
Abstract
Reports indicate that myeloid and plasmacytoid dendritic cells (mDCs and pDCs), which are key effector cells in host innate immune responses, can be infected with HIV-1 and are reduced in number and function during the chronic phase of HIV disease. Furthermore, it was recently demonstrated that a sustained loss of mDCs and pDCs occurs in SIV-infected macaques. Since loss of functional DC populations might impair innate immune responses to opportunistic microorganisms and neoplastic cells, we explored whether inoculation of naive and SIV- or SHIV-infected pigtailed macaques with the hematopoietic cytokine FLT3-ligand (FLT3-L) would expand the number of mDCs and pDCs in vivo. After the macaques received supraphysiologic doses of FLT3-L, mDCs, pDCs, and monocytes increased up to 45-fold in blood, lymph nodes, and bone marrow (BM), with DC expansion in the BM preceding mobilization in blood and lymphoid tissues. FLT3-L also increased serum levels of IL-12, at least transiently, and elicited higher surface expression of HLA-DR and the activation markers CD25 and CD69 on NK and T cells. During and after treatment of infected animals, APCs increased in number and were activated; however, CD4+ T cell numbers, virion RNA, and anti-SIV/SHIV antibody titers remained relatively stable, suggesting that FLT3-L might be a safe modality to expand DC populations and provide therapeutic benefit during chronic lentivirus infections.
Introduction
D
Studies on mDCs and pDCs typically focus on cells in blood, but both DC subsets in various stages of development are common in murine, macaque (Macaca), and human bone marrow (BM), their site of origin. 12 –15 DCs also accumulate in respiratory mucosae following allergic stimuli and in cerebrospinal fluid during bacterial meningitis, induce antiviral responses in the vaginal mucosa, and can be isolated from human tonsils, thymus, lungs, liver, kidney, colon, and peripheral lymph nodes (LNs). 16 –24 Reduction in the numbers and functions of circulating mDCs and pDCs can occur during viral infections, such as with hepatitis B and C viruses, human T cell leukemia virus, SARS coronavirus, and HIV-1. 25 –31 We and others showed that DC numbers are reduced in blood and secondary lymphoid organs of rhesus (Macaca mulatta) and pigtailed macaques (M. nemestrina) acutely and chronically infected with SIV. 32 –34 Characterization of pDCs in BM of normal pigtailed macaques also revealed that while these cells are less mature phenotypically than their blood counterparts, they were responsive to microbial stimuli. 15,35
Because of their importance in initiating immune responses to pathogens and Ags in vaccine formulations, developing novel strategies to mobilize, by activation and/or maturation, DC populations in vivo is a valid goal. CD34+ myeloid and lymphoid progenitor cells have high levels of Fms-like tyrosine kinase 3 (FLT3) receptors on their cell surfaces. 36,37 The ligand for FLT3 (FLT3-L) is a cytokine expressed in most tissues, but it is at the highest levels on peripheral blood mononuclear cells (PBMCs). A soluble isoform of FLT3-L is also present in human blood at concentrations typically between 50 and 100 pg/ml. 38 FLT3-L is elevated in serum from patients undergoing chemotherapy, and abnormally high levels are associated with hematologic disorders and autoimmune diseases. 39 –41 Initially, murine studies showed that 9 days of subcutaneous inoculation of FLT3-L increased functional splenic DCs up to 17-fold. 37,42,43 After human stem cell transplantation, administration of FLT3-L to patients elicited increases in DC numbers and enhanced recovery of DC and NK cell subsets. 44 –46 FLT3-L-induced DC expansion, with subsequent emigration from BM, was first performed in macaques using a 10-day regimen that efficiently led to activated and increased numbers of DCs in blood, 47 but Teleshova et al. 48 showed that a 7-day regimen was equally effective. Similar experiments using a chimeric FLT3 and granulocyte colony-stimulating factor (G-CSF) receptor agonist, progenipoietin-1 (ProGP), were performed in naive and SIVmac32H-infected macaques. 49 ProGP induced as much as a 50-fold expansion of blood DCs, but this increase was insufficient to augment responses to HIV Ags elicited by DNA vaccination and to protect macaques against challenge with SHIV-89.6P. 50 In contrast, Kwissa et al. 51 inoculated macaques with FLT3-L for 14 days followed by immunization, either alone or with CpG-B DNA, a TLR9 ligand, with plasmid DNA (recombinant vaccinia virus Ankara, rMVA) expressing both SIV and HIV proteins. Their results demonstrated that by day 14 of FLT3-L treatment, activation enhanced expression of CD86 on mDCs and monocytes and, after boosting twice with rMVA, the number of CD8+ T cells expressing IFN-γ increased. This regimen did not prevent infection in all immunized groups after rectal SIVmac251 challenge, but viremia was reduced at 2 and 24 weeks in the best responders, those that received CpG-B DNA and FLT3-L.
Although blood DCs can be activated and expanded during cytokine administration, previous studies did not clarify the extent to which mobilization occurred in macaque tissues, especially in BM where DC progenitors develop from CD34+ stem cells. 47 –50 Since DCs play important roles in both innate and adaptive immunity, loss and/or impairment of these cells could exacerbate immune dysfunction observed during lentivirus disease. To determine whether targeting DCs as a way to enhance their numbers and functional maturity was a viable therapeutic strategy, we examined the kinetics of FLT3-L-induced activation and migration of mDCs and pDCs in blood, BM, and secondary lymphoid tissues in naive and SIV/SHIV-infected macaques.
Materials and Methods
Animals and FLT3-L administration
Adult pigtailed macaques of both sexes were used and included four uninfected macaques, four macaques infected for a mean duration of 213 weeks with SHIV-89.6P, 52 and three macaques infected for a mean duration of 191 weeks with one of two SIVmac239-derived mutants: SIVmac239ΔGY(S>P) (macaques CT19 and 98P016) or SIVmac239-AIRPA (99P032). 53 Macaque 98P016 was dually infected with SIVsmE660 for 109 weeks; reverse transcriptase polymerase chain reaction (RT-PCR) showed that SIVmac239ΔGY(S>P) predominated in plasma, with SIVsmE660 detected only sporadically (P.N. Fultz, Q. Wei, M. Piatak, J. Lifson, and J. Hoxie, unpublished observations). Animals were inoculated subcutaneously with sterile human recombinant FLT3-L, a gift from Amgen Corporation, for 7 consecutive days (100 μg/kg/day). Before each procedure (Table 1), macaques were anesthetized with an intramuscular injection of ketamine-HCl (10 mg/kg), weighed, their physical conditions were evaluated, and blood was collected. Necropsies were performed and tissues were taken after euthanasia with an intravenous overdose of pentobarbital (Fatal Plus; Vortech Pharmaceuticals). Macaques were housed in BSL2 isolation facilities at UAB in accordance with institutional and Animal Welfare Act guidelines. All procedures were approved by the UAB Institutional Animal Care and Use Committee.
Days, relative to the first inoculation (day 0) of FLT3-L, on which blood was obtained and bone marrow (BM) and peripheral lymph nodes (PLNs) were biopsied.
