
Abstract
It has recently been demonstrated that toll-like receptors (TLRs) can recognize structural conserved motifs carried by circulating microbial products and lead to systemic immune responses in individuals infected with HIV-1. TLRs have been detected in CD8+ T cells at either a protein or RNA level. The role of TLRs on CD8+ T cells involved in the host's immune responses during HIV-1 infection has not been well characterized. In this study, we analyzed expression of TLR4, TLR5, TLR7, and TLR8 in CD8+ T cells in HIV-1 infection. All these four TLRs could be detected in CD8+ T cells, but only TLR7 in CD8+ T cells from HIV-1-infected individuals showed a higher expression level compared with that from healthy individuals (p < 0.05). The function of TLR7 in CD8+ T cells was then investigated. We found that TLR7 ligand responsiveness significantly increased the expression of immune activation markers on purified CD8+ T cells in HIV-1-infected individuals compared with healthy controls. And the levels of these markers were equivalent to those achieved by CD8+ T cells from peripheral blood mononuclear cells (PBMCs). However, we also observed that TLR7 ligand stimulated significant IFN-γ production by CD8+ T cells in an accessory cell-dependent manner. Therefore, although CD8+ T cells can be directly activated by TLR7, accessory cells must play an essential role in the activation of effective functions such as IFN-γ production. These findings suggest that the abnormal expression of TLR7 in CD8+ T cells from HIV-1-infected individuals may contribute to the abnormal immune activation in HIV-1 infection and play an important role in HIV-1 pathogenesis.
Introduction
H
Toll like receptors (TLRs) have emerged as one of the major components of the immune system. Recognition of pathogen-associated molecular patterns (PAMPs) by TLRs results in the activation of signaling events that induce the expression of effector molecules such as cytokines and chemokines, which control the activation of adaptive immune responses. 9,10 TLR4, either associated or not associated with CD14, identifies various ligands such as lipopolysaccharide (LPS), heat shock proteins, and fibronectin. Circulating LPS was significantly increased in chronically HIV-1-infected individuals and in simian immunodeficiency virus (SIV)-infected rhesus macaques due to the microbial translocation through the virus-linked compromised gastrointestinal mucosal surface. Increased blood levels of the TLR4 ligand LPS have been found in chronic HIV-1 infection and these levels directly correlate with immune activation and inversely with CD4+ T cell restoration after antiretroviral therapy. 11 TLR5 detects bacterial flagellin and can be cross-linked with its ligand to activate CD4+ and CD8+ T cells. TLR7/8 recognizes imidazoquinolines and single-stranded viral RNA. Triggering TLR7/8 stimulates the innate as well as the adaptive immune responses, leading to an increase of antiviral defense. 11 Recent reports have demonstrated that natural killer cells (NK cells) and CD8+ T cells are the main effector cells in TLR7/8-mediated anti-HIV-1 activity. 12 TLR ligands have been shown to dramatically modulate the function of dendritic cells, 13 –16 monocytes, 17,18 as well as B and T cells, 19,20 suggesting that stimulation through TLRs can directly activate and modulate effector cells of both the innate and adaptive immune system.
Materials and methods
Study individuals
Thirty-one HIV-1-infected individuals (median age, 42 years; range, 22–66 years; 20 male and 11 female) and 20 healthy volunteers (median age, 40 years; range, 24–60 years; 12 male and 8 female) enrolled in the study at Tangdu Hospital in China. Informed consent was obtained from participants in accordance with the guidelines for conduction of clinical research at the Fourth Military Medical University and Tangdu Hospital. All investigational protocols were approval by the Fourth Military Medical University and Tangdu Hospital institutional review boards. Among the HIV-1-seropositive individuals, 18 were receiving highly active antiretroviral therapy (HAART) and 13 patients were treatment naive. Laboratory values for patients who were receiving HAART were as follows: CD4+ T cell count, 287–806 × 106 cells/liter, CD8+ T cell count, 440–1069 × 106 cells/liter, and HIV-1 RNA load, 50–21,250 copies/ml.
For patients who were not receiving antiretroviral therapy, laboratory values were as follows: CD4+ T cell count, 13–951 × 106 cells/liter, CD8+ T cell count, 267–2342 × 106 cells/liter, HIV-1 RNA load, 753–8,815,000 copies/ml. Peripheral blood samples were obtained when there was no indication of opportunistic infection. Individuals who had acute or chronic diseases and were using any drug were excluded from the healthy volunteers. The interval between the times at which blood samples were obtained and analyzed (<30 min) was the same for HIV-1-infected individuals and healthy volunteers. Samples from HIV-1-infected individuals and healthy volunteers were always handled in parallel.
