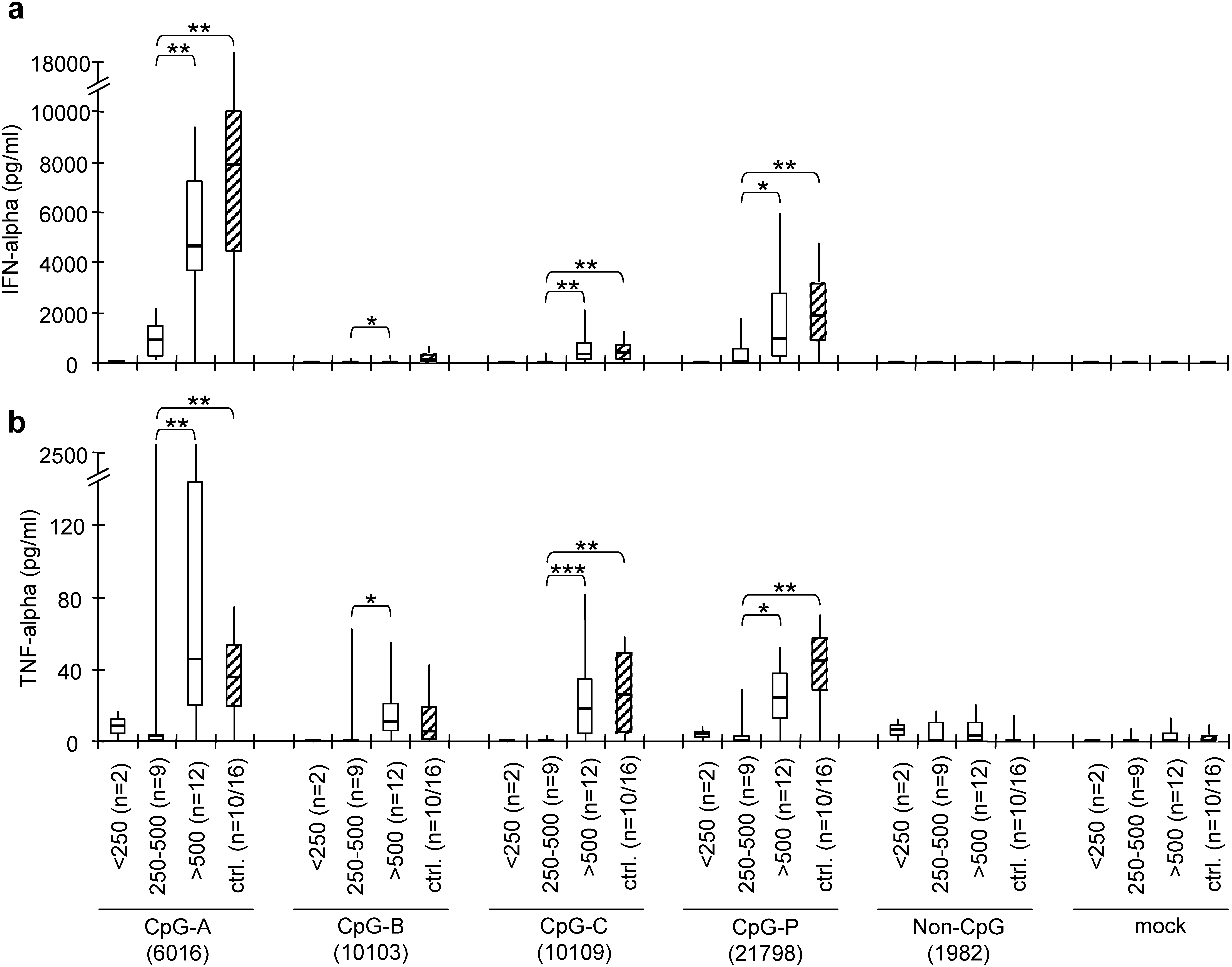
Induction of (
Induction of other cytokines by CpG ODN stimulation
TNF-α is produced by monocytes, macrophages, and to a minor extent by PDC. In control donors, TNF-α induction significantly differed between the ODN classes (A = P > C > B; p < 0.05). The CpG ODN induction profile was similar to IFN-α with significantly reduced TNF-α induction in patients below 500 CD4+ cells/μl (Fig. 3b). The CXC chemokine IP-10 is produced by various cell types and exhibits antibacterial and antiviral qualities. 37,38 It is reported to be indirectly induced in monocytes by CpG ODN-stimulated PDC. 39 In control donors, CpG-P induced higher levels of IP-10 compared to the other ODN classes (P > A > C > B; all p < 0.05). Notably, the PBMCs of HIV-infected individuals with more than 500 CD4+ cells/μl secreted significantly more IP-10 on CpG-A and CpG-P stimulation compared to controls (p < 0.05) (Fig. 4a). The IP-10 induction in HIV-infected subjects with 250–500 CD4+ cells/μl was comparable to control donors. IL-6 has been shown to be secreted by CpG-stimulated B cells. 40 In control donors and HIV-infected subjects, the IL-6 induction was not significantly different between the ODN classes. As with IP-10, the highest IL-6 values were observed, when PBMCs of HIV-infected individuals with more than 500 CD4+ cells/μl were stimulated using CpG-A and CpG-P (Fig. 4b).