Day of necropsy.
P indicates pretreatment biopsies that were performed between 1 and 4 weeks before FLT3-L was first administered.
Tissue collection and processing
Heparinized blood was collected from the femoral vein; PBMCs were isolated by density gradient centrifugation through lymphocyte separation media (ICN Biomedicals, Inc.). BM was obtained from the medullary cavity of the proximal humerus with 15-gauge BM aspiration needles (Medical Device Technologies) previously flushed with heparin; yellow fat layers were removed from BM in EDTA-treated tubes. LNs and spleen tissue samples were minced and single-cell suspensions were generated by passing the fragments through metal cell strainers. Sections of the superior lobes of lungs were dissected, minced, and digested in medium supplemented with 0.5 mg of collagenase/ml at 37°C for 1 h, then passed through a cell strainer. At necropsy 6-cm sections of jejunum were removed, opened lengthwise, and rinsed with phosphate-buffered saline (PBS). The epithelial layer was removed and incubated in medium containing 0.5 mg of collagenase/ml at 37°C for 20 min. Digested samples were passed through a cell strainer, suspended in media, and passed over two glass wool columns to remove residual epithelial cells. Contaminating red blood cells were lysed hypotonically using an ammonium chloride solution. Mononuclear cells (MCs) were resuspended in RPMI-1640 with 10% fetal bovine serum (FBS(, or were cryopreserved in a dimethyl sulfoxide/FBS solution in liquid nitrogen vapor.
Flow cytometry
Three- and four-color flow cytometry were used to enumerate percentages of different cell populations and to characterize macaque cells in EDTA-treated whole blood or single-cell suspensions of BM, spleen, and LN tissues; cryopreserved MCs were used in some experiments. Lymphocyte subsets were phenotyped as follows: T cells were identified using Pacific Blue-conjugated anti-CD3 (SP34-2) with subsets distinguished by PE-conjugated anti-CD4 (SK3) and PerCp-conjugated anti-CD8 (SK1); NK cells were identified as CD3−CD8+ and stained with PE-conjugated anti-CD16 (3G8) or anti-NKG2A (Z199; Immunotech). Activation markers were evaluated using FITC-conjugated anti-CD25 (2A3) or anti-CD69 (FN50). After identifying HLA-DR+, with PerCp-labeled anti-HLA-DR (G46-6), and lineage-negative (Lin−) populations using an FITC-conjugated antibody cocktail to Lin markers—CD3ɛ (SP34), CD14 (MφP9), CD16 (3G8), and CD20 (2H7)—pDCs were distinguished with PE-labeled anti-CD123 (7G3) and mDCs by substituting PE-labeled anti-CD11c (S-HCL-3) and replacing CD16 in the FITC-labeled Lin cocktail with anti-CD8 (SK1). The latter was done because CD8 excludes most NK cells and macaque mDCs sometimes express low levels of CD16. 32 Coreceptor and activation/maturation molecules were evaluated using anti-CD40 (5C3) or anti-CD86 (2331) antibodies, both of which were conjugated to allophycocyanin, or biotin-conjugated anti-CD80 (L307.4) and streptavidin-allophycocyanin. Isotype-matched antibody controls for each fluorochrome were always included. MCs were gated based on forward-and-side-scatter characteristics. All antibodies and reagents were purchased from BD Biosciences Pharmingen, unless otherwise noted; acquisitions were performed on a BD-LSRII.
Quantification of SIV viral RNA and SIV antibodies
SIV virion RNA (vRNA) in plasma was quantified at the Quantitative Molecular Diagnostics Core of the AIDS Vaccine Program, SAIC Frederick, NCI (Frederick, MD), as described previously (sensitivity, <100 copies/ml). 54 Serum SIV-specific antibody titers were determined using a cross-reactive HIV-2 enzyme immunoassay kit (Bio-Rad).
Quantification of cytokines
FLT3-L concentrations in serum were determined using a Quantikine ELISA kit (R&D Systems) with a lower limit of detection of 7 pg/ml. Concentrations of IFN-α, tumor necrosis factor (TNF)-α, and interleukin (IL)-12p40 in serum samples were determined using cross-reactive human cytokine ELISA kits (Biosource); lower limits of detection were 10, 2, and 4 pg/ml, respectively.
Results
Hematologic changes associated with FLT3-L administration
Although FLT3-L upregulates hematopoiesis and cell proliferation in mice and humans, studies with macaques have provided limited data beyond specific myeloid or lymphoid subsets. 42,45 –48 Previous studies did not examine the effects that subcutaneous inoculation of supraphysiologic doses of FLT3-L had on serum levels of this cytokine in normal macaques. On the day macaques first received FLT3-L (day 0), serum concentrations ranged from 18.1 to 170 pg/ml, with no obvious differences between infected and uninfected animals (Fig. 1A). However, by days 3 to 5 serum levels increased as high as 1370 pg/ml (macaque 97P009) and in individual animals the increase ranged from 7- to 44-fold (day 0 concentration to the highest amount). Serum FLT3-L in most animals rapidly declined after the last inoculation (day 6), with two exceptions: infected macaques CT19 and AV1C maintained high levels that were at or above 1000 pg/ml on day 12.
