
Abstract
Axin1, a regulator of Wnt signaling, was previously identified as playing a negative role in the late phase of human immunodeficiency virus type 1 (HIV-1) replication in HeLa-derived J111 cells. In this report, we studied the molecular mechanism of how Axin1 regulates HIV-1 replication. HIV-1 transactivator, Tat-dependent viral reporter gene expression was enhanced in J111 cells transfected with small interfering RNA (siRNA) against Axin1. In addition, viral transcription was upregulated in J111 cells transfected with siRNA against Axin1. In contrast, HIV-1 gene expression was not enhanced by transfecting HeLa cells with siRNA against Axin1. The expression levels of T cell factor-4 (TCF4) and β-catenin were higher in J111 than HeLa cells. In addition, siRNAs against TCF4 and β-catenin inhibited the Axin1 siRNA-dependent enhancement of HIV-1 gene expression in J111 cells. These results suggest that Axin1 plays a negative role in HIV-1 transcription through the Wnt signaling pathway in J111 cells under normal cell culture conditions.
RNA
Axin1 is a negative regulator of the Wnt signaling pathway that is involved in the regulation of many biological events, including cell growth and differentiation. 5 A hallmark of canonical Wnt signaling is the stabilization of cytoplasmic β-catenin followed by its nuclear translocation, which leads to the transcriptional activation of Wnt target genes. 5 Under unstimulated conditions, β-catenin as well as Axin1, adenomatous polyposis (APC), and glycogen synthase kinase 3β (GSK3β) form a β-catenin destruction complex, and β-catenin is constitutively phosphorylated by GSK3β. Phosphorylated β-catenin becomes ubiquitylated and is targeted for degradation by the proteasome. Following activation of the Wnt signaling pathway, β-catenin is released from the Axin1–APC–GSK3β complex, which leads to the cytoplasmic accumulation of hypophosphorylated β-catenin. Accumulated β-catenin translocates to nuclei, and regulates the expression of Wnt target genes through cooperation with the T cell-specific transcription factor/lymphoid enhancer-binding factor 1 (TCF/LEF) family of transcription factors. 5
Our previous observation indicated that Axin1 is involved in the regulation of the late phase of HIV-1 replication 2 ; however, it was still unclear whether the Wnt signaling pathway and/or other Wnt signaling regulators were involved in the regulation of HIV-1 replication. In this report, we performed detailed analysis to elucidate the molecular mechanism of how viral replication is regulated by Axin1.
HeLa and J111 cells were maintained in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA) supplemented with 10% heat-inactivated fetal calf serum. J111 cells (JCRB0073, Lot #030395; Japan Health Sciences Foundation, Tokyo, Japan) were originally described as human acute monocytic leukemia, but many data show that this is a subline of HeLa cells. 6 J111 cells were transfected with siRNAs against Axin1 or control siRNAs (final 25 nM unless otherwise stated) using HiPerfect transfection reagent (Qiagen), as described previously. 4 To confirm the specific RNAi effect of siRNA on Axin1 gene expression, two siRNAs targeting different positions in the Axin1 gene, Axin1 siRNA1 (target sequence: 5′-GAUACCUGCCGACCUUAAA-3′; RNAi Co., Tokyo, Japan) and Axin1 siRNA2 (#121445; Ambion, Austin, TX) were tested. In addition, siRNAs against Apaf-1 (#1024594; Qiagen, Hilden, Germany) and p53 (#16706, Ambion), as well as the control (nonsilencing) siRNA (#1022076, Qiagen), were used as negative controls.
