
Abstract
Characterization of HIV-1 from slow progressors is important to facilitate vaccine and antiviral drug development. To identify virus attenuations that may contribute to slower rates of disease progression, the full-length viral genomes from primary isolates of six slow progressing HIV-positive children were sequenced. Proviral DNA was extracted from cocultured peripheral blood mononuclear cells and used to PCR amplify, sequence, and extensively analyze the near full-length genomes and LTR regions. All primary HIV-1 isolates were HIV-1 subtype C throughout their genome, and amino acid (AA) sequence analysis revealed open reading frames for all genes. However, all isolates had at least one unusual gene/protein. For example, isolate LT5 had a 2AA insertion in the Vpr mitochondriotoxic domain. Isolate LT21 contained an additional 5AA in the C-terminus of tat exon 2, while integrase in isolate LT39 had an additional 4AA at the C-terminus. Rev from isolates LT45 and LT46 did not have the characteristic subtype C 16AA truncation, and in addition, had a further 3AA. Furthermore, altered functional domains were noted in several isolates, such as the cAMP-dependent kinase PKA phosphorylation site in Nef (LT5), a Vpr mutation involved in decreased proapoptotic activity (all isolates), and the Nef ExxxLL motif involved in the interaction with AP-1 and AP-2 (LT46). The slower HIV-1 disease progression in these six children may be attributed to altered protein functions. For example, LT46 Nef is unable to bind AP-1 and AP-2 and therefore is inactive on CD4 endocytosis. The biological relevance of these findings requires further investigation.
Introduction
Rates of disease progression to AIDS among HIV-1-infected patients are highly variable and depend on a complex interplay between the host's genetic factors, immunological responses (reviewed by Lama and Planelles
1
), and the pathogenic potential of the virus.
2
The natural history of HIV-1 infection and disease progression has been well established in adults, and the median time from infection to development of AIDS is approximately 10 years for intermediate progressors
3
or 3 to 6 months for rapid progressors.
4
Interestingly, some patients remain clinically asymptomatic (5% of infected patients) after 7–10 years postinfection in the absence of antiretroviral (ARV) therapy, and are termed slow progressors, SP, or long-term nonprogressors, LTNP.
5
A subset of infected individuals (elite controllers) with spontaneous and sustained control of HIV-1 infection, with viral loads <50 RNA copies/ml, in the absence of therapy has been described.
6
This spontaneous control occurs in approximately 1 in 300 HIV-infected persons, and the HIV Controller Consortium was recently established to study host genetics together with functional immunology studies in elite controllers (
Vertically infected children, however, have shorter clinical latent periods and survival times than HIV-1-infected adults. 7 In the developed world approximately 20% of children progress rapidly to AIDS within the first 2 years after birth, 7 whereas in sub-Saharan Africa up to 50% of children are rapid progressors. 8 Asymptomatic vertically infected children older than 7 years of age are unusual, 9 and in a recent prospective study of the ANRS French Pediatric Cohort, only 2% of children infected showed no clinical progression at 10 years. 10
SP, LTNP, or elite controllers are of great medical interest because they provide insights relevant to understanding HIV-1 pathogenesis and biological and molecular correlates of protection against HIV-1 infection and disease progression, viral attenuation, novel therapies, and vaccine design. 6 Various host factors have been described that modulate HIV-1 disease progression to AIDS. Nonprogressors are more likely to develop vigorous HIV-specific CD4+ T helper (Th1) cell responses, 11 strong neutralizing antibodies, 12 and strong and persistent cytotoxic T-lymphocyte responses. 13 Similarly, disease progression is influenced by diverse host genetic factors such as the genotype and extent of heterozygosity at major histocompatibility complex class I loci, 14,15 polymorphisms in CCR5, 16 SDF-1, or CXCL12 17 and APOBEC3G (apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3G), 18 and polymorphisms in and near the ring finger protein 39 (RNF39) and zinc ribbon domain-containing 1 (ZNRD1) genes. 15
Several viral factors that may impact disease progression have also been described. The subtype of the infecting virus may influence the rate of HIV-1 disease progression to AIDS: in Senegal, patients infected with non-A subtypes, such as C, D, and G, were shown to be eight times more likely to progress to AIDS than those infected with subtype A. 19 Similarly, in Uganda several studies have shown that subtype D is associated with faster disease progression compared to subtype A. 20,21 Infection with, and persistence of attenuated HIV viruses and/or mutations in functional sites of viral genes and proteins have been correlated with low replicative capacity and the lack of disease progression. The most studied defects associated with the absence of disease progression have been found in the nef/LTR region as documented in an asymptomatic patient from central Massachusetts, 22 eight patients in the Sydney Blood Bank Cohort (SBBC, sequenced the full genomes), 5,23,24 and a slow progressing Italian individual. 25 Subsequently, numerous researchers have described “defective” nef alleles that impact disease progression in various paediatric and adult cohorts. 25 –32 Several researchers have found LTNP with unusual or defective genes other than nef/LTR, including gag, env, vpu, vpr, vif, rev, and tat genes. 33 –44 However, since only selected genes were sequenced, we cannot exclude the possibility that other defective genes may contribute to viral attenuation in these studies. Full genome sequencing of HIV from individuals with slow or nonprogressing disease established an association between structural or functional alterations in gp41, Vpu, Gag, and Nef, 45 and Vif 37 and disease progression.