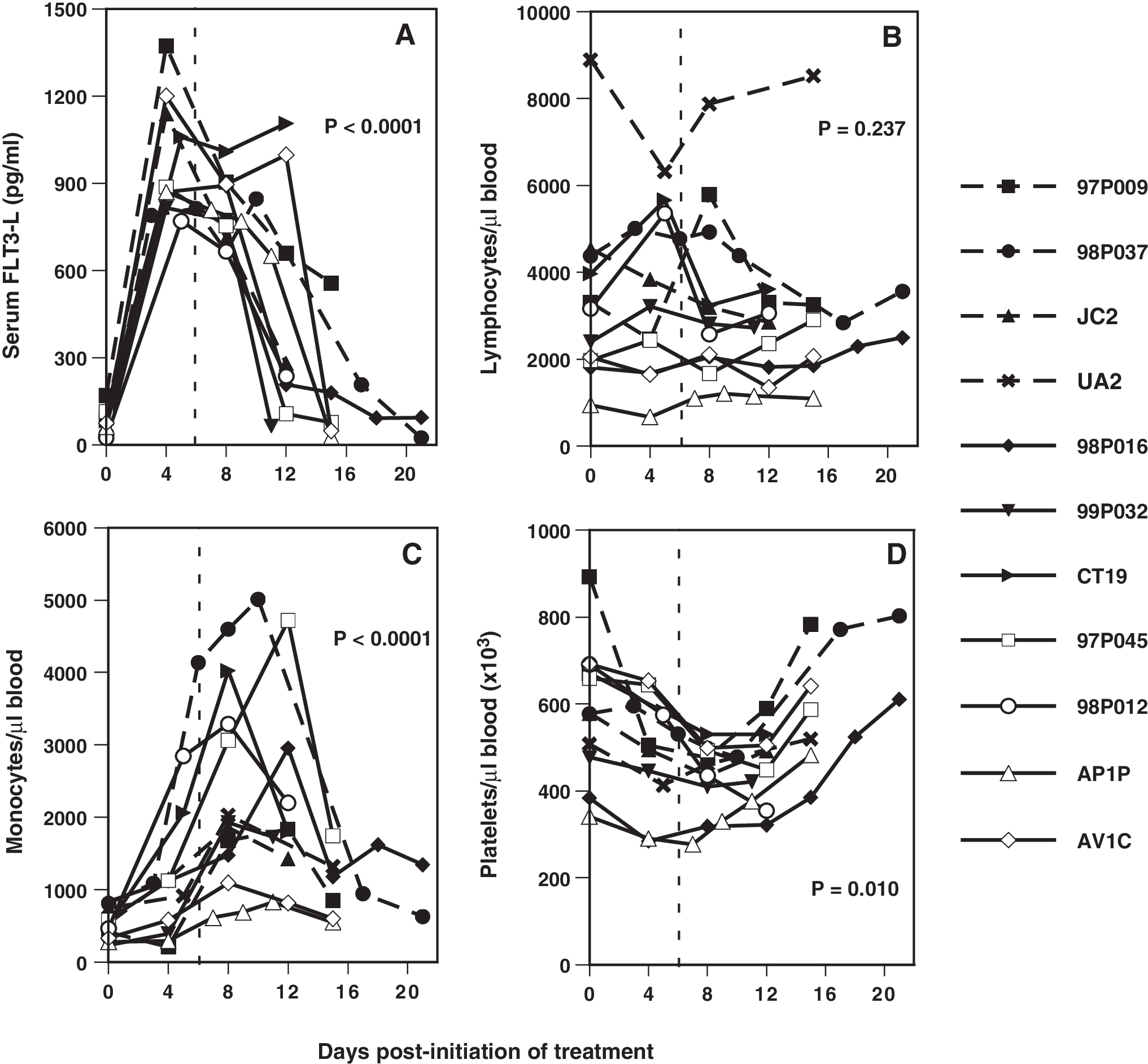
Hematologic parameters during and after administration of FLT3-L to macaques. (
The numbers of circulating lymphocytes (B, T, and NK cells), monocytes, and DCs were determined using complete blood counts and differentials and percentages of each population obtained by FACScan. The total numbers of blood lymphocytes before inoculation of FLT3-L appeared lower in infected animals compared to naive macaques: mean cell numbers/microliter for naive and SIVmac239- and SHIV-89.6P-infected macaques were 5279, 2558, and 2033 lymphocytes, respectively. Whether day 0 lymphocyte numbers from the naive outlier UA2 were included or excluded from its cohort, the mean number of lymphocytes in the naive group was statistically greater (Mann–Whitney U test), but only when compared to the means for the SHIV-89.6P-infected animals (p = 0.029) or for SIVmac239- and SHIV-89.6P-infected animals combined (p = 0.012, UA2 included; p = 0.033, UA2 excluded). Lymphocyte counts in most infected animals were relatively stable during FLT3-L treatment, whereas total lymphocytes in all naive animals fluctuated (Fig. 1B).
Numbers of blood monocytes in infected compared to control animals were marginally lower on day 0, but this population increased in all groups on days 6 to 12 (Fig. 1C). Because thrombocytopenia is a common hematologic manifestation of HIV- and SIV-induced disease, 55 –57 platelet numbers were also evaluated. Before treatment, numbers of blood platelets had a large range in the SIVmac239- and SHIV-89.6P-infected macaques, but none was thrombocytopenic. Combining results from all animals, when day 0 platelet numbers were compared with nadirs that occurred between days 8 and 12, the observed transient decrease was significant (p < 0.001, Wilcoxon test).
Changes in predictors of disease progression in SIV- and SHIV-infected macaques
Because the FLT3-L-induced changes in numbers of PBMCs might influence virus replication in the infected animals, vRNA in plasma was quantified. Most animals had initial viral loads at or below the limit of detection; these levels changed marginally during or after treatment, with three exceptions (Fig. 2A). Macaques CT19 and 97P045 developed transient viremias on days 5 and 8, respectively. Macaque 98P016, infected with both SIVmac239ΔGY(S>P) and SIVsmE660, had 1900 copies of vRNA/ml on day 0, which then increased to 11,000 copies/ml on day 15, before decreasing to pretreatment levels. All SIVmac239- and SHIV-89.6P-infected animals had viral antibody titers that ranged from 400 to 409,600 (reciprocal serum dilutions); no significant changes in titers from those on day 0 were observed during or after FLT3-L treatment, which is consistent with minimal changes in viral loads. Loss of CD4+ T cells is a hallmark of progressive lentiviral disease; before FLT3-L treatment the mean numbers of CD4+ T cells in pigtailed macaques chronically infected with SIVmac239 mutants (702 /μl) or SHIV-89.6P (462 cells/μl) were lower than in naive controls (1499 cells/μl). Comparison of the medians of CD4+ T cells on day 0 to the highest number for each animal revealed that changes in this cell population were not significant (p = 0.212, Mann–Whitney U test) (Fig. 2B).