We studied the late phase of HIV-1 replication by transfecting cells with the luciferase reporter proviral construct. Using this assay system, the efficiency of the late phase of the HIV-1 life cycle, including individual steps such as viral RNA transcription, translation, and posttranslational modification of viral proteins, virion assembly, and budding steps, can be monitored by measuring the level of HIV-1 Gag p24 antigen in the culture supernatant. In addition, we can monitor the level of viral gene expression by measuring the firefly luciferase activity in reporter proviral construct-transfected cells. Forty-eight hours after siRNA transfection, J111 cells were transfected with pNL4-37-derived proviral construct bearing the firefly luciferase gene in place of the viral nef gene, pNL-Luc-E-R+, 8 and the Renilla luciferase-expression vector, pTK-RL (Promega, Madison, WI), using Lipofectamine 2000 transfection reagent (Invitrogen). Forty-eight hours after transfection, the concentration of HIV-1 Gag p24 antigen in the cell culture supernatant was measured by enzyme-linked immunosorbent assay (Vironostika HIV-1 Antigen Microelisa System, bioMérieux, Boxtel, The Netherlands). In addition, the activity of firefly and Renilla luciferases in the transfected cells was measured using Dual-Glo luciferase assay system (Promega) with an LB960 microplate luminometer (Berthold, Bad Wildbad, Germany). Renilla luciferase activity was used to monitor transfection efficiency. The results showed that the level of the late phase of the viral replication cycle (Fig. 1A), as well as viral gene expression (Fig. 1B) was markedly higher in cells transfected with siRNAs against Axin1 than in cells transfected with control siRNAs, suggesting that siRNA against Axin1 enhances the late phase of the HIV-1 replication cycle by upregulating viral gene expression in J111 cells. In contrast, the levels of viral gene expression were comparable among HeLa cells transfected with siRNAs against Axin1 or control siRNAs (Fig. 1C). These results suggest a certain difference in the intracellular signal transduction pathway involving Axin1 between parental HeLa and its subline, J111 cells.
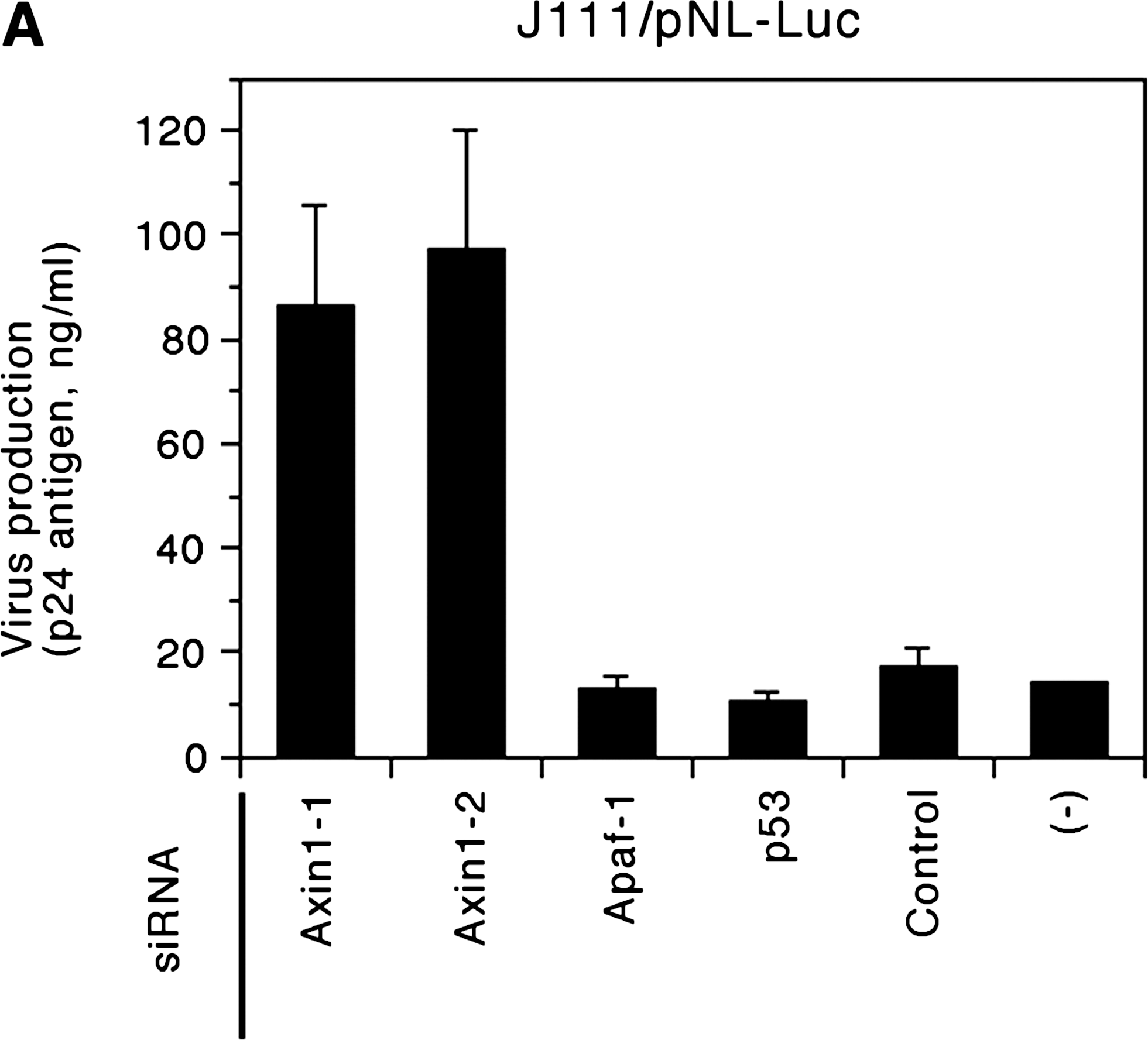
siRNAs against Axin1 enhance the late phase of HIV-1 replication by upregulating viral gene expression in J111, but not in HeLa cells. J111 (
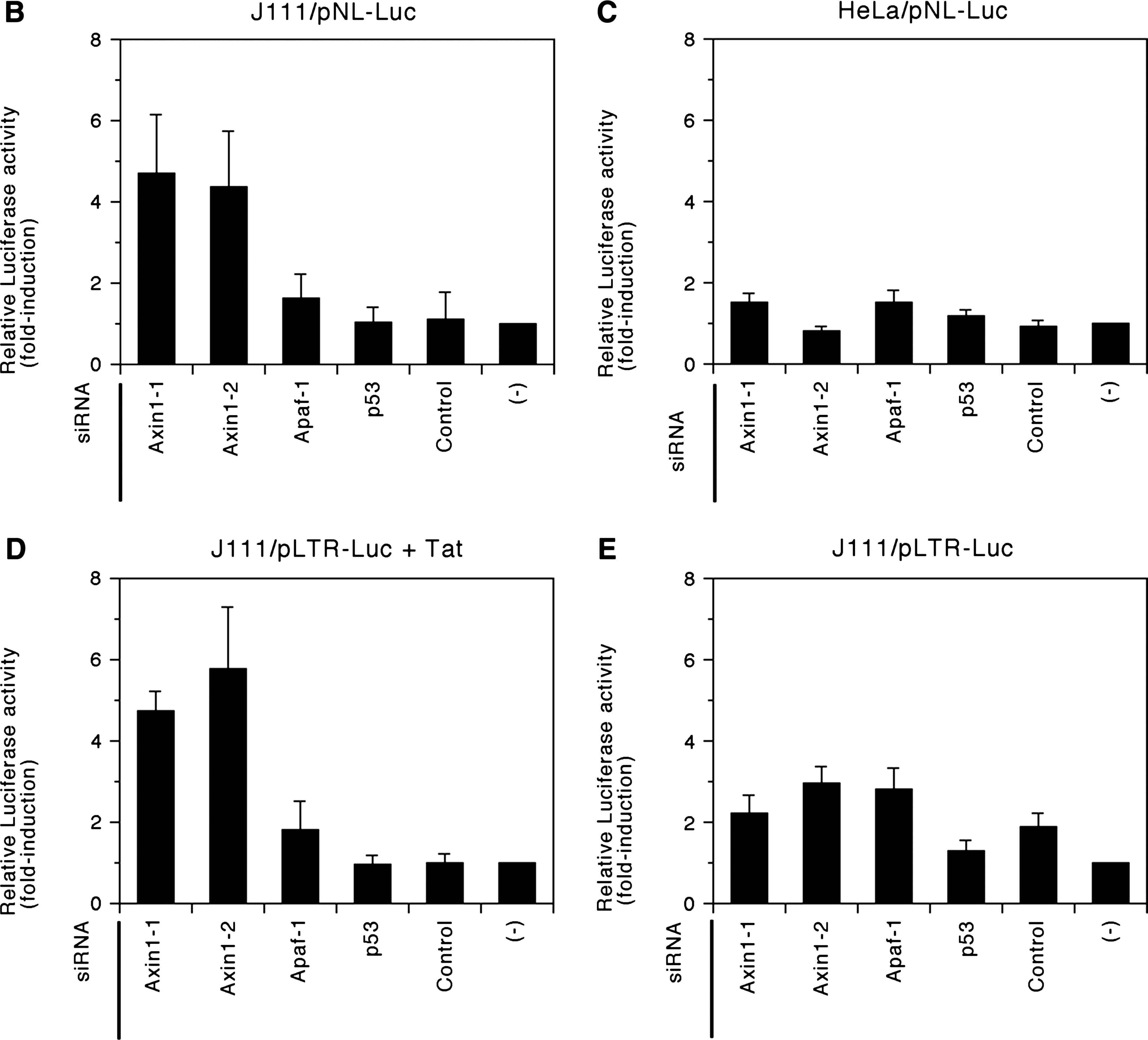
We further studied the HIV-1 gene expression in cells transfected with siRNA against Axin1 by reporter gene analysis. Forty-eight hours after siRNA transfection, cells were cotransfected with the HIV-1 promoter-dependent firefly luciferase reporter gene construct, pLTR-Luc, 9 pTK-RL, and the effecter plasmid including HIV-1 Tat expression vector, pRSV-Tat, and empty vector, pRc/RSV (Invitrogen). Forty-eight hours after transfection, the activity of firefly and Renilla luciferases in the cells was measured, as described above. The results showed that Tat-dependent viral reporter gene expression was higher in cells transfected with siRNAs against Axin1 than in cells transfected with control siRNAs (Fig. 1D). In contract, the level of Tat-independent, basal viral gene expression was comparable among cells transfected with siRNAs against Axin1 or control siRNAs (Fig. 1E). These results suggested that Axin1 was involved in the regulation of Tat-dependent viral gene expression. It is noteworthy that HIV-1 promoter-dependent firefly luciferase gene expression from pLTR-Luc was more than 100-fold enhanced by cotransfecting J111 as well as HeLa cells with pRSV-Tat (data not shown), demonstrating that the level of Tat-dependent viral gene expression was significantly higher than that of Tat-independent, basal viral gene expression.