Available evidence suggests that disease progression is a complex process, and justifies the need for studying cohorts of patients infected with different HIV subtypes. South Africa has one of the fastest growing HIV-1 epidemics, with over 5.2 million infected people (
Materials and Methods
Patient samples and primary virus isolates
Primary viral isolates obtained from six ARV drug-naive vertically infected children attending a clinic at Chris Hani Baragwaneth Hospital, Johannesburg, were selected for the purposes of this pilot study. The children fit the following definition of SPs: documented infection lasting at least 6 years and asymptomatic or mild/moderate symptoms in the absence of ARV drug use. The patient's epidemiological data are shown in Table 1. Ethical approval was obtained at the time of sample collection (M971010) from the University of the Witwatersrand Committee for Research on Human Subjects.
Epidemiological and Clinical Data of HIV-1 Vertically Infected Children a
Data from children attending the Chris Hani Baragwaneth Hospital in Johannesburg, 72 identified as potential slow progressors, and the biological and genotypic data of their primary viral isolates.
At time sample was taken.
R5/NSI: CCR5-utilizing, non-syncytium-inducing.
Antiretroviral drug naive.
V82A in protease reduces susceptibility to ritonavir, indinavir, nelfinavir, and lopinavir in vitro.
Infected peripheral blood mononuclear cells (PBMCs) from the six primary HIV-1 isolates (passage one) were expanded by coculturing with 2 × 106 cells/ml phytohemagglutinin (PHA)-activated seronegative donor PBMCs, according to standard PBMC coculture techniques. 46 Viral growth was monitored using the Murex p24 enzyme-linked immunosorbent assay (ELISA) kit (Abbott Murex, Dartford, UK). The expanded passage (P2) cultures were centrifuged at 1400 rpm, and the infected PBMCs and supernatants were stored at −130°C and −70°C, respectively. Proviral DNA was extracted from p24 antigen-positive cultures from P2 PBMCs using the High Pure Template Preparation Kit (Roche), as per the manufacturer's instructions. The virus containing supernatant from P2 was used to establish primary virus phenotypes and biotypes by infection of MT-2 and CCR5 and CXCR4-expressing U87.CD4 cell lines, respectively, as described previously. 46 Viral supernatant was used to extract viral RNA (High Pure RNA Isolation kit; Roche), as required.
Polymerase chain reaction and sequencing
A nested polymerase chain reaction (PCR) was carried out to amplify the virtually full-length genomes using the outer primers MSF12 and OMFR1 and inner primers F2NST and OFM19 (when necessary), and cycle conditions as described previously, 38 using the Expand Long Template Taq Polymerase kit (Roche) in the GeneAmp PCR systems 9700 thermal cycler (Applied Biosystems, Foster City, CA). A separate LTR amplicon was generated using the outer primers JL17 and LTR3, inner primers VWLG-for and VWLG-rev, and cycle conditions as described previously, 38 using Amplitaq Gold (Applied Biosystems) in the GeneAmp PCR systems 9700 thermal cycler (Applied Biosystems). Pooled virtually full-length genome PCR products were purified on a Microcon-YM50 column (Millipore, Bedford, MA), and LTR PCR products were purified using a High Pure PCR Product Purification Kit (Roche) as per the manufacturer's instructions. Primers spanning the entire HIV-1 genome 47 were used to sequence the PCR products in both directions. Amplicons were sequenced using the ABI Big Dye terminator system (version 3.1) according to the manufacturer's instructions, and run on the ABI Prism 3100 Genetic Analyzer (Applied Biosystems).
