
Abstract
In Brazil, where three distinct HIV-1 subtypes (B, F, and C) cocirculate, a significant portion of the HIV-infected population has been exposed to antiretroviral drugs. This study analyzes the antiretroviral resistance profiles of HIV-1-infected individuals failing antiretroviral therapy. Genotypic resistance profiles of 2474 patients presenting virologic failure to antiretroviral therapy in the city of São Paulo, Brazil, were generated and analyzed. Resistance mutations to protease inhibitors and nucleoside reverse transcriptase inhibitors were less common in subtype C viruses, whereas nonnucleoside reverse transcriptase inhibitor resistance mutations were less common in subtype F viruses. The thymidine analog mutation pathway known as pathway 1 was more prevalent in subtype B viruses than in subtype C viruses, whereas pathway 2 was more prevalent in subtype C viruses. Selected resistance mutations varied according to subtype for all three classes of antiretrovirals. We describe two distinct pathways of nonnucleoside reverse transcriptase inhibitor resistance (to nevirapine and efavirenz). Although cross-resistance to etravirine should occur more frequently among individuals failing nevirapine treatment, the prevalence of cross-resistance to etravirine, darunavir, and tipranavir was found to be low. We found that increases in the number of resistance mutations will be related to increases in the viral load. Special attention should be given to resistance profiles in non-B subtype viruses. The accumulation of knowledge regarding such profiles in the developing world is desirable.
Introduction
U
There are many unresolved issues regarding HIV-1 behavior in relation to its genetic diversity. It is conceivable that the genetic diversity of the virus impairs immune responses to candidate vaccines, causes false-negative results in diagnostic/monitoring laboratory tests (especially those involving nucleic acids), 4,5 and alters disease progression. 6,7 Most of the resistance mutation profiles described to date have been in subtype B HIV-1 viruses. However, in non-B subtypes subjected to the selective pressure exerted by antiretroviral therapy, specific resistance mutations or pathways can be selected. For instance, some thymidine analog mutations (TAMs), as well as some PI resistance mutations, are more frequently selected in subtype B viruses than in non-B subtypes. 8,9 Emerging in vitro data suggest that some natural polymorphisms in the pol gene interfere with antiretroviral efficacy. For instance, in subtype C viruses, the V106M mutation is related to lower activity of nonnucleoside reverse transcriptase inhibitors (NNRTIs), 10 and the I93L mutation increases susceptibility to lopinavir. 11 In addition, subtype C viruses, due to a silent polymorphism, also have a lower genetic barrier to tenofovir resistance. 12 In the present study, we sought to analyze the secondary resistance among HIV-1-infected individuals failing antiretroviral therapy in the city of São Paulo, Brazil in a cohort of patients for whom information related to previous antiretroviral exposure was available, with a special emphasis on how well subtype correlates with resistance and with resistance pathway.
Materials and Methods
Population
Samples from 2474 patients followed at the outpatient's clinics of the Federal University of São Paulo and at the São Paulo Municipal Health System were collected between 2002 and 2006. All participating patients gave written informed consent.
Viral load and number of mutations
The number of resistance mutations was calculated as the sum of all nucleoside reverse transcriptase inhibitor (NRTI), NNRTI, and major PI resistance mutations (minor PI resistance mutations were excluded). Means, standard deviations, and medians were calculated for viral load in each group of patients. As the best fit distribution for viral load was the lognormal distribution instead of the normal distribution, the ANOVA could not be used. The Mood's median test, a nonparametric alternative to ANOVA that tests the equality of medians from two or more populations, was performed. This test is more robust than the Kruskal–Wallis test against outliers and some extreme values. Furthermore, analyses were performed using the proportion test based on binominal distribution to determine the number of patients in each group presenting a viral load ≥100,000 copies/ml. All groups were compared to an antiretroviral-naive group of 194 individuals, in which 0.19% of the patients had a viral load ≥100,000 copies/ml. Values of p ≤ 0.05 were considered statistically significant.
Genotypic resistance analysis
Plasma samples from all patients were subjected to RNA purification, followed by PCR amplification and sequencing using ViroSeq v2.0 (Celera Diagnostics, Alameda, CA). Resistance mutations were identified according to the IAS mutations list. 13 Statistical analyses were performed using Minitab Release 14 Statistical Software.