To determine how Axin1 affects HIV-1 gene expression, we next semiquantified HIV-1 transcripts in cells transfected with siRNA against Axin1 by real-time PCR following reverse transcription of viral mRNA. Total cellular RNA was extracted from cells transfected with pNL-Luc-E-R+. An RNA sample was treated with RNase-free DNase (TaKaRa Shuzo, Kyoto, Japan), and then reverse transcribed into cDNA by SuperScript III reverse transcriptase (Invitrogen) either using oligo(dT)20 or the HIV-1-specific reverse primer, NL723AS, 5′-TTGCCGTGCGCGCTTCAGCAAGCC-3′ [nucleotide (nt) 723 to 700 of pNL4-3]. NL723AS-reverse transcribed cDNA was used to semiquantify HIV-1 transcripts, as follows. The cDNA sample was mixed with an 18-μl reaction mixture containing 2 × TaqMan universal PCR master mix (Applied Biosystems, Foster City, CA), HIV-1-specific primers RS, 5′-AACTAGGGAACCCACTGCTTAAG-3′ (nt 500 to 522) and U5AS, 5′-CGCCACTGCTAGAGATTTTCCAC-3′ (nt 640 to 618) (300 nM each), as well as the TaqMan RU5 probe, 5′-FAM-ACACTACTTGAAGCACTCAAGGCAAGCTTT-TAMRA-3′ (nt 559 to 530) (200 nM). Real-time PCR was then carried out using an ABI 7500 Fast Real-Time PCR system (Applied Biosystems). To monitor the efficiency of RNA extraction as well as cDNA synthesis, β-actin cDNA was amplified from the oligo(dT)20-reverse transcribed sample using TaqMan β-actin control reagents (Applied Biosystems). The plasmid, pNL-Luc-E-R+, was serially 10-fold diluted and subjected to real-time PCR to make a standard plot, and the copy number of HIV-1 transcripts was calculated using Sequence Detection Software version 1.3.1 (Applied Biosystems). The level of HIV-1 transcripts was standardized by dividing the copy number of HIV-1 transcripts by that of β-actin to compensate for variations in the quantity or quality of cDNA samples, and the results are shown as the relative amount of HIV-1 transcripts. The results showed that the level of viral transcripts was markedly higher in cells transfected with siRNA against Axin1 than in cells transfected with control siRNA (Fig. 2). The viral transactivator, Tat, plays a major role in HIV-1 transcriptional elongation. 10 In addition, Tat potentially regulates the initiation process of HIV-1 transcription. 11 Our results suggest that the signal transduction pathway involving Axin1 plays a role in the regulation of HIV-1 transcription in the presence of Tat.