Cell purification
For CD8+ T cell purification, peripheral blood specimens were incubated with a tetrameric antibody complex mediating cell enrichment from whole blood (Rosette Sep Human CD8+ T cell Enrichment Cocktail, Stem Cell Technologies Inc.), then separated by Ficoll-Hypaque density gradient according to the manufacturer's protocol. An additional positive selection was performed by magnetic cell sorting using MACS CD8+ enrichment MicroBeads and MS Mini MACS Columns (Miltenyi Biotec). The purity of the CD8+ T cells was analyzed by staining with fluorochrome-conjugated antibodies of CD3, CD14, and CD19 (BD Biosciences) and subsequent four-color flow cytometric analysis. The CD8+ T cell purity was above 99% and free of detectable CD14+ monocytes or CD19+ B cells for all experiments performed. At the same time, peripheral blood mononuclear cells (PBMCs) were separated from each whole blood sample by Ficoll-Hypaque (Sigma) density gradient centrifugation.
Quantitative real-time polymerase chain reaction (PCR)
Relative RNA quantification was performed by using PCR standard protocols and premixed PCR reagents (TaKaRa). Briefly, total RNA was extracted from approximately 1 × 106 cells using Trizol reagent (Invitrogen) and reversely transcribed into cDNA in accordance with the manufacturer's directions (TaKaRa). Real-time PCR was then performed using the LightCycler system (Roche). Oligonucleotides were used as follows: human TLR4 (NM 138554) sense 5′-AGGATGATGCCAGGATGATGTC-3′ and antisense 5′-TCAGGTCCAGGTTCTTGGTTGAG-3′; human TLR5 (NM 003268) sense 5′-CAGTATTTGAGGTGGCCTGAGGA-3′ and antisense 5′-GTTGCTACAGTTTGCAACGGAATG-3′; human TLR7 (NM 016562) sense 5′-CAATTGCTTCCGTGTCATCCAG-3′ and antisense 5′-TCCCTATGGAACCCAGAAGCAG-3′; human TLR8 (NM 016610) sense 5′-AAACTTGACCCAACTTCGATACCTAA-3′ and antisense 5′-GATCCAGCACCTTCAGATGAGG-3′; glyceraldehydes-3-phosphate dehydrogenase (GAPDH) (NM 002046) sense 5′-GCACCGTCAAGGCTGAGAAC-3′ and antisense 5′-TGGTGAAGACGCCAGTGGA-3′. All samples were performed in triplicate, and relative expression of TLR4, TLR5, TLR7, and TLR8 was determined by normalizing to GAPDH to calculate a fold change in value.
Gene expression was calculated using the comparative delta-delta-Ct method. 21 Briefly, each sample was assayed for the number of PCR cycles required to cross the threshold of the linear range of the reaction (C t). The amplification efficiencies of the target gene and reference gene were approximately equal. The three C t values for each sample were averaged, whereas the difference between C t values was below 1. Samples were normalized for the total template amount by subtracting the average GAPDH C t from the average TLR4/TLR5/TLR7/TLR8 C t (ΔC t). The normalized gene expression (ΔC t) of the healthy control was then subtracted from the normalized expression of HIV-1-infected individuals (ΔΔC t). The relative gene expression of TLR4/TLR5/TLR7/TLR8 in CD8+ T cells from HIV-1-infected individuals in relation to healthy control cells was calculated: 2−(ΔΔCt).
CD4+ and CD8+ T cell counting
The CD4+ and CD8+ T cell counts were determined by analyzing peripheral blood samples using flow cytometry. In brief, blood specimens were collected by venipuncture into evacuated tubes containing K3 EDTA anticoagulant and performed the test of CD4+ and CD8+ T cell subsets within 6 h. CD4+ and CD8+ T cell subsets in whole blood were quantified using three-color immunofluorescent staining with the following labeled monoclonal antibodies: anti-CD3PerCP, anti-CD8PE, and anti-CD4FITC (BD) according to the manufacturer's instructions (BD) and the tube was incubated at room temperature for 20 min in the dark. Red blood cells were lysed by the addition of 450 μl 1× FACS lytic solution (BD) for 15 min in the dark. Then cell counts were analyzed with Multiset software on FACSCalibur (BD).