The prevalence of mutations in subtype B viruses was compared with that observed for subtype F and C viruses using chi-square tests and tests of proportion based on a binomial distribution.
HIV-1 subtype analysis
The nucleotide sequences of the pol gene were subtyped using BLAST analysis (
Results
Patient characteristics
A total of 2474 consecutive patients were enrolled in this study. All were being treated with antiretroviral drugs, and all were experiencing virologic failure: 19.25% for the first time (19% for subtype B, 20.94% for F, and 21.7% for C—Chi-square nonsignificant); 23.72% for the second time (23.45% for subtype B, 25.21% for F, and 30.19% for C—Chi-square nonsignificant); or 57.03% for the third or subsequent time (57.55% for subtype B, 53.85% for F, and 48.11% for C—Chi-square p = 0.07 for differences between clade B and C). The mean viral load was 123,845 copies/ml.
Sample characteristics
Only 1.2% of strains presented no antiretroviral drug resistance mutations. The subtyping analysis showed that 78.3% (1938) of the viruses belonged to subtype B, 4.3% (106) belonged to subtype C, and 8.6% (214) belonged to subtype F. In addition, 0.5% (12) were B/C recombinants and 8.2% (204) were B/F recombinants. According to the analysis of individual fragments, the prevalence of protease and reverse transcriptase subtype B was, respectively, 79.6% and 85.8%, compared with 4.3% and 4.8% for subtype C and 16.1% and 9.5% for subtype F.
Genotypic resistance patterns
Mutations were identified in the following proportions: 54.3% (major PI resistance mutations), 89.7% (NRTI resistance mutations), and 62.5% (NNRTI resistance mutations). The prevalence of antiretroviral resistance by drug class and by HIV-1 subtype is presented in Table 1. Overall, 21% of individuals presented resistance to one antiretroviral class, 44.8% presented resistance to two classes, and 34% presented resistance to all three classes. As can be seen, there was a differential acquisition of resistance mutations by subtype, which was not explained by antiretroviral exposure (data not shown). Considering only major PI resistance mutations, the prevalence of PI resistance was lower in subtype C than in the other subtypes (Table 1). Additionally, subtype C viruses accumulated fewer NRTI resistance mutations than did subtype B or subtype F viruses. Subtype F viruses presented significantly fewer NNRTI resistance mutations than did subtype B viruses (Table 1).
PI mutations included in the analysis were L23I, L24I, D30N, V32I, V33F, M46I/L, I47V/A, G48V/M, I50L/V, F53F/V, I54V/T/A/L/M, G73S/T, L76L/V, V82A/F/T/S, I84V/A/C, N88T/S, and L90M. PI, protease inhibitor; NRTI, nucleoside reverse transcriptase inhibitor; NNRTI, nonnucleoside reverse transcriptase inhibitor; NS, not statistically significant.
Analysis of PI resistance mutations
Of the major PI resistance mutations, L90M, M46I/L, N88D/S, and D30N were the most frequently observed, whereas G48V and I50V/L were the rarest (Fig. 1A). Individuals failing nelfinavir as the first PI were evaluated. Among those infected with subtype B (n = 168) the following resistance pathways were identified: D30N in 52.5%, L90M in 38.3%, and the D30N–L90M combination in 9.6%. Among those infected with subtype F (n = 48), 30.4% presented the D30N pathway and 47.8% presented the L90M pathway. Using end-point PCR, we generated 10 clones from five individuals that presented the D30N–L90M combination and observed that the D30N or L90M mutations never coexisted in the same viral clone (data not shown). At the time of plasma sample collection from this group of patients, there had been no exposure to tipranavir or darunavir. Of the samples evaluated, 2.14% harbored three or more darunavir resistance mutations: V11I (1.18%), V32I (3.51%), L33F (3.55%), I47V (2.75%), I50V (1.07%), I54M (0.61%), L74P (0.8%), L76V (2.10%), I84V (7.44%), and L89V (1.49%). In addition, 22.15% of samples harbored three or more tipranavir resistance mutations: L10V (12.21%), I13V (30.46%), K20M/R (10.50%), L33F (6.49%), E35G (1.34%), M36I (49.43%), K43T (4.31%), M46L (5.73%), I47V (2.75%), I54A/M/V (2.60%), Q58E (5.31%), H69K (7.86%), T74P (2.21%), V82L/T (3.09%), N83D (0.84%), and I84V (7.52%). Of the individuals evaluated, 0.38% harbored viruses with three or more mutations associated with resistance to tipranavir and darunavir, 18.9% harbored viruses with three or more mutations associated with resistance to tipranavir, but not to darunavir, and 1.8% harbored viruses with three or more mutations associated with resistance to darunavir, but not to tipranavir.