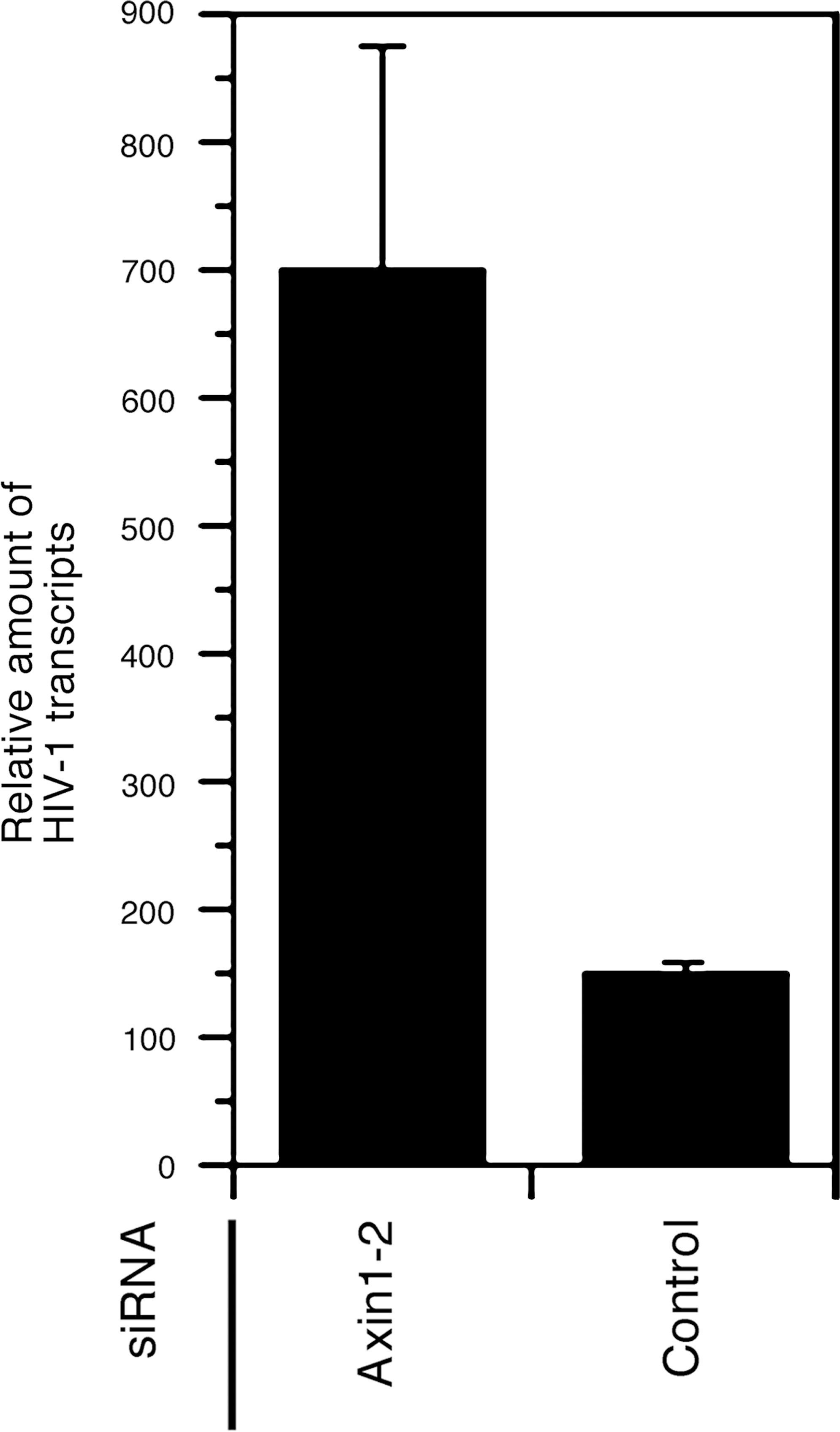
The level of HIV-1 transcripts is increased in cells transfected with siRNA against Axin1. J111 cells were transfected with Axin1 siRNA2 or control (nonsilencing) siRNA. Forty-eight hours after siRNA transfection, the cells were transfected with pNL-Luc-E-R+. Total cellular RNA was extracted 48 h after transfection, and treated with DNase. Then, cDNA was reverse transcribed from 1 μg of RNA, and subjected to real-time PCR using the primers/probe set for detecting viral transcripts. The level of HIV-1 transcripts was standardized by dividing the copy number of HIV-1 transcripts by that of β-actin, and the results are shown as the relative amount of HIV-1 transcripts. Data are presented as the means and standard deviations (error bars) of three independent experiments.
Hypophosphorylated β-catenin is accumulated in the cytoplasm under activated conditions of Wnt signaling. 5 Therefore, we determined whether the Wnt signaling pathway was activated in cells transfected with siRNA against Axin1 by measuring the level of phosphorylated β-catenin. In addition, we examined the RNAi effect of the siRNA against the regulators of Wnt signaling, including Axin1. For this purpose, immunoblot analysis was performed using the following antibodies: anti-Axin1 monoclonal antibody (MAB4327; Chemicon, Temecula, CA), anti-β-catenin monoclonal antibody (#610153; BD Biosciences, San Jose, CA), anti-phospho-β-catenin (Ser33/37/Thr41) polyclonal antibody (#9561; Cell Signaling Technology, Danvers, MA), anti-TCF4 polyclonal antibody (#2566; Cell Signaling Technology), and anti-β-actin monoclonal antibody (A5316; Sigma-Aldrich, St. Louis, MO). The results showed that the level of phosphorylated β-catenin was markedly reduced in cells transfected with siRNAs against Axin1, suggesting that the Wnt signaling pathway was activated in these cells (Fig. 3A, second panel). In addition, siRNAs against TCF4, β-catenin, and Axin1 specifically reduced the expression levels of target gene products (Fig. 3A).