Genome and phylogenetic analysis
All generated sequence data were edited using the Sequencing analysis 3.3 program (Applied Biosystems), and the near full-length and LTR sequences were assembled using Sequencher 4.5 (Genecodes, Ann Arbor, MI) to yield the complete full-length genome sequence in one contig. A multiple alignment of the near full-length genomes with reference sequences from HIV-1 subtypes A to K and CRF01_AE and CRF02_AG (
Results
The HIV-1 primary virus isolates from six vertically infected children were successfully expanded using PHA-stimulated donor PBMCs. All six isolates were found to utilize the CCR5 coreceptor for cell entry, and exhibited the non-syncytium-inducing (NSI) phenotype (Table 1). Proviral DNA was successfully extracted and used to PCR amplify and successfully sequence the near full-length genomes and LTR fragments. Extensive sequence analysis revealed that all six full-length genomes encoded for the correct open reading frames (ORFs), and ranged from 9703 to 9767 bp in length (Table 1). Nucleotide sequences for the full-length genomes were submitted to GenBank and are available under the accession numbers listed in Table 1. Phylogenetic analysis for the near full-length genomes and the LTR regions showed that all six isolates clustered within subtype C with a bootstrap value of 100% (Fig. 1B). In addition, all six isolates were subtype C throughout their genomes by similarity plots (results not shown).
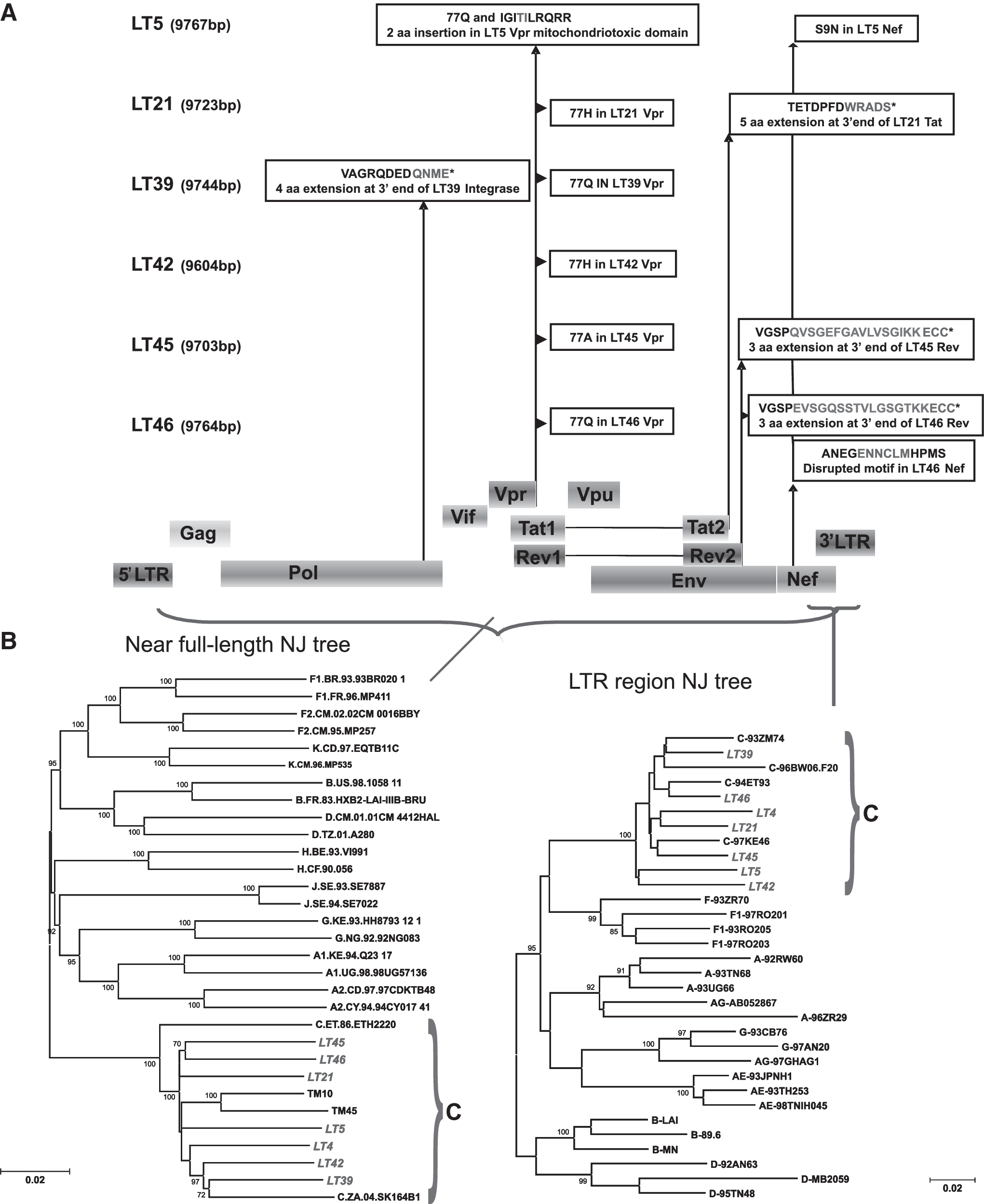
(
The nucleotide sequences and translated amino acids for all the HIV-1 proteins of all six isolates were extensively analyzed and compared to each other to identify aberrant genes/proteins. All isolates were found to have unusual genes/proteins (listed in Table 1). A schematic representation of the translated amino acid sequences for the unusual integrase, vpr, tat, rev, and nef genes is shown in