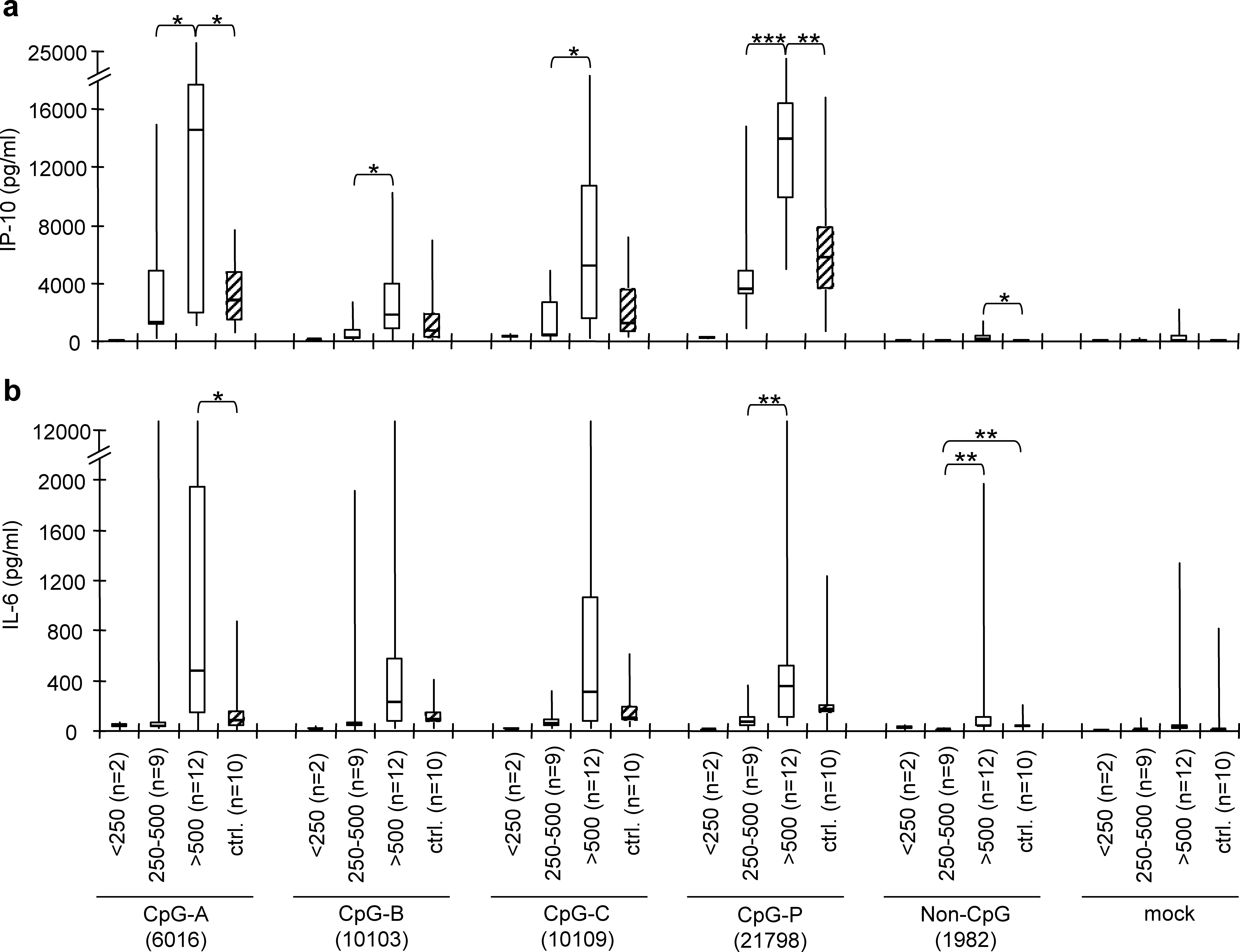
Induction of (
Comparable inducibility of PDC surface markers in HIV-infected subjects and controls by CpG ODN stimulation
The regulation of PDC surface markers was evaluated after exposure of PBMCs to different classes of CpG ODN for 20 h. In patients and controls, CCR7 (Fig. 5a), CD80 (Fig. 5b), and CD83 (Fig. 5c) were significantly upregulated on ODN exposure in comparison to mock-stimulated cells (all p < 0.05). These surface markers were also significantly upregulated by the control ODN 1982. This may be due to its phosphorothioate backbone, which is known to interact with TLR9. 41 The endocytosis marker BDCA2 was significantly downregulated on the PDC of control donors after exposure to B-, C-, and P-class ODN (all p < 0.01) (Fig. 5d). This effect was not observed in the PDC of HIV-1-infected subjects, because BDCA2 was already downregulated in mock-stimulated cells (Fig. 5d).