Immunofluorescent staining for TLRs
TLR7 expression was assessed using fluorochrome-conjugated antihuman TLR7 antibodies (Abcam) for flow cytometry. In brief, 500,000 freshly isolated CD8+ T cells were stained with anti-CD8APC for 30 min in the dark at 4°C. After incubation, 2 ml of fixative medium (1% paraformaldehyde) was added to cells and incubated in a dark environment at room temperature for 10 min. Following incubation, 500 μl of permeabilization medium (perm B, Caltag Lab Inc.) was added to the cells. After intracellular staining for 30 min with rabbit antihuman TLR7 antibodies, the cells were incubated with goat antirabbit IgG-FITC for 30 min in the dark at 4°C and resuspended for flow cytometric analysis.
Intracellular cytokine staining
Secretion of interleukin (IL)-2 and interferon (IFN)-γ was determined using intracellular cytokine staining, while expression of CD69, CD38, and HLA-DR was determined using extracellular staining. Then 1 × 106 CD8+ T cells or PBMCs in 1 ml of complete RPMI 1640 containing 10% fetal calf serum were stimulated with 0.5, 1, and 5 μg/ml CL097 (InvivoGen). In every experiment, cells incubated without CL097 were used as negative control. Cells stimulated with 20 ng/ml PMA plus 1 μg/ml ionomycin served as positive control. The cultures were incubated for 1 h at 37°C in 5% CO2 followed by an additional 6 h or 20 h incubation with 10 μg/ml of the secretion inhibitor Brefeldin A (Sigma). Cells were first stained with Abs of anti-CD8APC, anti-CD69PE, anti-CD38FITC, and anti-HLA-DRPE (BD) for surface markers. After fixation and permeabilization (Fix perm A and B, Caltag Lab Inc.), cells were intracellular stained with anti-IFN-γFITC and anti-IL-2PE. For analysis, 100,000–150,000 cells were acquired on a FACSCalibur flow cytometer (BD Biosciences) and FACS data were analyzed using Cellquest software.
HIV-1 viral load testing
Plasma HIV-1 viral loads were analyzed by real-time quantifying PCR assay (Roche Corporation) according to the manufacturer's instructions.
Statistical analysis
Unpaired two-tailed student t-tests were employed to assess the statistical significance of differences. Relations between variables were tested using the Spearman rank correlation test. p-values of less than 0.05 were considered significant.
Results
CD8+ T cells in HIV-1-infected individuals express high levels of TLR7 mRNA
To examine the expression levels of TLR4, TLR5, TLR7, and TLR8 during HIV-1 infection, we sorted CD8+ T cells from HIV-1-infected individuals and healthy volunteers, then evaluated TLRs-specific mRNA in CD8+ T cells by quantitative reverse transcriptase (QRT)-PCR. The results showed that mRNA of all four TLRs could be detected across both HIV-1-seropositive and -seronegative populations. The mRNA expression of TLR 4, 5, and 8 was equivalent between HIV-1-infected and -uninfected subjects (Fig. 1A, B, and D), whereas the expression of TLR7 was significantly increased in freshly isolated CD8+ T cells from HIV-1-infected subjects compared with healthy controls (p < 0.05) (Fig. 1C), Thus, increased expression of TLR7 in CD8+ T cells appears to be associated with HIV-1 infection. We demonstrate that the expression of TLR7 mRNA in CD8+ T cells in HIV-1-infected individuals increased significantly for the first time.

Upregulation of TLR7 mRNA expression in CD8+ T cells from HIV-1-seropositive individuals. Total RNA was isolated from ex vivo purified CD8+ T cell from both HIV-1-seropositive and -negative individuals. Relative expressions of TLR4, 5, 7, and 8 were determined by real-time fluorescence quantitative PCR. C t means the number of PCR cycles required to cross the threshold of the linear range of the reaction. Samples were normalized for the total template amount by subtracting the average GAPDH C t from the average TLR4/TLR5/TLR7/TLR8 C t (ΔC t). ΔC t has an inverse relation with the expression of TLR7. The significant differences in TLR7 mRNA expressions are represented as p values (*p < 0.05). Data are mean ± SEM.