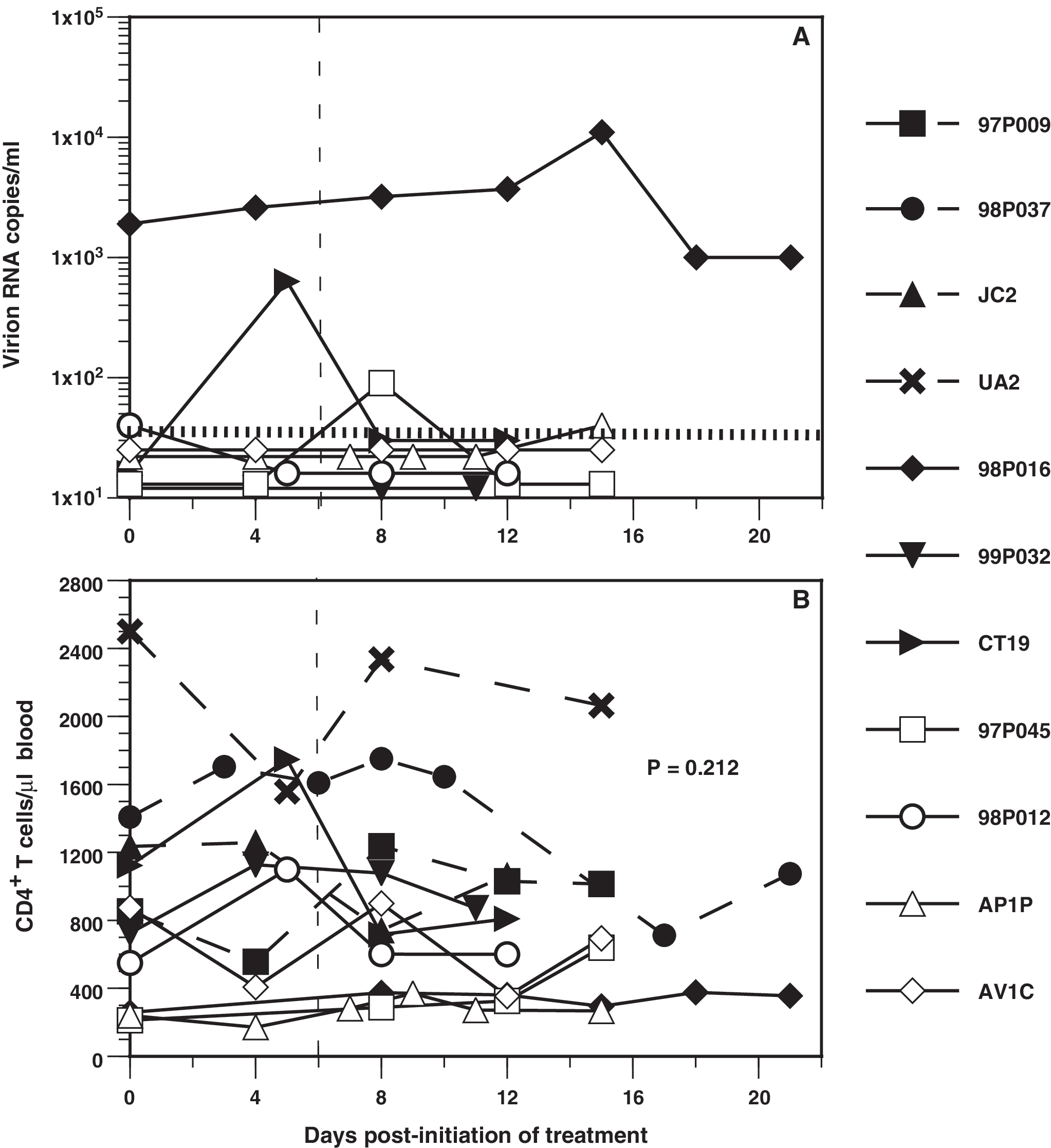
Changes in plasma virion RNA (
Changes in circulating numbers of mDCs and pDCs associated with FLT3-L
CD34+ myeloid and lymphoid progenitor cells express the FLT3 receptor; therefore, FLT3-L exogenously administered should promote proliferation and differentiation of DC subsets (Fig. 3A). Before injection of FLT3-L, the mean numbers of blood mDCs were lower in SIVmac239- and SHIV-89.6P-infected (50,200 and 67,800 mDCs/ml, respectively) macaques compared to those in naive animals (116,000 mDCs/ml) (Fig. 3B). However, no significant difference (Mann–Whitney U test) between naive and infected animals was noted, probably due to the broad distribution of mDCs and small sample sizes of each group. In general, the numbers of mDCs began to increase by day 4 or 5 and peaked around day 8; relative increases for individual animals ranged from 1.5- to 12-fold, with no differences between naive and infected animals (Fig. 3B). The relative increases in mDCs in SIVmac239-infected were higher than those in either SHIV-89.6P-infected or naive animals, resulting in slightly greater numbers of circulating mDCs when the numbers peaked.
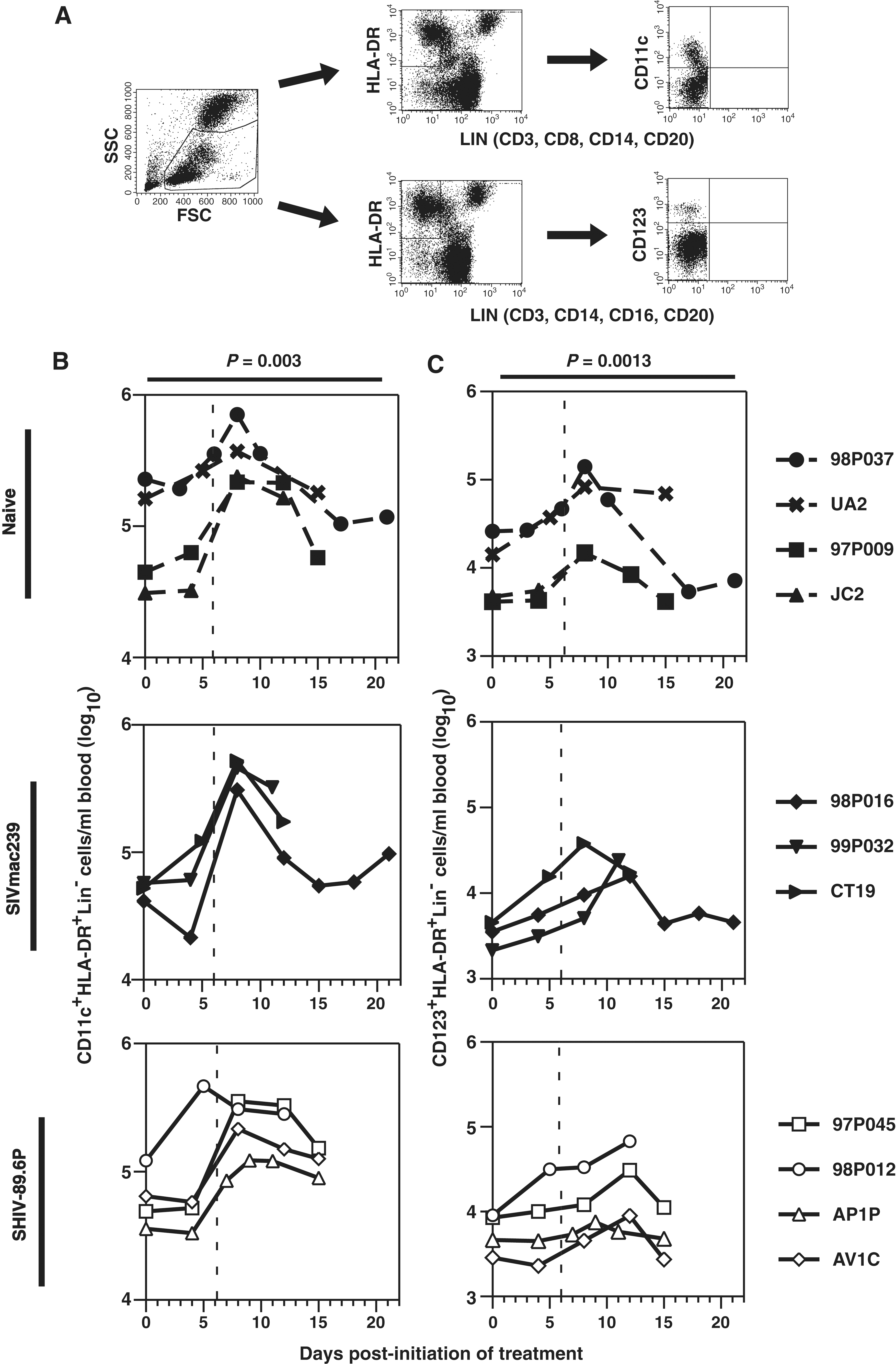
Influence of FLT3-L on DC subsets in macaques. (
Consistent with previous observations, on day 0 the mean numbers of blood pDCs were lower in animals infected with SIVmac239 (3405 pDCs/ml) and, to a lesser extent, SHIV-89.6P (6,242 pDCs/ml), compared to naive macaques (12,220 pDCs/ml) (Fig. 3C). 32,33 Again, these differences were not statistically different (Mann–Whitney U test). After FLT3-L treatment, pDCs increased as much as 10-fold in all groups, but the total numbers remained lower in infected compared to naive animals. Furthermore, the peak numbers of pDCs in both infected cohorts were greater than the normal range of 4000 to 17,000 pDCs/ml of blood previously reported for SIV-naive pigtailed macaques. 33 Although the numbers of pDCs in all naive macaques peaked on day 8, in five of seven infected animals, the peak occurred on day 12. Numbers of circulating mDCs and pDCs did not correlate with serum levels of FLT3-L (data not shown).