Regulation of plasmacytoid dendritic cell (PDC) surface markers after stimulation with different classes of CpG oligodeoxynucleotides (ODN). Markers for (
Discrepant effects of class A, C, and P ODN on the CCR7 phenotype in HIV-infected individuals
Because the PDC of HIV-1-infected individuals expressed lower levels of CCR7 at baseline, we specifically investigated the effects of CpG ODN stimulation on this migration marker. In the PDC of control donors, CCR7 was upregulated by all classes of CpG ODN (all p < 0.01) (Fig. 5a). In contrast, CpG-A ODN 6016 was significantly less active in upregulating CCR7 on the PDC of HIV-1-infected compared to control donors (p < 0.05). These data suggest that PDC are particularly impaired in responding to CpG-A stimulation in HIV-1 infection.
Dynamics of PDC surface receptor regulation after CpG ODN stimulation
To obtain more detailed information about the time frame in which PDC surface receptors are regulated, PBMCs of three uninfected control donors were exposed to the P-class ODN 21798 and analyzed 3, 6, 9, 12, 24, and 48 h poststimulation (Fig. 6a). CCR7 and CD83 were upregulated within the first 3–6 h of exposure, whereas BDCA2 was downregulated within the first 24 h. Activation of PDC as evident from CD80 upregulation was not observed before 48 h of stimulation. These data indicate that CpG ODN stimulation results in a timely coordinated upregulation of CCR7, CD80, and CD83 together with a downregulation of BDCA2. A parallel experiment using PBMCs of four HIV-1-infected donors revealed a delayed dynamics of surface marker regulation (Fig. 6b).
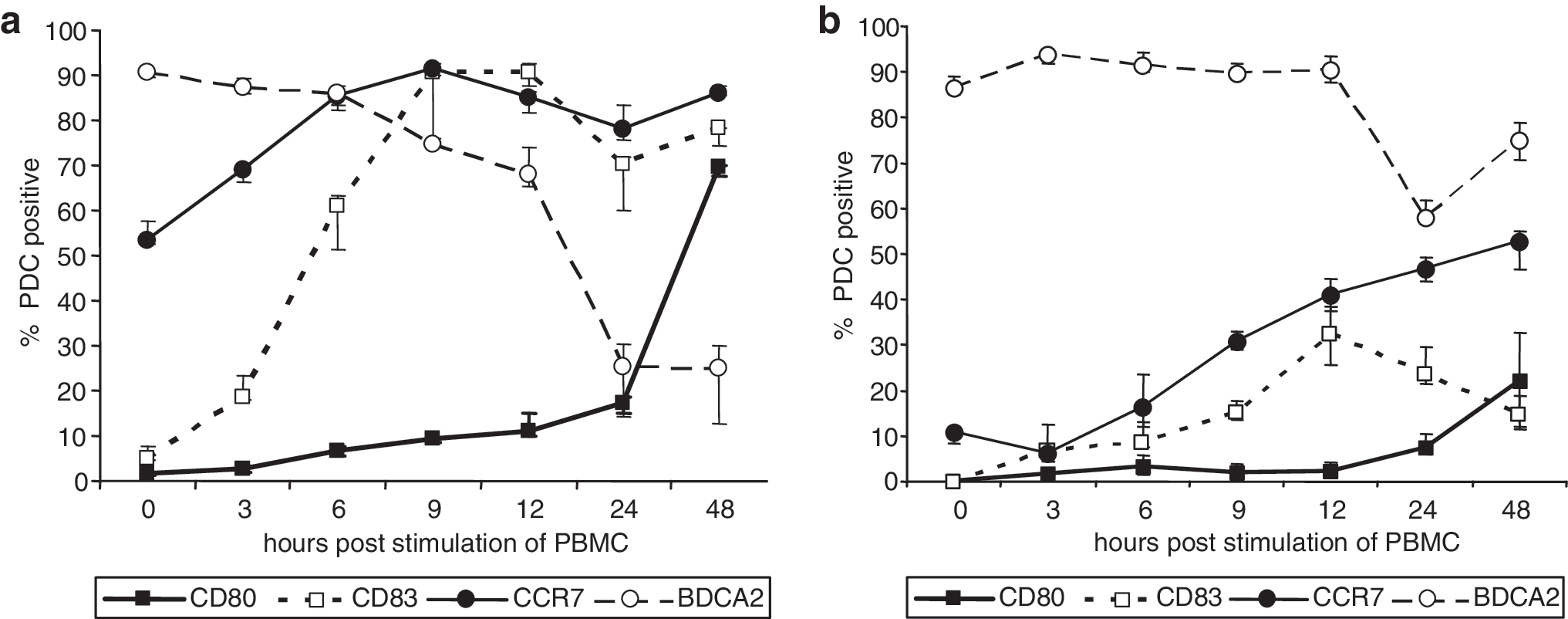
Dynamics of plasmacytoid dendritic cell (PDC) surface receptor regulation after CpG-P stimulation (ODN 21798). Surface markers for migration (CCR7), activation (CD80), maturation (CD83), and endocytosis (BDCA2) were evaluated on PDC at different time periods after stimulation of PBMCs. Data are presented as median and interquartile ranges and are representative of (
Discussion
Our study analyzed the phenotype and function of PDC in HIV-1-infected individuals and controls after stimulation of PBMCs with different CpG ODN classes. Our study was limited in the number of study participants, in particular subjects with less than 250 CD4+ cells/μl. The latter reflects the current guidelines for antiretroviral therapy. Still, we could conclusively show that PDC counts, phenotype, and function were severely impaired in HIV-1 infection. In this respect, we observed a significantly lower expression of CCR7 after CpG-A stimulation (Fig. a) on the PDC of HIV-1-infected individuals. Similarly, it has been reported that CCR7 was not adequately upregulated on TLR7 stimulation in antiretrovirally treated HIV-infected children with immunologic or virologic failure. 