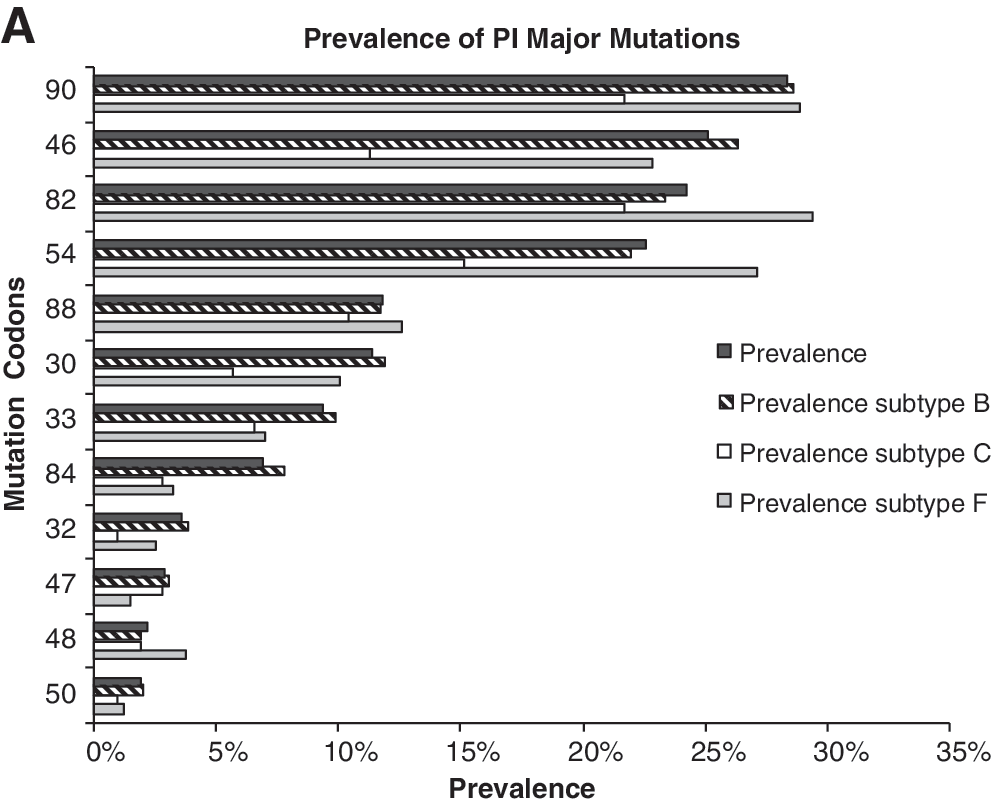
Prevalence of codons with antiretroviral resistance mutations according to HIV-1 subtype (B, F, and C). (
Although exposure to different PIs or to the number of previous PIs did not vary among the different subtypes, there were differences in the prevalence of primary and secondary PI resistance mutations. In subtype B viruses, resistance mutations at codons 33, 34, 58, 63, 73, 71, 77, and 84 were more common than in subtype F or C viruses, whereas those at codons 20, 36, and 89 were less common. In subtype F viruses, resistance mutations were more common at codons 10, 20, 35, 36, 48, 74, 57, 82, and 89, whereas they were less common at codons 47 and 93. In subtype C viruses, the frequency of resistance mutations was higher at codons 20, 36, 89, and 93 and lower at codons 10, 30, 43, 46, and 74 (p < 0.05 for all).
Analysis of NRTI resistance mutations
The NRTI resistance mutation M184V/I was present in 66.3% of patients (Fig. 1B), whereas 3.5% harbored viruses with multiple NRTI resistance mutations related to the Q151M complex and to the codon 69 insertion. The K65R mutation was found in only 1% of patients. The NRTI resistance selected by the TAMs M41L, D67N, K70R, L210W, T215Y/F, and K219Q/E was 70%. Of the individuals evaluated, 13.31% presented one TAM, 22.35% presented two TAMs, 17.09% presented three TAMs, 12.13% presented four TAMs, 2.48% presented five TAMs and 1.37% presented six TAMs.