CD8+ T cells in HIV-1-infected individuals express high levels of TLR7 protein
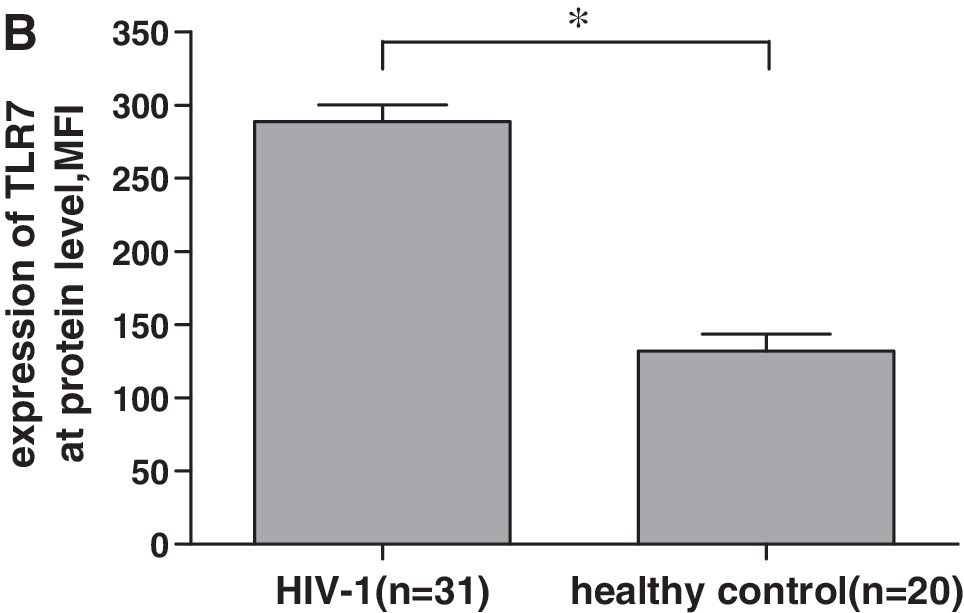
To further confirm the enhanced expression of TLR7 mRNA in CD8+ T cells in HIV-1-infected individuals, we examined TLR7 expression at the protein level in CD8+ T cells by flow cytometric analysis using intracellular staining. Freshly isolated CD8+ T cells from HIV-1-infected individuals had significantly higher TLR7 protein expression than those from healthy volunteers (Fig. 2). The mean fluorescence intensity of CD8+ T cells stained with antihuman TLR7 antibodies was 131.95 ± 52.38 for healthy controls compared with 288.81 ±63.11 for HIV-1-infected individuals (p < 0.05). These data reinforce the result of QRT-PCR, and demonstrate, for the first time, an increased TLR7 protein expression in CD8+ T cells from HIV-1-infected individuals, which suggests that overexpressed TLR7 in CD8+ T cells during HIV-1 infection might play some role in HIV-1 pathopoiesis or immunoprotection.
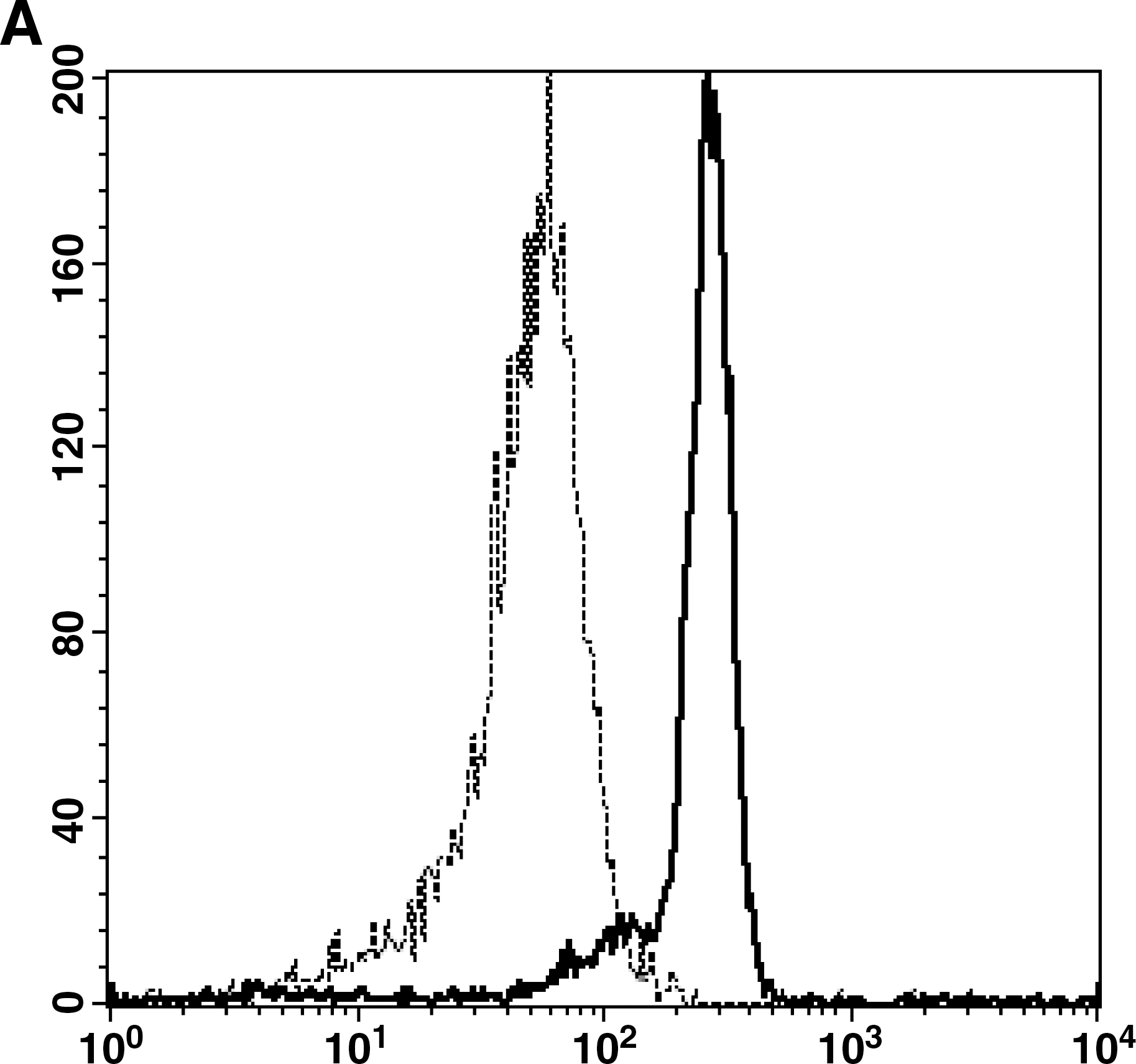
Upregulation of TLR7 protein expression in CD8+ T cells from HIV-1-seropositive individuals. TLR7 protein expression was assessed using fluorochrome-conjugated antihuman CD8 and TLR7 antibodies (Abcam) to stain freshly isolated CD8+ T cells and then subjected to flow cytometric analysis. (
Next, we determined the association between peripheral CD4+ T cell counts or plasma HIV-1 RNA levels and expression of TLR4, TLR5, TLR7, and TLR8. We found no statistically significant correlations between the expression of these TLRs and CD4+ T cell counts or HIV-1 viremia. Furthermore, when dividing patients into groups according to receipt of antiretroviral therapy, different CD4+ T cell counts (>200 cells/μl and <200 cells/μl), and different HIV-1 RNA levels (>105 copies/ml and <105 copies/ml), no differences in TLR4, TLR5, TLR7, and TLR8 expression were observed.