Fig. 1A. Isolate LT39 had a 4AA sequence extension at the C-terminus of integrase (IN). Isolate LT5 has a 2AA insertion in the mitochondriotoxic domain of Vpr (viral protein R). In addition, isolates LT5, LT39, and LT46 contain 77Q, isolates LT21 and LT42 contain 77H, and isolate LT45 has the 77A polymorphism in Vpr. Isolate LT21 contained an additional 5AA in the C-terminus of Tat exon 2. Rev in isolates LT45 and LT46 had a 3AA extension. Isolate LT5 contains the Nef S9N mutation and the Nef ExxxLM motif is disrupted in LT46. There were no obvious “defects” in the LTR regions and the remainder of the coding regions of the genome for all isolates (results not shown).
Analysis of the polymerase gene nucleotide sequences for the presence of ARV drug resistance mutations (Stanford database,
The V3 loop region of the envelope glycoprotein (gp120) is involved in coreceptor usage, thus the V3 amino acid sequences were extensively analyzed and compared (Fig. 2). The length, overall positive charge, amino acids at positions 11 and 25, and tetrapeptide motif at the tip of the V3 loop are characteristic of CCR5 utilizing envelopes and are correctly correlated with the phenotype/biotype of each primary isolate (Table 1).

Alignment of the V3 amino acid sequences of the six newly characterized HIV-1 subtype C isolates. The sequences are compared to the HIV-1 subtype C consensus and ancestral sequence (
Discussion
In this pilot study we have shown that the six primary HIV-1 subtype C viruses isolated from potential SP or LTNP all harbored potentially defective genomes. Analysis of the polymerase gene nucleotide sequences for the presence of ARV drug resistance mutations revealed that no resistance mutations to RT or protease inhibitors were found, with the exception of V82A in the protease of LT46, which reduces susceptibility to ritonavir, indinavir, nelfinavir, and lopinavir in vitro.