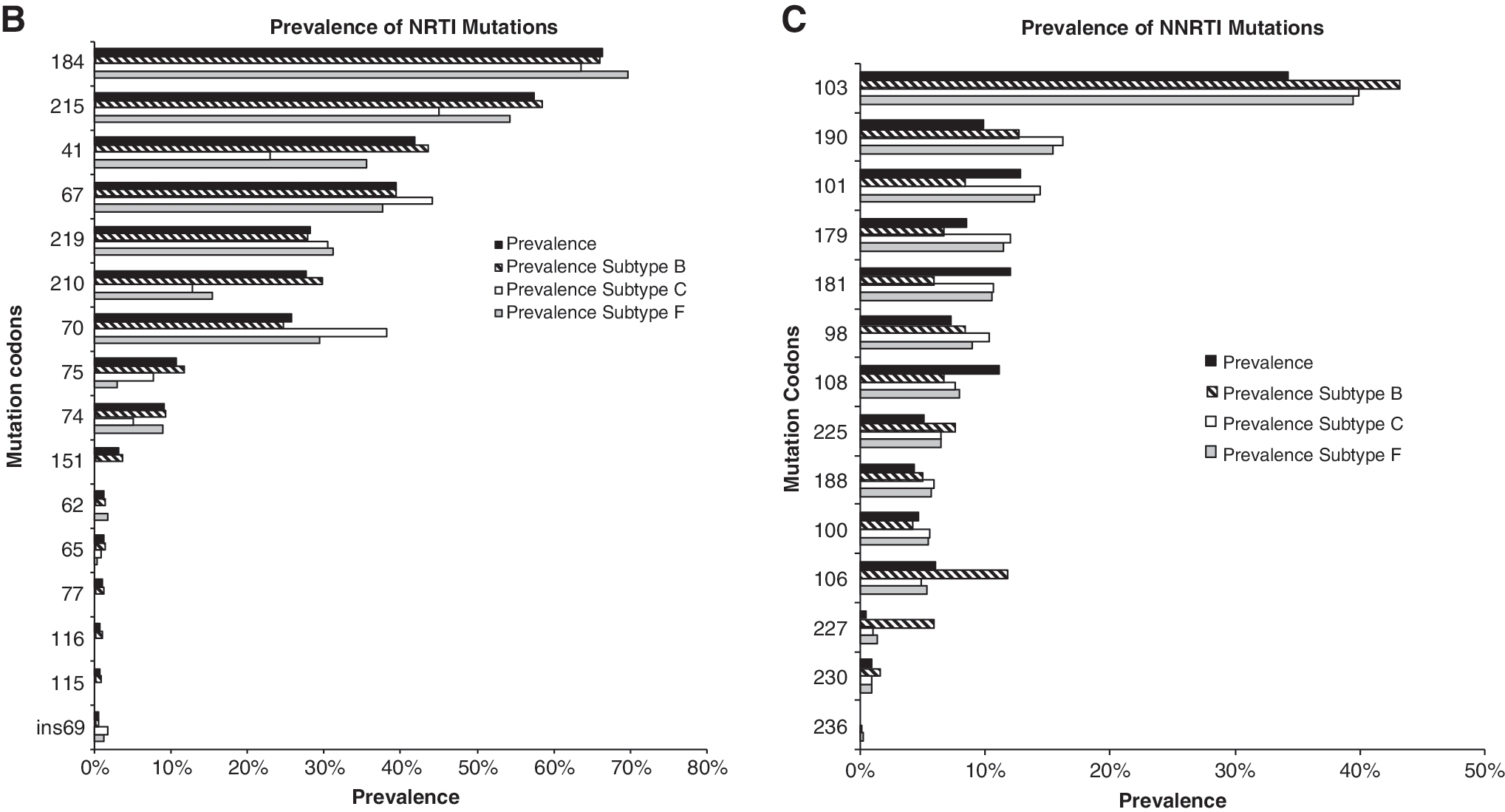
Two TAM pathways have been described, in which TAM pathway 1 includes mutations at codons 41, 210, and 215, whereas TAM pathway 2 includes mutations at codons 67, 70 and 219. 14 Interestingly the TAM pathway 1, which includes mutations at codons 41, 210, and 215, was most prevalent among subtype B viruses, whereas mutations at codons 67, 70, and 219 were most prevalent among subtype C and F viruses (Fig. 2).