Ligand for TLR7 increases expression of activation markers on CD8+ T cells from individuals with HIV-1 infection
It has been demonstrated that some cell surface molecules such as CD69 are upregulated following T cell activation and differentiation. To assess whether overexpressed TLR7 could increase immune activation in HIV-1-infected individuals, we analyzed CD69 expression on CD8+ T cells, which is considered an early activation marker after exposure to tested stimuli, after being stimulated with TLR7/8 agonist CL097. Although CD69 expression on CD8+ T cells was significantly increased in both HIV-1-infected and -uninfected individuals after stimulation of purified CD8+ T cells with CL097, the increase in CD69 expression was significantly more marked in the HIV-1-infected group (p < 0.05). Since we have demonstrated that there was no significant difference of TLR8 expression between HIV-infected and -uninfected individuals, TLR7 should be responsible for CD69 overexpression in HIV-1-infected group. Furthermore, we evaluated whether CL097 of different concentrations (0.5, 1, and 5 μg/ml) could have different effects on CD69 expression. It was demonstrated that the maximal stimulating effect of CL097 occurred at a concentration of 0.5 μg/ml, as shown in Fig. 3. In addition, we detected the expression of other activation markers, including CD38 and HLA-DR. These molecules on CD8+ T cells were also upregulated after TLR7 agonist stimulation (data not shown). This result indicates that TLR7 signaling has a general effect on CD8+ T cell activation.