Immune activation following administration of FLT3-L
Initially, expression of activation markers on blood DCs during or after administration of FLT3-L was not examined. Later, cryopreserved PBMCs, obtained either before treatment or on either day 8 or 12 (depending on availability), were used to compare expression of CD40, CD80, CD86, and HLA-DR. No significant changes in expression (MFI) of CD40, CD80, or CD86 were observed on either mDCs or pDCs, but in most animals, CD40 on mDCs was elevated moderately and changes in MFIs approached significance (Fig. 4). In contrast, after FLT3-L treatment HLA-DR expression on both DC subsets was upregulated significantly, with MFIs increasing as much as 3-fold.
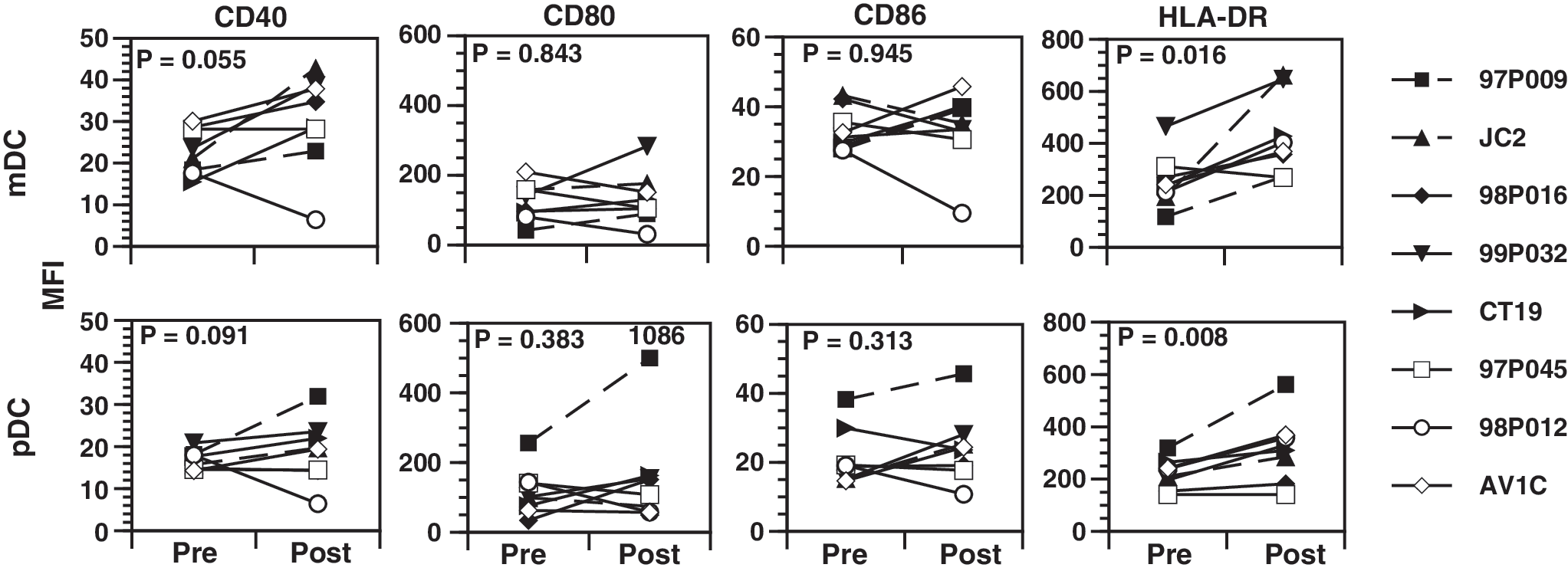
Expression of activation markers on FLT3-L-mobilized DCs. Using four-color flow cytometry and cryopreserved PBMCs, cell-surface expression (MFIs) of the activation/costimulatory molecules CD40, CD80, and CD86 was measured on CD11c+HLA-DR+Lin− mDCs and CD123+HLA-DR+Lin− pDCs. Cells collected before FLT3-L administration (Pre) were compared to those obtained on either day 8 (JC2, 97P009, CT19, 98P012) or day 12 (99P032, AV1C, 98P016, 97P045), depending on availability; PBMCs were not available for three animals; Wilcoxon signed rank test.
Monocytes and mDCs produce IL-12 and also secrete TNF-α; these cytokines synergize with pDC-secreted IFN-α to induce NK cell activation and promote a strong Th1 response. 58 –61 However, during HIV infection, secretion of IL-12 and IFN-α is impaired. Exogenous FLT3-L resulted in transient increases in absolute numbers of circulating monocytes, mDCs, and pDCs, and also was associated with higher cell surface expression of HLA-DR; therefore, we determined whether these changes coincided with increased concentrations of DC- and monocyte-specific cytokines. On day 0, IL-12 was detected in serum from all animals, but the mean values were more than 2-fold higher in naive compared to both SIVmac239- and SHIV-89.6P-infected animals (Fig. 5A). Moderate increases in serum IL-12 occurred between days 8 to 12, with the largest increase (>4-fold) in macaque 97P045. Before the first FLT3-L injection, only two macaques had detectable serum IFN-α (Table 2). In macaques 97P009, CT19, AP1P, and AV1C, IFN-α was measured as early as day 4, but generally after day 10. On day 0, all serum samples were negative for TNF-α, but in one naive and three of four SHIV-89.6P-infected macaques, TNF-α was detected on one or more occasions during the 21 days of FLT3-L treatment and observation. Concentrations of TNF-α increased to greater than 27 pg/ml in 97P045 and 97P009, which was unusual because serum TNF-α typically is undetectable in naive pigtailed macaques and in those infected with various SIV/SHIV strains. 62

Immune system activation in association with FLT3-L administration to macaques. (
IFN-α/TNF-α (pg/ml) were measured in aliquots of serum obtained on days when blood was drawn from each animal (see Table 1). A minus sign (−) indicates undetectable. Only animals with at least one detectable serum cytokine are listed; all serum samples from macaques 98P016, 99P032, and JC2 were negative for both cytokines.
Animal had been euthanized.