23 These data may indicate that the migratory capacity of PDC to secondary lymphatic tissue is reduced, subsequently affecting T cell activation. In addition, the calcium-dependent lectin BDCA2 was significantly downregulated on the PDC of HIV-1-infected individuals at baseline (Fig. 1d) and after cultivation for 20 h without stimulation (Fig. 5d).
The surface molecule BDCA2 was described as being endocytosed and recycled to the cell surface when PDC were incubated with BDCA2 antibodies; this ligation suppressed the induction of type I IFN production by different stimuli. 42 Reduced BDCA2 expression may indicate a reduced endocytic capacity to respond to pathogen-associated molecular patterns, further aggravating impaired PDC innate immune responses in HIV-1 infection. Recently, the reduced responsiveness of PDC to de novo stimulation has been associated with upregulated expression of IFN-stimulated genes, arguing that the diminished function may be a consequence of prior activation of PDC via type I IFNs or HIV particles in vivo. 43 In our study, however, plasma IFN-α levels were not significantly different between HIV-1-infected and control subjects (data not shown). In addition, the pattern of reduced expression of BDCA2 on the PDC of HIV-1-infected subjects at baseline and the delayed dynamics of surface marker regulation (Fig. 6b) rather argue for silencing of PDC than in vivo activation.
In addition to phenotypic alterations in PDC of HIV-1-infected subjects, we observed a reduced IFN-α production (Fig. 3a), as described by others. 11,13,14,20,34 Notably, the sensitivity of responses to different CpG classes was comparable between patients and controls, i.e., the peak of IFN-α production was not shifted to higher ODN concentrations in HIV-infected subjects, but the amounts of IFN-α produced were substantially lower (Fig. 2). Furthermore, the rank order between different CpG ODN classes (A > P > C > B) in IFN-α induction was comparable between patients and control, suggesting that the PDC of HIV-1-infected subjects responded in a similar manner, but to a more limited degree. 44 When the low concentration of 0.25 μM was used for stimulation, the class P ODN 21798 and the class A ODN 6016 induced comparable amounts of IFN-α in PBMCs of control donors. At this concentration, ODN 21798 was even more active compared to ODN 6016 in the PBMCs of HIV-1-infected individuals.