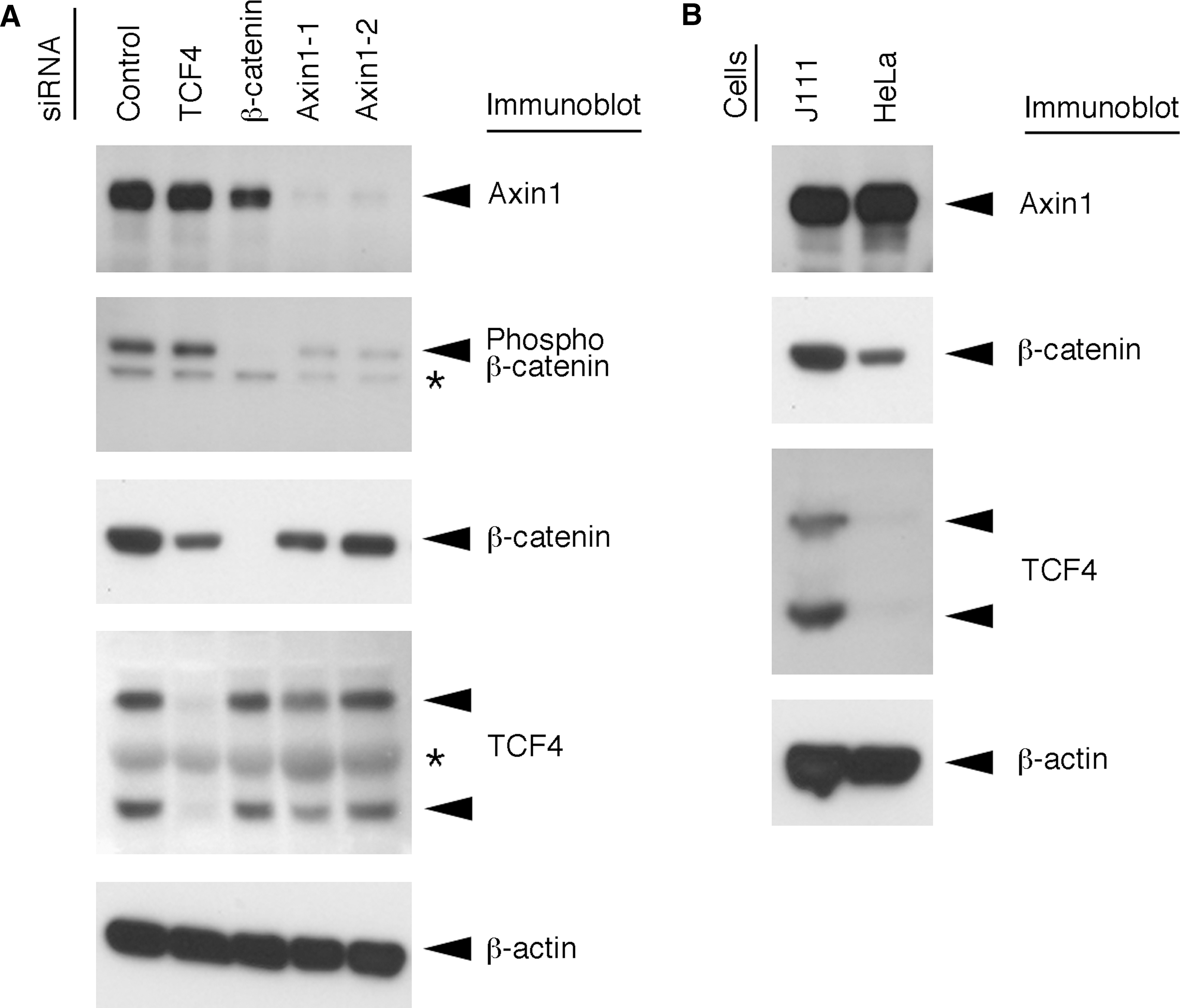
Immunoblot analysis of the regulators of Wnt signaling. J111 (
siRNA against Axin1 enhanced HIV-1 gene expression in J111 cells (Fig. 1B), but not in HeLa cells (Fig. 1C); therefore, we next compared the expression levels of the regulators of Wnt signaling between J111 and HeLa cells by immunoblot analysis. The results showed that the expression level of Axin1 was comparable between J111 and HeLa cells (Fig. 3B, first panel). In contrast, the expression level of β-catenin was higher in J111 cells compared with that in HeLa cells (Fig. 3B, second panel). Furthermore, TCF4 was strongly expressed in J111 cells, whereas it was weakly expressed in HeLa cells (Fig. 3B, third panel). These results suggested that β-catenin and TCF4 were required for Axin1 siRNA-dependent enhancement of HIV-1 gene expression. Therefore, we next studied the possible involvement of the regulators of Wnt signaling in Axin1 siRNA-dependent enhancement of HIV-1 gene expression using siRNAs against β-catenin (#42816, Ambion) and TCF4 (#116412, Ambion). The results showed that single transfection of cells with siRNA against β-catenin or TCF4 did not have any effect on the level of viral reporter gene expression (Fig. 4A). In contrast, cotransfection experiments revealed that siRNAs against β-catenin and TCF4 strongly inhibited the Axin1 siRNA-dependent enhancement of viral reporter gene expression (Fig. 4B). These results suggest that β-catenin and TCF4 are required for Axin1-mediated regulation of HIV-1 gene expression.