48
The presence of V82A in untreated individuals has been reported (
Nucleotide and amino acid sequence analysis revealed all viruses had open reading frames for all genes and encoded for all the expected proteins. However, all the viral isolates were found to harbor at least one unusual gene/protein. Overall, all patients contain HIV-1 harboring Vpr with altered/decreased proapoptotic activity. Furthermore, HIV-1 from patient LT21 exhibited an unusual tat exon 2 variant that could alter tat-mediated reverse transcription. Patient LT39 is infected with HIV-1 containing IN with potentially altered efficiency for full site integration. Finally, patients LT45 and LT46 harbored virus with an ECC extension in Rev that may alter the Rev structure, impairing its ability to bind to RNA and other cellular factors or prevent ternary complex disassembly. In addition, the HIV-1 Nef from patient LT5 contains the S9N mutation, which alters the cAMP-dependent kinase PKA phosphorylation site, and LT46 contains a disrupted ExxxLM motif that may render the LT46 Nef unable to bind adapter protein 1 (AP-1), and therefore is inactive on CD4 endocytosis. It is possible that these alterations lead to altered viral replication, which impacts disease progression in these children.
The 4AA QNME extension in the IN of LT39 was unique to this isolate. IN mediates the integration of cDNA into the host chromosome, an essential step in HIV replication. Integration is mediated by a large nucleoprotein complex (PIC) containing viral DNA, Vpr, IN, and several cellular proteins such as the high mobility group protein A1 (HMGA1), 49 the integrase interactor protein (IN-1/SNF5), 50 and cellular lens epithelium-derived growth factor (LEDGF/p75). 51 The recognition sequence for the IN enzyme is located at the ends of the viral LTRs, 52 binding to which requires at least a cross-linked tetrameric form of IN to perform full-site integration. 53 Integration of proviral DNA occurs preferentially into transcriptional units of active genes, 54 while centromeres and telomeres are not favored targets. 55 IN catalyzes the integration of viral cDNA into the host chromosome via 3′ processing and strand transfer: the first is by processing the 3′ ends of linear viral cDNA in which the GT nucleotide pairs of the viral DNA are removed leaving a CA dinucleotide at each 3′ end (occurs in the cytoplasm), and the second is the strand transfer (or joining reaction) in which the processed viral DNA ends are inserted into host DNA (occurs in the nucleus). The C-terminal region of IN is involved in nonspecific DNA binding. It is also important for oligomerization of the enzyme and is necessary since IN monomers/dimers can integrate only one side of the LTR. It is possible that the tetrapeptide QNME insertion identified at the C-terminus of IN from isolate LT39 disrupts IN oligomerization, thereby altering the efficiency of the full site integration activity. However, recent results from our laboratory show this C-terminal tetrapeptide insertion is found among other HIV-1 subtype C IN sequences (unpublished data), warranting further studies for its role in the integration process.
Vpr is responsible for multiple functions of HIV-1, including nuclear import of the preintegration complex, apoptosis, fidelity of the RT process, G2 cell cycle arrest, and transactivation of the LTR and/or target genes thereby controlling HIV-1 gene expression. 56 The 96 amino acid Vpr protein is packaged into the HIV-1 virion and released into the cytoplasm during initial cell infection where it can exert its actions. The vpr gene exhibits a wide range of naturally occurring polymorphisms, 57 and the association between these and disease progression warrants further investigation. The Vpr R77Q mutation has been associated with long-term nonprogression and affects the mitochondriotoxic domain of the wild-type gene, which has been shown to induce T cell apoptosis, 58 and may be necessary for virulence. 59 Isolate LT5 has a unique 2AA insertion in the mitochondriotoxic domain of Vpr. Previous research showed that R77Q was associated with slower disease progression by decreasing the proapoptotic activity of Vpr. 58 Importantly, all isolates contained either 77Q, 77H, or 77A and are all likely to contain Vpr with altered/decreased proapoptotic activity. Interestingly, Vpr has neurotoxic effects, 60 and a defective vpr gene has recently been implicated in HIV-1 dementia. 61
Tat is mainly responsible for transactivation of HIV-1 gene expression in conjunction with the TAR element, 62 and other diverse Tat functions have been reviewed by Harrich et al. 63 Tat is encoded by two exons, and in natural infection both exon 1 and exon 2 are transcribed by cells, so different forms have different molecular functions. Tat exon 1 is highly conserved and its main function is to mediate transactivation of the viral LTR promoter. It can also induce monocyte chemotaxis and protein–protein interactions as well as uptake of soluble Tat by cells. Exon 2 conveys improved viral replication and has been implicated in chemoattraction of monocytes and dendritic cells, 64 in cell adhesion and binding to cell surface integrins, in modulation of MHC class I gene expression, in its ability to inhibit reverse transcription, Tat-mediated apoptosis, 65 and neurotoxicity, 66 and can bind human translation elongation factor 1 delta. 67 Isolate LT21 contained an additional 5AA in the C-terminus of Tat exon 2. There are no previous reports of a 5AA extension in the databases (rarely see 4AA). Tat exon 1 has been shown to facilitate the first strand transfer reaction of RT to cDNA, while Tat exon 2 (regions 86 to 101) inhibits cDNA synthesis when Tat concentrations reach a critical level. Future work needs to elucidate whether the LT21 tat exon 2 (86 to 106) variant is an enhancer or suppressor of tat-mediated reverse transcription.