Prevalence of TAMs by subtype. Pathway 1 TAMs are represented by reverse transcriptase codons 41, 210, and 215, and pathway 2 TAMs are represented by codons 67, 70, and 219. Statistical analyses were performed using chi-square tests, and p values are shown for each subtype comparison.
In subtype B viruses, resistance mutations other than TAMs were most often observed at codons 75 and 151; in subtype C viruses, such mutations were least common at codons 44 and 118 (p < 0.05 for all).
Analysis of NNRTI resistance mutations
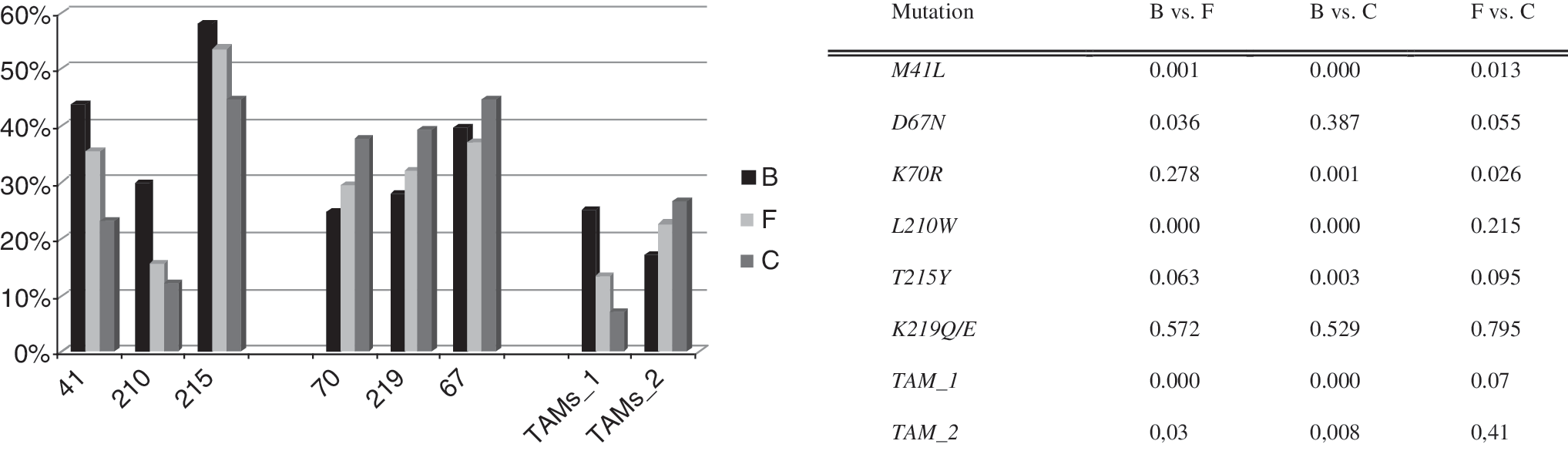
The prevalence of individual NNRTI resistance mutations is presented in Fig. 1C. An interesting new pathway of NNRTI resistance was detected in this study. Individuals failing efavirenz presented K103N as a key selected mutation, whereas those failing nevirapine presented the Y181C mutation (Fig. 3A). In addition, mutations L100I and P225H were strongly associated with the K103N mutation, whereas mutations K101E and G190A were strongly associated with the Y181C mutation (p < 0.000 for all). Therefore, we were able to describe two pathways of NNRTI resistance: one in which failing nevirapine is most often related to mutations Y181C, K101E, and G190A, and one in which failing efavirenz is most often related to mutations K103N, L100I, and P225H (Fig. 3B). Noticeable, these patients were not in the same period of virologic failure or the same line of therapy, and these resistance profiles were conserved among the different HIV-1 subtype. Potential cross-resistance to etravirine was evaluated, and 3.01% of the individuals presented three or more resistance mutations to this new antiretroviral drug: A98G (9.00%), L100I (5.40%), K101P/E/H (9.24%), V106I (5.30%), E138A (2.67%), V179E/F/I (9.84%), Y181C/I/V (9.36%), G190A/S (15.4%), and M230L (1.18%).