siRNAs against TCF4 and β-catenin do not affect HIV-1 gene expression in single transfection experiments (
Our results suggest that the β-catenin-dependent Wnt signaling, which is activated by transfecting cells with siRNA against Axin1, upregulates HIV-1 gene expression in HeLa-derived J111 cells. It was previously suggested that TCF4 plays a negative role in HIV-1 gene expression through the β-catenin-independent signaling pathway in astrocytic U87MG cells. 12 In addition, β-catenin signaling is suggested to inhibit HIV-1 replication in peripheral blood mononuclear cells. 13 There is a discrepancy among the results in the previous reports 12,13 and ours regarding the role of Wnt signaling in HIV-1 gene expression. This discrepancy is potentially due to the difference in cell types used in the experiments. Namely, unknown cell type-specific factors might be involved in the regulation of HIV-1 gene expression by Wnt signaling. Such cell-specific factors might affect the host cell tropism of HIV-1. In addition, such a discrepancy might be due to the difference in experimental methods used in these reports. Wnt signaling pathway was activated in previous reports by overexpressing TCF4 in the cells 12 or by treating cells with lithium chloride, an inducer of Wnt signaling, 13 whereas Wnt signaling was activated by transfecting cells with siRNA against Axin1 in this study. Different regulatory molecules are possibly involved in the signal transduction pathway activated by these different methods. Taken together, we propose that further studies are required to fully understand the regulatory mechanism of HIV-1 gene expression by the Wnt signaling pathway.
Footnotes
Acknowledgments
We are grateful to Dr. Yoshitake Nishimune (Research Institute for Microbial Diseases, Osaka University) and Dr. Panasda Isarangkura-na-ayuthaya (National Institute of Health, Department of Medical Sciences, Ministry of Public Health) for their valuable help with this study. We thank Dr. Hironori Sato [Division of Molecular Genetics, National Institute of Infectious Diseases (NIID), Tokyo, Japan], Dr. Jun Komano (AIDS Research Center, NIID), Dr. Naoki Yamamoto (AIDS Research Center, NIID), Dr. Yukukazu Natori (RNAi Co., Ltd, Tokyo, Japan), Dr. Masato Fujino (RNAi Co., Ltd), and Ms. Shinobu Oguchi (RNAi Co., Ltd) for their help in establishing the siRNA minilibrary. This work was supported in part by a Grant-in-Aid for Scientific Research (KAKENHI) from the Japanese Society for the Promotion of Science; the program of Founding Research Center for Emerging and Reemerging Infectious Diseases launched by a project commissioned by the Ministry of Education, Cultures, Sports, Science and Technology (MEXT) of Japan; and a research grant from the Department of Medical Sciences, Ministry of Public Health of Thailand. The manuscript was proofread by Medical English Service (Kyoto, Japan). RCC-ERI was established by the Research Institute for Microbial Diseases, Osaka University, Japan and the Department of Medical Sciences, Ministry of Public Health, Thailand.
Disclosure Statement
No competing financial interests exist.