TLR7 agonist triggered CD8+ T cell activation. CD8+ T cell activation was determined by upregulation of CD69 expression in response to CL097. Then 1 × 106 purified CD8+ T cells or PBMCs from HIV-1-infected subjects and healthy controls in 1 ml of complete RPMI 1640 were stimulated with 0.5, 1, and 5 μg/ml CL097 (InvivoGen) for 6 h in a 5% CO2 incubator. Cells were then stained with Abs of anti-CD8APC and anti-CD69PE (BD) for surface markers and then subjected to flow cytometric analysis. Data were analyzed using Cellquest software (BD). (
To further assess whether the activation of CD8+ T cells by CL097 depended on the presence of accessory cells, we evaluated the CD69 expression of CD8+ T cells from HIV-1-infected individuals following stimulation of PBMCs with CL097 (0.5, 1, and 5 μg/ml). As expected, treatment of PBMCs with CL097 resulted in a significant increase of CD69 expression (Fig. 3). However, no significant difference of CD69 expression was observed between CD8+ T cells from PBMCs and highly purified CD8+ T cells in response to CL097 stimulation. Our results suggest that the TLR7 ligand might change the activation status of CD8+ T cells from HIV-1-infected individuals in an accessory cell-independent manner.
TLR7 ligand induced IFN-γ production by CD8+ T cells from individuals with HIV-1 infection in an accessory cell-dependent manner
To examine whether the ex vivo differences in TLR7 expression could have functional consequences, we analyzed the effect of CL097 at concentrations of 0.5, 1, and 5 μg/ml on IL-2 and IFN-γ production of CD8+ T cells from HIV-1-infected individuals and healthy controls by intracellular cytokine staining. After 6 h and 20 h of incubation with CL097, purified CD8+ T cells alone from two groups produced neither of the cytokines. We subsequently studied cytokine secretion of CD8+ T cells following stimulation of PBMCs with CL097. After 6 h of incubation, CD8+ T cells from two groups hardly produced IFN-γ and IL-2. However, we observed that CL097 upregulated IFN-γ production significantly by CD8+ T cells from HIV-1-infected individuals and healthy controls after 20 h of incubation, but still failed to modulate IL-2 production. Moreover, IFN-γ production was significantly more marked in the HIV-1-infected group (p < 0.05). As shown in Fig. 4, the effects of CL097 on IFN-γ production were dose dependent and maximal at a concentration of 1 μg/ml. These data suggest that the TLR7 ligand can induce significant IFN-γ secretion in CD8+ T cells in an accessory cell-dependent manner during HIV-1 infection. Further work needs to be done to determine what kinds of cells take part in cytokine secretion in CD8+ T cells via the TLR7 pathway.