Rev initiates the late phase of virus gene expression by interacting with the Rev responsive element (RRE) on the full length 9-kb and singly spliced 4-kb viral RNA transcripts, and their nuclear import and export are mediated by binding of the Rev nuclear localization signal to importin β, or the Rev nuclear export signal to Crm1/Exportin-1, respectively. Rev thus plays a critical role in the production of HIV-1 structural proteins. 68 Rev in isolates LT45 and LT46 had a 3AA extension. There are only two other reports of this Rev polymorphism: the SBBC LTNP viruses 39 and SP from vertically infected siblings (TM10 and TM45) from the same cohort in South Africa. 38 Rev activity is defined by domains or motifs for nuclear export of RNA, RNA binding, and nuclear import signals, which have been shown to be conserved during vertical transmission. 69 Thus the presence of the ECC extension may alter the Rev structure, impairing its ability to bind to RNA and other cellular factors or prevent ternary complex disassembly.
The functions of Nef have been reviewed by Das and Jameel. 70 Nef causes endocytosis and lysosomal degradation of CD4 and downregulation of MHC class I and II. The leucine residues in the Nef ExxxLL motif are required for CD4 downregulation and are involved in the interaction with AP-1. This motif is disrupted in LT46 (ExxxLM). This may render LT46 Nef unable to bind AP-1, and therefore inactive on CD4 endocytosis. Thus, no/low degradation of CD4 in an infected cell will most likely impact the production of infectious virions. In addition, the cAMP-dependent kinase PKA phosphorylation site in Nef of LT5 is altered. Nef aberrations have been well described 26 and frequently involve the LTR regions of the viral genome. The inability of the virus to cause downregulation of MHC and degradation of the CD4 molecule may contribute to immune recognition of the infected cells and subsequent cytotoxicity, and lead to host resistance. In contrast, variations in Nef-dependent epitopes have been shown to facilitate immune escape and an attenuated response of cytotoxic T-lymphocytes. 71 Nef mutations have recently been linked to different rates of disease progression in children. 27 The latter studies were in a cohort of 34 children and sequence analysis was notably done only on the nef gene.
Overall, this pilot study describes the identification of six potential HIV-1 subtype C attenuated viruses with unusual genes coding for possibly defective IN, Vpr, Tat, Rev, and Nef proteins that may affect viral replicative capacity, which may be partly attributed to the long-term survival of these children. 72 Studies are ongoing to evaluate the impact of the sequence and structural alteration of these proteins on function. Although the small patient sample size is a limitation in this study, it highlights the possibility that all SP children may harbor defective or attenuated HIV-1. This warrants future studies with longitudinal follow-up of a well-characterized, large cohort to evaluate viral and host genetics together with functional immunology studies to understand the many confounding variables that contribute to slower or long-term nonprogression of vertically infected HIV-1-positive children.
Sequence Data
The six full-length genome sequences were annotated and submitted to GenBank using Sequin V5.35 (
Footnotes
Acknowledgments
We are grateful to Drs. T.M. Meyers and L Kuhn for providing the original patient samples, and for research funding from the Department of Molecular Medicine and Hematology, University of the Witwatersrand, South Africa and The International Society for Infectious Diseases (ISID) small grants. C.T.T. is a Wellcome Trust International Senior Research Fellow (076352/Z/05/Z).
Disclosure Statement
No competing financial interests exist.