Both mDCs and pDCs activate T and NK cells, either by direct interactions or indirectly through cytokines, such as IFN-α, TNF-α, and IL-12, suggesting that the larger numbers of circulating DCs and higher serum cytokine concentrations might be associated with generalized activation and proliferation of both T and NK cells. 9,58 –60 However, no consistent changes were observed in the numbers of CD3+CD4+ T cells and any increases were probably influenced by fluctuations in the absolute numbers of total lymphocytes that occurred in response to FLT3-L (Fig. 2B). Also, there were no significant changes in the total numbers of CD3+CD8+ T cells and NK cells (defined as CD16+CD8+CD3− or NKG2A+CD8+CD3−) in either the infected or naive animals (data not shown). As a measure of T cell activation, we determined the frequencies of cells expressing the IL-2 receptor (CD25), which can be used either as a marker of T cell activation and proliferation or to identify Tregs. 63 –66 In most animals, increases in the frequencies of CD4+ and CD8+ T cells expressing CD25 were observed: mean days of maximum expression on CD4+ and CD8+ T cells were 11.4 and 7.9 days, respectively (Fig. 5B). The frequencies of NK and CD8+ T cells expressing another activation marker, CD69, also significantly increased; the mean times of maximum CD69 expression on CD8+ T cells were 10.4 days and on NK cells were 11.2 days. The frequencies of CD69+ NK cells were always greater than those of CD8+CD69+ T cells, indicative of a more activated cell phenotype and consistent with previous observations for pigtailed macaques (Q. Wei and P.N. Fultz, unpublished data). 62 Increases in both serum cytokine concentrations and activation states of T and NK cells suggested that FLT3-L administration induced peripheral immune system activation that coincided with maturation and increased numbers of circulating mDCs and pDCs.
Effects of FLT3-L on DCs in BM and secondary lymphoid tissues
To ascertain the effects of exogenous FLT3-L on DCs in macaque BM and peripheral LNs (PLNs), multiple single-cell suspensions of these tissues were evaluated. Before the study, the range of mDCs in BM was broad, but there was no apparent difference between numbers of mDCs in naive and infected animals, possibly related to the limited sample size (Fig. 6A). Similarly, the range of pDCs was highly variable, with some indication that pDCs in untreated animals might be lower in infected compared to naive macaques. In contrast to changes observed in blood, in the three animals for which day 4 biopsies were performed, the absolute numbers of both mDCs and pDCs in BM increased 8.5- and 2.5-fold, respectively. In most BM biopsies done after day 4, the numbers of mDCs and pDCs were consistently higher than those in the biopsies done pre-FLT3-L; e.g., in 98P016 mDCs and pDCs increased approximately 45- and 7-fold, respectively.
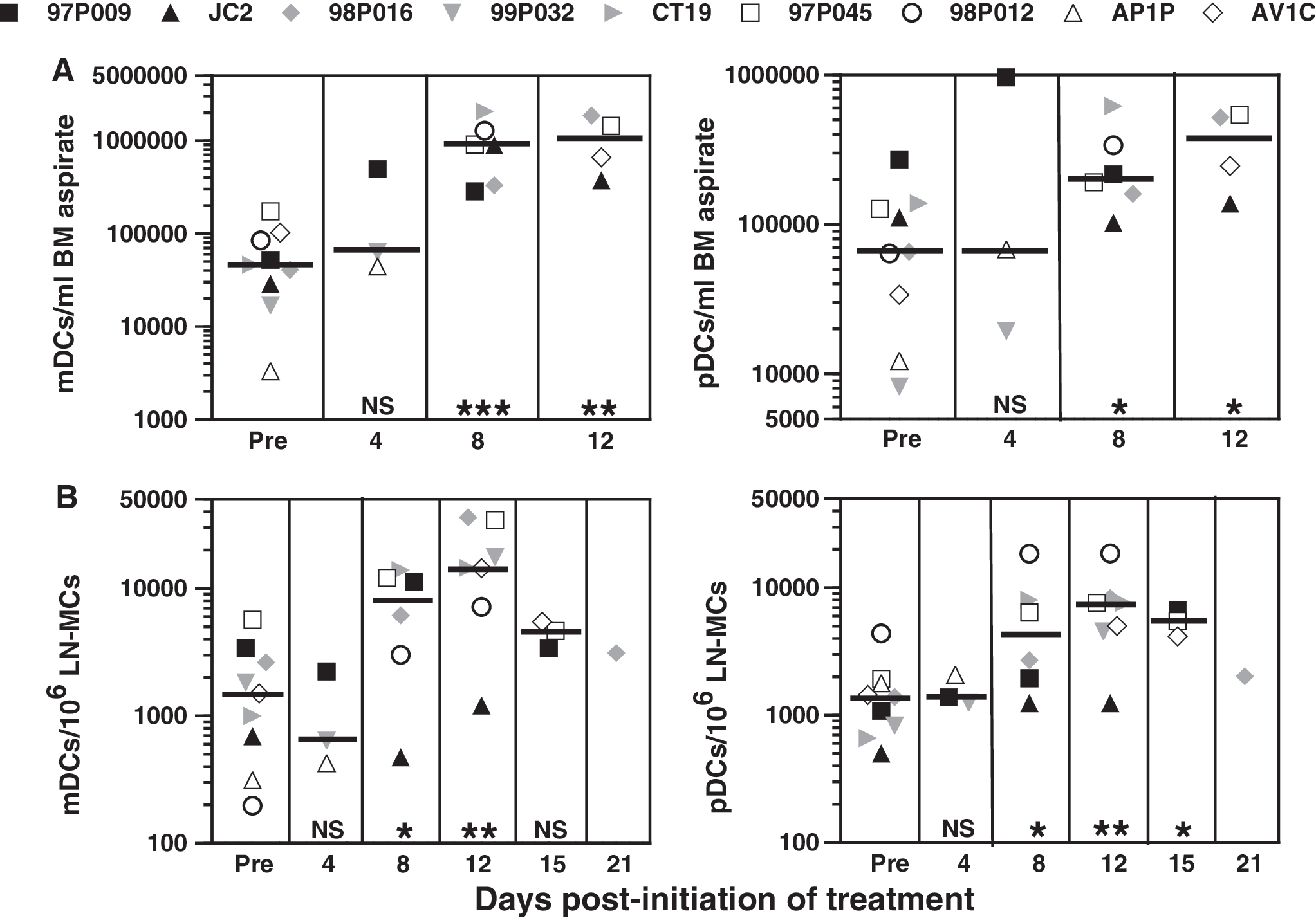
Augmentation of DC subsets in BM (
In these cohorts of naive and infected animals there were no differences in the initial numbers of either DC subset in PLNs (Fig. 6B). In the three macaques for which PLN biopsies were performed on day 4, no changes in either mDCs or pDCs were observed; however, mDCs and pDCs in most animals increased during days 8 to 12 and then stabilized or decreased. The largest increase in mDCs was 35-fold for macaque 98P012 and in pDCs, 11-fold for macaque CT19.