Analysis of 798 patients failing NNRTIs as the most recent treatment regimen. (
Resistance mutations at codon 106 were most prevalent in subtype C viruses, resistance mutations at codon 190 were less prevalent in subtype F viruses, and resistance mutations at codon 225 were more prevalent in subtype B viruses (p < 0.05 for all).
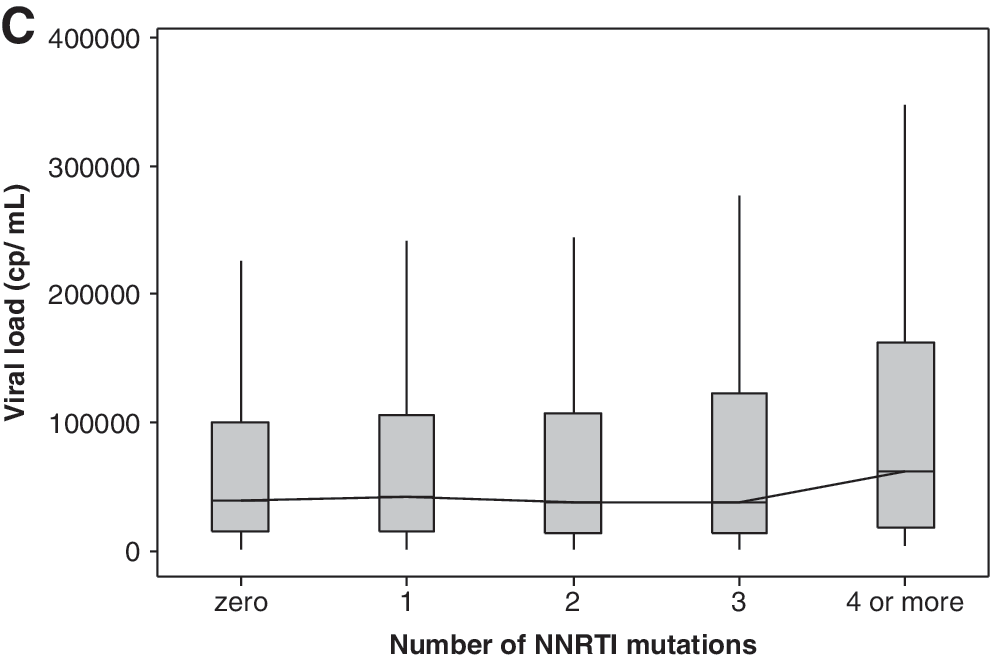
Viral load and number of resistance mutations
Figure 4 shows the mean viral load according to the number of resistance mutations accumulated during antiretroviral treatment. Initially, viral load decreased in parallel with the increase in the number of resistance mutations, after which it increased as the number of resistance mutations peaked and stabilized (Fig. 4). The viral load values are classified as being below or above the overall median, and a chi-square test for association was performed. The χ 2 value of 41.59 with a p-value = 0.000 for NRTI, PI, and NNRTI mutations indicates that there is at least one significant difference between the groups. The 95% confidence intervals of the individual group medians help to find where the significant difference is. If there is no overlap between the confidence intervals, a significant difference can be assumed. In this case, at least number of mutations 1 and ≥13 have significantly higher viral loads (Fig. 4A). In case of NRTI and PI mutations, the χ 2 value of 51.85 with a p-value = 0.000 also indicates significant differences among medians (Fig. 4B). The inspection of the 95 CI suggests number of mutation 0, 10, 11, and ≥12 as the groups with significant higher viral loads. Mood's median test showed no significant differences for NNRTI number of mutations (Fig. 4C). The results for all groups were also compared with those obtained for the treatment-naive group, which consisted of 194 patients with recent HIV-1 infection, as determined using the serologic testing algorithm for recent HIV seroconversion. 15 Viral loads analyzed for each individual patient in this group of naive patients were chosen after the viral load set point (6 months after inclusion in the cohort).