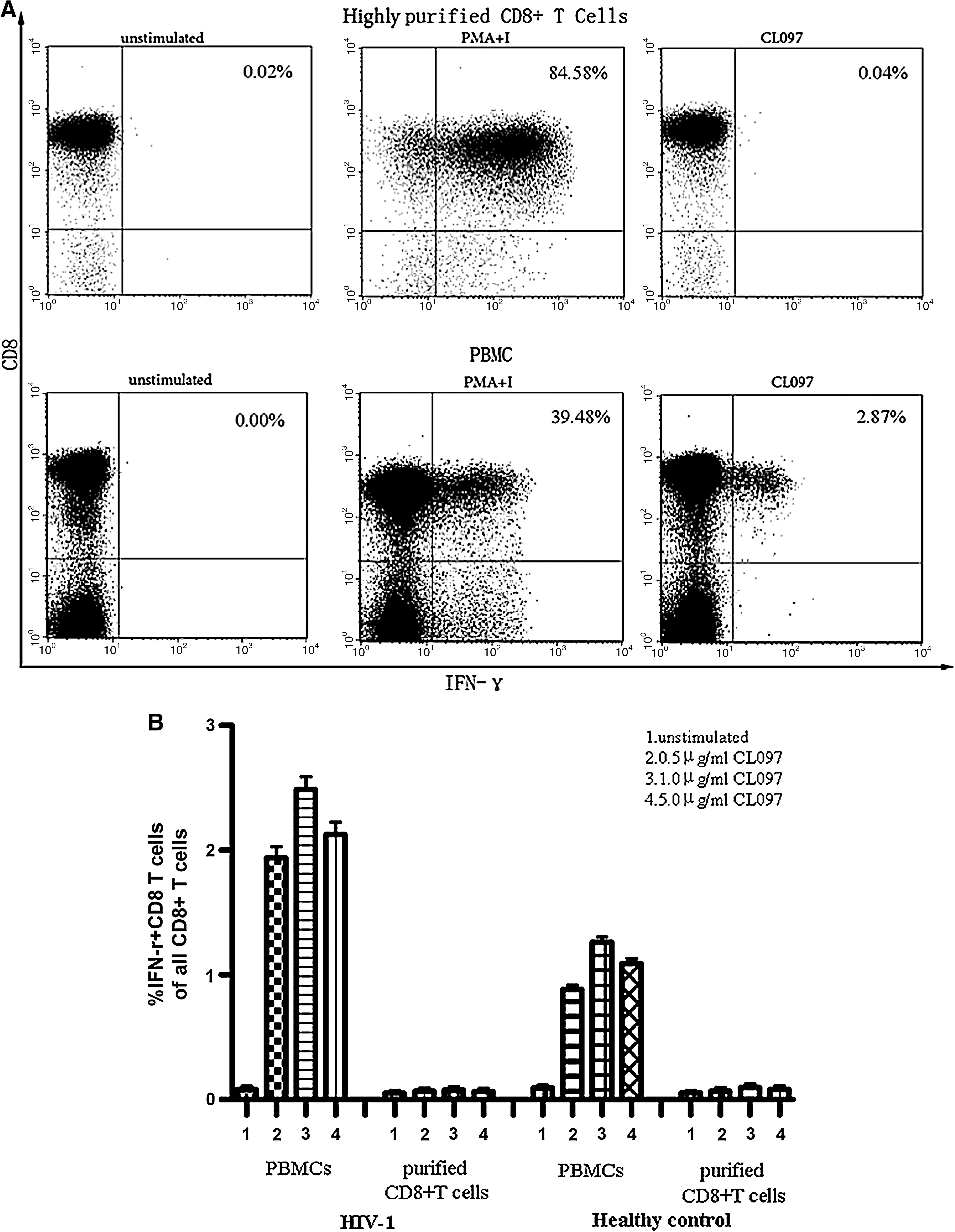
CL097 triggered IFN-γ production by CD8+ T cells in an accessory cell-dependent manner. Purified CD8+ T cells or PBMCs from HIV-1-infected subjects and healthy controls were treated with CL097 at various concentrations. IL-2- and IFN-γ-secreting CD8+ T cells were measured at 20 h poststimulation, respectively, by intracellular staining and flow cytometric analysis. CD3+CD8+ T cells were gated first. (
Discussion
TLRs are evolutionarily conserved molecules that play a key role in the initiation of innate antimicrobial immune responses. Through their influence on dendritic cell maturation, these receptors are also thought to indirectly shape the adaptive immune response. Most investigations on TLR expression and functions have initially mainly focused on cells of the innate system, as these cells are the first line of defense against pathogen infection. Although innate immunity may constitute the primary functions of TLRs, it is becoming increasingly clear that TLR can also be expressed by adaptive immune cells such as T lymphocytes, and that they can modulate the functions of these cells. 16,22 –25 However, no data are currently available regarding both TLR expression and function on CD8+ T cells in HIV-1 infection. Considering the important role of CD8+ T cells in controlling HIV-1 viral spread and replication, we investigated the expression of TLR4, TLR5, TLR7, and TLR8 in CD8+ T cells and assessed whether TLR7 ligand can directly modulate the function of CD8+ T cells in HIV-1 infection.
Previous studies reported that TLR2, TLR3, TLR4, TLR5, TLR7, TLR8, and TLR9 have been detected in CD8+ T cells at either the protein or RNA level. 26 –29 . In agreement with these studies, we observed that human CD8+ T cells could express TLR4, TLR5, TLR7, and TLR8 at both RNA and protein levels. Importantly, we demonstrated an increased TLR7 expression in CD8+ T cells during HIV-1 infection. These data are, to our knowledge, the first demonstration that CD8+ T cells express high level TLR7 during HIV-1 infection. A recent study of 200 HIV-1-infected women showed that increased expression of TLR7 in PBMCs appears to be associated with plasma HIV-RNA load. 30 However, when HAART, peripheral CD4+ T cell counts, or plasma HIV-1 levels were associated with TLR7 expression in CD8+ T cells, we found no clear impact among them. This discrepancy may be due to the study methods and sample size.