The differences between the IFN-α induction in PBMC of HIV-1-infected and control donors were explained to some extent by the lower PDC counts of HIV-1-infected subjects. However, additional functional deficits were particularly evident in patients with less than 500 CD4+ cells/μl after stimulation with class A, class P, and class C ODN (Fig. 3a). Similar deficits were also observed for the induction of TNF-α production (Fig. 3b), indicating that PDC functions are already impaired when HIV-infected subjects present with CD4+ cell counts above the threshold for antiretroviral therapy. The picture was different for the induction of IP-10 and IL-6 production. IP-10 is mainly produced by monocytes, which are stimulated by IFN-α produced by CpG-stimulated PDC, 39 and IL-6 by CpG-stimulated B cells. 40 CpG-P ODNs proved to be particularly effective in inducing IP-10 (Fig. 4a). In HIV-infected patients with 250–500 CD4+ cells/μl, the IP-10 induction was comparable to controls. Notably, HIV-infected subjects with more than 500 CD4+ cells/μl responded to CpG-P and CpG-A stimulation with significantly elevated IP-10 and IL-6 production compared to controls (Fig. 4). One explanation may be that HIV-infected subjects had higher monocyte counts (Table 2), which respond to CpG stimulation (i.e., bacterial infection) as long as PDC counts and function are not significantly impaired. The excessive IP-10 and IL-6 production may also reflect the chronic immune stimulation of monocytes and B cells in HIV-1 infection.
The upregulation of CD80 (Fig. 5b) and CD83 (Fig. 5c) after CpG ODN stimulation shows that PDC of HIV-1-infected subjects can respond to TLR9 ligation, as similarly shown by others. 34 When the upregulation of CCR7 was investigated on the PDC of HIV-1-infected individuals, different stimulation profiles were observed for class A, class C, and class P ODN: CCR7 remained significantly downregulated with CpG-A, but was upregulated with CpG-C and CpG-P ODN (Fig. 5a). Thus, class P and class C ODN may be beneficial in enhancing the migratory capacity of PDC in HIV-1 infection. It has to be considered, however, that our study was performed in a group of HIV-infected individuals with heterogeneous disease progression. Therefore, effects of ODN stimulation should be separately analyzed in larger cohorts with either long-term nonprogressive infection, progressive infection, or primary infection. Since our study investigated the effects of CpG ODN only in cell culture, it remains to be determined whether similar effects can be observed when ODNs are administered to patients.
Class A and class B ODN have successfully been applied as adjuvants in SIV-infected macaques and HIV-infected patients in boosting HBV humoral immune responses. 32,33 It has to be noted, though, that anti-HBs levels were still significantly lower in infected animals and humans than in uninfected controls. A and B class ODN were safely administered in SIV-infected monkeys. 32 In HIV-1-infected patients on antiretroviral therapy, however, a transient decline of CD4+ cells was observed for 1–2 weeks after administration of a B-class ODN. 33 Recently, a phase III trial in HIV-uninfected adults using a hepatitis B vaccine plus immunostimulatory sequences was put on hold by the FDA, because one serious adverse event, namely systemic vascular inflammation, occurred. Induction of IFN-α production (and potentially other cytokines as well) may in fact be harmful in the context of HIV-1 infection. 45 It also needs to be evaluated whether the upregulation of CCR7 may result in a decline of peripheral PDC counts, which might impair their sentinel function in detecting opportunistic pathogens. This effect may be particularly relevant if a chronic administration of CpG ODN is envisaged as immunostimulatory and antiviral drug, as has been investigated in patients with chronic hepatitis C infection. 36,46 Altogether, the new P class ODN appear to have a unique profile in inducing proinflammatory cytokines and enhancing CCR7 expression, which may be beneficial in boosting innate and adaptive immune responses in HIV-1-infected patients.
Footnotes
Acknowledgments
The data were presented in part at the 3rd European Congress of Virology, September 1–5, 2007, Nürnberg, Germany. We thank Bernhard Fleckenstein for continuous support. We want to acknowledge funding by the German Competence Network HIV/AIDS, the German Research Foundation (SCHM 1702/1-1; SFB466, Project A12; SCHM 1702/2-1; SCHM 1702/2-2), the ELAN-Fonds (VI-08-06.18.1), the “Akademie der Wissenschaften und Literatur zu Mainz,” and Coley Pharmaceutical GmbH. The latter provided the CpG ODN for testing their immunostimulatory capacity in cells of HIV-1-infected and -uninfected subjects. Philipp Schuster is supported by the graduate college 1071 (“Viruses of the immune system”). We thank Arthur M. Krieg for helpful comments on the manuscript.
Author Disclosure Statement
Jörg Vollmer has a potential conflict of interest as he is an employee of Pfizer and provided the CpG ODN used in the study. The company had no influence on the study design; collection, analysis, and interpretation of data; writing of the report; and the submission of the manuscript.