Box-plot showing median viral load values for each category according to the number of antiretroviral drug resistance mutations: number of NRTI, NNRTI, and PI resistance mutations (
Therefore, the proportion test based on binomial distribution was used to analyze the groups stratified according to the number of mutations in terms of the distribution of patients with viral loads greater than 100,000 copies/ml. When all resistance mutations were analyzed, the proportion of individuals with viral loads greater than 100,000 copies/ml was significantly higher in the groups with eight or more resistance mutations than in the treatment-naive group. There was also a higher proportion of individuals with viral loads greater than 100,000 copies/ml in the groups with at least one NNRTI resistance mutation than in the treatment-naive group.
Discussion
This study enabled us to profile genotypic resistance to antiretrovirals in a large group of patients failing antiretroviral therapy and from whom information related to previous treatment was available. It can be seen that the majority of individuals failing antiretroviral treatment in this region of Brazil, where genotyping was requested by the attending physician, had extensive antiretroviral exposure, and 50% were failing their third or subsequent antiretroviral regimen. This extensive antiretroviral exposure and failure, consequences of, among other factors, long-term treatment with suboptimal antiretroviral schemes, lead to extensive antiretroviral resistance. Of these patients, 34% harbored resistance to all three classes of antiretrovirals, a scenario that has been associated with a high risk of mortality. 16
In the present study, resistance to NRTIs revealed the extensive exposure to thymidine analogs and lamivudine. Rare resistance mutations related to didanosine (codon 74) and stavudine (codon 75) were also prevalent in our patient sample, with a prevalence of approximately 10%. In contrast to what have been reported in previous studies with smaller number of individuals, 17,18 we observed a differential prevalence of resistance mutations among HIV-1 subtypes without any apparent difference in the previous antiretroviral exposure. Subtype B viruses presented more NRTI resistance mutations than did subtype C or F viruses. Nonetheless, it is important to note that the estimation year for the introduction of HIV-1 subtype B in Brazil was 1970, for the subtype F was 1981, and for the subtype C was 1987, and, therefore, subtype B was theoretically more exposed to the selective pressure of mono/biotherapy and to unboosted PIs. 19
Another recent report also showed that among infected individuals failing antiretroviral therapy, those infected with subtype C viruses present less accumulation of resistance mutations to NRTIs than do those infected with subtype B viruses. 20
One reasonable explanation for this differential acquisition of antiretroviral drug resistance mutations is related to the current ignorance regarding the genotypic correlates of viral resistance among HIV-1 subtypes other than subtype B. Alternatively, and perhaps less likely, resistance to NRTIs in these non-B subtypes could be related to resistance mutations to other as yet uninspected HIV genomic regions, such as Rnase H. 21,22 A similar explanation could be given for the fact that NRTI resistance mutations were more frequently acquired in subtype B viruses than in subtype F viruses. We found less acquisition of protease mutations in subtype C viruses than in subtype B or F viruses. Indeed, previous studies have also produced the same results comparing subtype C and B only. 20 However, it must be borne in mind that some natural polymorphisms, such as the L89M substitution present in subtype F viruses, might be sufficient to decrease the susceptibility to a number of PIs, 23 perhaps underscoring the importance of using boosted PIs for the treatment of subtype F-infected individuals. 24
Identifying pathways of antiretroviral resistance is considered important for predicting cross-resistance to antiretrovirals. The term antiretroviral resistance pathway refers to the group of resistance mutations selected in vivo or in vitro by a given antiretroviral agent. One likely explanation for these resistance pathways is the genetic structure of the virus. This insight comes from the observation that subtype B viruses exposed in vivo or in vitro to nelfinavir will typically select the D30N mutation, whereas non-B viruses will select the L90M mutation. 25 Supposedly, the D30N mutation rarely coexists with the L90M mutation. In confirmation of this, we found that in the few cases in which these two resistance mutations occurred concomitantly, they arose in different genomes. We also found that the L90M mutation was present in subtype B-infected individuals at a higher proportion than that previously reported, 26 in 47.9% of our patients, including the viruses containing the D30N–L90M combination. This indicates that cross-resistance to PIs selected by nelfinavir was higher than expected in this population, since cross-resistance to other PIs is lower among D30N-containing viruses. In addition, and in contrast of what has been generally speculated, we found D30N mutations in 30.4% of the subtype F-infected individuals in our sample. 26