mDCs and pDCs were enumerated in mesenteric LNs (MLNs), spleens, lungs, and jejunums collected at necropsy (Table 3). While the numbers of DCs in each of these tissues varied considerably, mDCs typically were more prevalent than pDCs, consistent with observations in blood, BM, and PLNs. Lung and spleen tissues were particularly enriched for mDCs, while jejunums appeared to contain the lowest numbers (Fig. 7). Since MC fractions from jejunums were eluted over glass wool columns to remove epithelial cells and mDCs exhibit some adherent properties (R.K. Reeves, unpublished), the prevalence of these cells in jejunums might be underestimated. Furthermore, although DCs were not enumerated in MLNs, spleens, lungs, and jejunums of FLT3-L-naive animals, we previously reported pDC frequencies in MLNs and spleens of SIV/SHIV-infected and uninfected macaques that did not receive FLT3-L. 33 The median numbers of pDCs/106 MLN-MCs were 300, 600, and 400 in SIVmac239- and SHIV-89.6P-infected and uninfected animals, respectively. The numbers of pDCs in MLNs of FLT3-L-treated macaques in the current study were consistently greater (up to 15-fold, Table 3), regardless of viral status. Interestingly, the numbers of pDCs in spleens of FLT3-L-treated virus-naive and SHIV-89.6P-infected macaques were comparable or only slightly greater than the numbers of pDCs in the corresponding FLT3-L-naive macaques previously studied. 33 All FLT3-L-treated SIVmac239-infected animals had greater numbers than the median of approximately 600 pDCs in untreated, similarly infected macaques; e.g., 99PO32 had almost 5000 pDCs/106 splenic MCs. These results suggested that expansion of DCs in tissues other than blood, BM, and PLNs also occurs in response to high levels of FLT3-L; however, a caveat to the tissue data (Table 3) is that MLNs, spleens, lungs, and jejunums from FLT3-L-treated macaques were taken on day 12 or later, which could be after the time of peak DC expansion.

Detection of mDCs (
mDCs and pDCs (gated using the same strategies as for blood; see Fig. 3A) are expressed as the number of DCs per 106 mononuclear cells (MCs) isolated from the indicated tissues.
MLN, mesenteric lymph node; Jeju, jejunum; NA, not analyzed.
Days at which each animal was euthanized and necropsied after the first dose of FLT3-L was given.
Discussion
Strategies to treat HIV disease and its associated hematologic abnormalities are often tested using macaque models. In this study we inoculated uninfected and SIV- or SHIV-infected pigtailed macaques with the hematopoietic cytokine FLT3-L and evaluated its effects on (1) hematologic parameters, (2) SIVmac239 and SHIV-89.6P viremia and disease progression, and (3) immune cell activation. Previous studies in which humans and nonhuman primates were inoculated with FLT3-L did not quantify serum concentrations and most did not evaluate animals infected with a lentivirus. Normal serum concentrations of FLT3-L in pigtailed macaques were similar to levels found in two studies of healthy humans, which reported medians of 69 and 72 pg of FLT3-L/ml, 38,40 but they were lower than a mean of 132 pg of FLT3-L/ml reported for a cohort of 72 cynomolgus macaques (M. fascicularis). 67 In our study, serum FLT3-L increased to more than 1000 pg/ml in some animals, but quickly returned to baseline levels, suggesting rapid receptor-mediated uptake and/or metabolism of the exogenous cytokine. Our findings, however, indicate that peak levels of FLT3-L in macaques were similar to those observed in humans and macaques with chemotherapeutic- or radiation-induced aplasia where hematopoiesis is upregulated due to pancytopenia. 41,44,67
Higher serum concentrations of FLT3-L coincided with higher numbers of monocytes, but alterations in the numbers of circulating lymphocytes were inconsistent. This finding agrees with that of Papayannopoulou et al. 68 who showed that in a cohort of FLT3-L-treated naive pigtailed macaques, myeloid progenitors were mobilized at a greater frequency than lymphoid progenitor cells. Coates et al., 47 however, reported a significantly higher frequency of lymphocytes in rhesus macaques given FLT3-L and, although the absolute numbers of circulating lymphocytes were not shown, this disparity might reflect minor differences in the responsiveness to FLT3-L of pigtailed compared to rhesus macaques. We observed a transient decrease in platelets, a result not unexpected since it was reported that when added to cell cultures, FLT3-L inhibited megakaryocyte differentiation from CD34+ progenitors. 69,70 Furthermore, increased serum levels of FLT3-L in cynomolgus macaques that developed radiation-induced aplasia were associated with a decline in platelets, and FLT3-L treatment counteracted the effects of thrombopoietin-induced restoration of platelet counts in myelosuppressed rhesus macaques. 67,71 Although the decrease in platelets that we observed was minimal and resolved within about 4 days after the last FLT3-L injection, it is possible that therapeutic administration of FLT3-L during advanced lentiviral disease, when thrombocytopenia is common, might elicit complications.
FLT3-L is a requisite cytokine for development of progenitor cells of lymphoid and myeloid lineages in BM and, also, for expanding mDCs and pDCs derived from these progenitor cells in peripheral lymphoid tissues. 36,43 –46,72,73 We report here expansion of mDCs and pDCs not only in naive but also in infected macaques that have reduced DC numbers, similar to HIV-1-infected patients. 25,28,31 –33 Compared to previous studies in which mDCs in SIV-naive rhesus macaques were amplified more than 50-fold, 47,48 we observed variable increases of up to 45-fold in mDCs. Since the magnitude of expansion of mDCs in naive and infected macaques did not differ, the generally higher fold increases in other studies cannot be attributed to infection status, but might reflect either differences in rhesus versus pigtailed macaques or natural variation among outbred animals. Also, it is possible that our evaluations were not done at optimum times since Teleshova et al. 48 found the peak of DC expansion to be 10 days after initiation of the 7-day regimen, a day on which we did not examine most animals. While the magnitude and kinetics of monocyte and mDC expansion did not differ significantly between naive and infected macaques, the peak numbers of pDCs generally occurred later in infected (day 12) than in naive animals (day 8). The initial numbers of pDCs in blood from SIVmac239- and SHIV-89.6P-infected, compared to naive, animals were lower, suggesting that immediate pDC precursors might also be lower, which might delay increases of pDCs in blood. Although viral replication was generally not detected in these animals, it is possible newly activated, more mature pDCs, which are susceptible to lentivirus infection, 33 might have become infected and destroyed, thus obscuring migration of these cells in blood. It should be noted, however, that tissue pDCs have short half-lives and must be replenished continually. 74 Thus, maintenance of higher numbers of pDCs in LNs during HIV/SIV infections implies continual migration of these cells from BM to blood to LNs, which would be facilitated by persistent inflammatory conditions.