Several observations suggest a crucial role of chronic immune activation in HIV pathogenesis. 31 –35 The precise mechanism by which HIV causes immune activation remains poorly understood. Why is a small fraction of cells infected, while a general immune activation is present? T cell recognition of HIV antigens, Nef and gp120-mediated signaling, 36,37 and regulatory CD4+ T cell depletion 38 have all proposed to be major contributors to immune activation. In addition, “microbial translocation” of gastroenteric flora or its components into the circulation has been described as a cause of systemic immune activation in HIV, 19 and probably signals through TLR. However, most studies focused on innate immune cells, and demonstrate the role of TLRs from innate immune cells in immune activation during HIV infection. 39 –41
Recently, it has been demonstrated that TLR engagement increases the expression of activation markers on CD4+ T cells and CD8+ T cells. 42,43 Here, we found that CD8+ T cells could express activated markers by being stimulated with TLR7 agonist. Furthermore, this method of activation could be amplified in HIV-1-infected subjects through a higher level of TLR7 expression than in healthy controls. It has been demonstrated that the components of HIV-1 itself can induce strong immune activation via TLR7 and TLR8, which is associated with the level of HIV-1 viremia. 44 Therefore, these results provided a new method by which HIV-1 induces abnormal immune activation.
These findings suggest that CD8+ T cell activation in HIV-1 infection can be partially attributed to TLR7 triggering. Further studies will be needed to evaluate the contribution of immune activation of CD8+ T cells via TLRs pathway in HIV infection. In addition, we observed that there was no significant difference between the level of CD69 on purified CD8+ T cells and CD8+ T cells in PBMCs from HIV-1-infected individuals following stimulation with the TLR7 ligand. Our results thus suggest that TLR7 engagement may change the activation status of CD8+ T cells from HIV-1-infected individuals in an accessory cell-independent manner and may provide insights as to how systemic exposure to TLR ligand induces immune activation in HIV-1 infection. This finding reinforces recent data showing that in addition to indirect effects mediated by innate immune cells, TLR ligands may also have the capacity to directly regulate T cells. 45
Triggering TLRs causes complex changes in the cellular microenvironment, including release of cytokines, cellular activation, and upregulation of the MHC. 46 One of the main consequences of TLR7-induced signaling in cells of the innate immune system is the secretion of cytokines, such as interferons, which are antiviral cytokines. A previous study showed that CD8+ T cells are the main effector cells for anti-HIV-1 activity after triggering TLR7. 12 Although CL097 is sufficient to increase expression of CD69 on CD8+ T cells during HIV-1 infection, our study shows no effect of CL097 on IFN-γ and IL-2 production by highly purified CD8+ T cells.
Recent studies reported that R-848, an agonist of TLR7/8, directly enhances IFN-γ production by CD4+ T cells. 45 However, in accordance with our data, Meier et al. also failed to detect a direct effect of TLR7 ligands on IFN-γ and IL-2 production by CD8+ T cells. 44 The absence of effect of CL097 may be related to the absence of accessory molecules. This hypothesis is confirmed by additional experiments that showed CL097 could induce IFN-γ secretion of CD8+ T cells following stimulation of PBMCs from HIV-1-infected individuals.
Several studies demonstrated that TLRs expressed by adaptive immune cells such as T cells can modulate the function of these cells by enhancing cytokine production, such as IFN-γ. Our data suggest that TLR7-related mechanisms could induce an antiviral immune response via CD8+ T cells. Moreover, the CD8+ T cell effective function of cytokine production in response to TLR7 ligand depends on the presence of accessory cells. An understanding of the precise mechanism of the activation of CD8+ T cells and how CD8+ T cells fulfill their function via TLR7 in viral infection will require further investigation.
In summary, our data demonstrate an increased expression of TLR7 in CD8+ T cells in HIV-1 infection. TLR7 ligand can induce TLR7-mediated activation and accessory cell-dependent IFN-γ production of human CD8+ T cells. Our data suggest that the abnormal expression of TLR7 in CD8+ T cells from HIV-1-infected individuals may contribute to the abnormal immune activation in HIV-1 infection and play an important role in HIV-1 pathogenesis.
Footnotes
Acknowledgments
We thank all the subjects who agreed to participate in this study. This work is supported by a grant from the National Basic Research Program (Project 973) of the Ministry of Science and Technology of the People's Republic of China (Grant 2006CB504204), a Grand Program on Key Infectious Disease 2008ZX10001-002 project 2, and a grant from the health research program of the Health Department in Shaanxi Province (Grant 06D44). Y.S. and Y.Z. contributed equally to this work.
Disclosure Statement
No competing financial interests exist.