Another important pathway for resistance described more recently refers to the TAM profile of resistance mutations. The initial selection of TAMs has been shown to occur either within pathway 1, which includes resistance mutations at codons 41, 210, and 215, or within pathway 2, which includes resistance mutations at codons 67, 70, and 219. The practical implication of this is cross-resistance to tenofovir, where resistance mutations at codon 41 or codon 210 in a total of three TAMs will result in a marked decrease in the activity of this adenosine analog. 27 Although it has been demonstrated that pathway 1 and pathway 2 present an approximately equal risk of initiating resistance, 28,29 we found that both of these pathways correlated strongly with the viral subtype. Clearly, the prevalence of acquired resistance mutations related to pathway 1 would be higher among subtype B-infected individuals, whereas that of those related to pathway 2 would be higher among subtype C-infected individuals. In this sense, cross-resistance to tenofovir would be more prevalent in subtype B-infected individuals than in subtype C-infected individuals. Therefore, although the results of in vitro studies have suggested that the genetic barrier to tenofovir resistance is lower in subtype C viruses due to rapid selection of the K65R mutation, 12 we can speculate that salvage therapy with tenofovir is more effective among subtype C-infected individuals, especially in view of the possibility that K65R will not emerge in the presence of TAMs, since TAMs and K65R rarely coexist. 30,31 It is of note that in the present study, subtype F viruses were intermediate between subtype B and C viruses in terms of the pathway 1- and pathway 2-selected TAMs, suggesting that in subtype F viruses, the chance of pathway 1 TAMs being selected is approximately the same as that of pathway 2 TAMs being selected.
In the present study, we also detected two distinct pathways to NNRTI resistance, one related to the selective pressure of efavirenz and other to that of nevirapine. The most common mutation related to efavirenz resistance is the K103N mutation, whereas the most common mutation related to nevirapine resistance is the Y181C mutation. It is noteworthy that mutations K101E and G190A were associated with the Y181C mutation and with nevirapine exposure, whereas mutations L100I and P225H were associated with the K103N mutation and with efavirenz exposure. All three mutations associated with nevirapine exposure are related to etravirine cross-resistance, whereas among those associated with efavirenz exposure, only the L100I mutation is related to etravirine cross-resistance. Therefore, nevirapine failure might impair future salvage therapy with etravirine, which was designed to be active against NNRTI-resistant HIV strains. Nevertheless, cross-resistance to etravirine appears to be low (1.06% of samples presented three or more related mutations), and etravirine might be a good option for salvage therapy in this population (those failing other NNRTIs). Similarly, darunavir and tipranavir might also be good options for salvage therapy, as cross-resistance to both seems to be equally low in this population. Because strains resistant to darunavir will sometimes remain sensitive to tipranavir and vice versa, the availability of both drugs will be important in such cases.
A well-defined paradigm related to the accumulation of antiretroviral drug resistance mutations is the concomitant decrease in the replicative capacity of a virus, which will ultimately decrease viral fitness. It has been suggested that the presence of a resistant virus will lead to a more modest decrease in CD4 T cell counts, 32 as well as to less pronounced cell activation, 33 thereby potentially delaying disease progression. It has been established that the viral load can be a surrogate marker of viral fitness. 34 On the basis of this assumption, we attempted to draw a correlation between the number of resistance mutations accumulated and the viral load in order to infer the impact that the accumulation of resistance mutations has on viral fitness. We found that as the number of resistance mutations increases, there is in fact a drop in the mean viral load. However, after the number of resistance mutations reaches a certain threshold, the viral load begins to increase in direct proportion to the number of resistance mutations. We therefore speculate that the replicative capacity of the virus is restored after a virus has accumulated a large number of resistance mutations, and that this explains the more rapid disease progression seen in individuals presenting more extensive resistance. 16 Another possible explanation for viral load increasing in parallel with the number of resistance mutations is that since all of our patients were receiving antiretroviral therapy, there was less residual activity among those with more extensive resistance. Nevertheless, it is undeniable that the strains containing a high number of resistance mutations and producing high viral loads also present high fitness. Because the probability of accumulating antiretroviral drug resistance mutations over time is directly proportional to the magnitude of the residual viral replication in cases of treatment failure, 35 it would be reasonable to pursue a viral load that would be as low as possible, even in patients harboring resistant viruses, to minimize the chances of accumulating additional resistance mutations.
In conclusion, it is becoming clear that special attention should be given to the resistance profile selected in non-B subtypes, and further studies along this line are warranted.
Footnotes
Author Disclosure Statement
No competing financial interests exist.