Compared to uninfected persons, activation marker expression on DCs is generally higher during chronic HIV-1 infections, 31 but we found no such distinction in CD40, CD80, and CD86 expression when comparing cells from untreated naive and infected macaques. These differences might be related to the aviremic status of most of the macaques, a condition that might be associated with minimal immune system activation, or simply to differences in virus–host interactions in humans versus macaques. Also it is possible that the limited number of animals in our cohorts did not exhibit the full range of expression of these cell surface proteins. FLT3-L treatment of macaques, however, was associated with increases in HLA-DR expression on both mDCs and pDCs; CD80, CD86, and CD40 appeared to be upregulated in some animals. Using the same FLT3-L regimen, Teleshova et al. 48 reported significant upregulation of CD80 and CD86 on mDCs and pDCs in rhesus macaques. In FLT3-L-treated melanoma and renal cancer patients, CD86 and HLA-DR, but not CD80, were upregulated on DCs. 75 DCs mobilized in the lungs of FLT3-L-treated mice expressed higher levels of CD40, CD86, and HLA-DR than DCs in placebo-treated controls. 76 Combined with our data, these observations strongly indicate that, in general, blood DCs not only increase in number during FLT3-L treatment, but also can be activated.
Coinciding with increases in circulating mDCs and pDCs were transiently higher serum concentrations of IL-12 and, in some macaques, of IFN-α and TNF-α. Consistent with our results, FLT3-L treatment of melanoma patients elicited higher serum levels of TNF-α (up to 100 pg/ml), 77 and isolated DCs from both macaques and mice produced higher amounts of IFN-α and IL-12 ex vivo. 78 Similarly, in vivo administration of FLT3-L resulted in higher amounts of IL-12 that accumulated in medium when macaque PBMCs were cultured ex vivo and compared to cultures from untreated animals. 48 The results with IFN-α were in contrast to those regarding IL-12. In both SIV-infected macaques and HIV-infected persons, pDCs were activated and produced large amounts of IFN-α in vivo, but these cells were refractory to stimulation in vitro and, consequently, produced lower amounts of this cytokine when cultured. 79,80 While the overall conclusion is that administration of FLT3-L to macaques enhances cytokine production, the identity of specific cell types and the tissue compartments in which they reside is unclear.
FLT3-L treatment of macaques induced higher frequencies of circulating CD25+CD4+ T cells, which are likely to be activated CD4+ T cells, with the possibility that some might be Tregs. Although evaluation of a more definitive Treg marker, such as FoxP3, would confirm the latter idea, mDCs and pDCs are known to induce Treg development by mechanisms that can involve IFN-α, TNF-α, and indoleamine 2,3-dioxygenase. 11,81 –85 Specifically, during acute SIV infection of cynomolgus macaques, Malleret et al. 85 found that pDCs and type I IFNs played important roles in eliciting an immunosuppressive state. Furthermore, IFN-α/β produced by pDCs following HIV infection can modulate proliferation and depletion of effector CD4+ T cells, also contributing to immunosuppression. Since Tregs can influence HIV/SIV disease progression by limiting virus-specific immune responses, 65,66,85 –88 an increase in this population potentially might be detrimental for FLT3-L-based immunotherapies for HIV. However, Tregs, in general, are only a minor proportion of any CD4+CD25+ population. 89 Our data also indicated that FLT3-L administration upregulated activation states, but not numbers, of NK cells, which contrasts with some murine studies in which FLT3-L treatment led to significant increases in NK cells concurrent with DC mobilization. 90 The increased CD69 expression on NK cells that we observed likely is linked to higher serum IL-12, which activates NK cells. 91 Whether activation enhanced the lytic activity of NK cells, which would be beneficial in lentivirus infections where NK cell numbers and functions can be impaired, 92 –94 was not evaluated.
FLT3-L promotes cellular activation and proliferation; therefore, a major concern when treating lentivirus-infected individuals with this cytokine is the possibility of inducing virus replication. Transient increases in viremia were detected only in three animals, similar to Bucur et al. 95 who found no increases in viral burdens in a group of SIVmac239-infected rhesus macaques treated with FLT3-L. Also, Koopman et al. 49 observed no changes in viremia in a cohort of SIV-infected macaques treated with the related FLT3 agonist, ProGP-1. However, in our study, vRNA in plasma from 98P016 increased almost 6-fold after 15 days, suggesting that FLT3-L potentially might augment viral replication in animals with measurable viremia. Studies using larger numbers of macaques with higher levels of viremia at the start of the study are needed to address this possibility.
With the exception of Thomson's group, who also evaluated LNs, livers, and kidneys in rhesus macaques, 47,96 previous studies showing FLT3-L-induced mobilization of DCs in humans and macaques evaluated only blood. Other studies utilized rhesus or cynomolgus macaques whereas we used pigtailed macaques—three distinct species—that are known to differ physiologically and in their pathological responses to some SIV strains. FLT3-L is expressed ubiquitously in tissues, while the FLT3 receptor is limited to CD34+ progenitor cells, including those for DCs 36,37 ; therefore, initial DC activation should occur where CD34+ cells are concentrated. As in humans, we verified this site to be macaque BM and both mDC and pDC activation and mobilization in BM appeared to precede that in blood. 35 In support of this finding, there was a significant correlation between serum levels of FLT3-L and DC numbers in BM (mDCs, R = 0.5630, p = 0.006; pDCs, R = 0.5991, p = 0.003), but not in blood. Although BM damage occurs in HIV, SIV, and SHIV infections, 97 –99 it was not obvious that DC activation in BM was impaired in infected animals. Our detection of significant increases in both mDCs and pDCs in the PLNs of all animals is similar to that in previous surveys of macaques. It is interesting that, as in blood, DC numbers did not increase in PLNs on day 4, which is highly suggestive that DCs originated in the BM and then migrated through the blood to LNs. Because we detected low frequencies of CD34+ cells in both blood and LNs (not shown), we cannot exclude the possibility that some newly generated DCs were derived from localized progenitor cells and did not originate directly from BM precursors. Consistent with this possibility, Waskow et al. 73 recently showed that DC precursors transit from BM to spleen and LNs where their division is controlled by FLT3-L.
Thus, based on our preliminary results in macaques chronically infected with SIV or SHIV and those of others showing increased numbers of different leukocyte subsets in blood and tissues following administration of FLT3-L to macaques, 47 –49 additional studies are required to establish the potential efficacy of FLT3-L therapy in HIV-infected persons. These future studies should be a comprehensive evaluation of the positive and negative effects of FLT3-L administration to SIV-infected macaques that are nonprogressors with low viral loads and to those exhibiting signs of disease progression.
Footnotes
Acknowledgments
The authors thank M.L. Spell and the UAB Center for AIDS Research Flow Cytometry core, supported by NIH Grant P30 AI027767, for flow cytometric analyses; Dr. Robert Baker and Deidra Isbell for performing biopsies; and Drs. Mike Piatak and Jeff Lifson for determining plasma virion RNA concentrations. Amgen Corp. (Thousand Oaks, CA) generously provided FLT3-L. This work was supported by grants from the National Institutes of Health: UO1 AI028147; three animals were supported by NIAID RO1 AI049784, as part of a collaborative study with Dr. James Hoxie (to be reported elsewhere), who provided the SIVmac239 mutant viruses for the original study. R.K.R. was supported by an NIH Basic Mechanisms of Virology training grant, T32 AI007150-29.
Disclosure Statement
No competing financial interests exist.