
Abstract
Numerous reports have documented disturbances of sulfur metabolism in AIDS patients. There is a generalized loss of sulfur from the body, measured as cysteine and glutathione. The enzyme, cystathionase, has been shown to be greatly decreased in the liver of AIDS patients. Cystathionase is known to catalyze beta elimination of cystine giving rise to sulfane sulfur, which has potent stimulatory properties for lymphocytes. When both cystine and cystathionase are deficient in AIDS, the lymphocytes would lack this important regulator, which might be replenished by giving cystamine. Cystamine is a small disulfide that gives rise to sulfane sulfur when it undergoes oxidation catalyzed by diamine oxidase (a ubiquitous enzyme in animals). Cystamine has been shown to cause marked suppression of HIV replication in cultured lymphocytes and macrophages; the inference is that the cystamine/diamine oxidase system may replace the cystine/cystathionase system as a source of sulfane sulfur. Sulfane sulfur could have two beneficial effects: (1) it could increase the vigor and resistance of the lymphocyes and (2) it could interfere with the HIV replication process. A clinical trial of cystamine in AIDS is indicated.
There are three lines of evidence relating sulfur metabolism to HIV: (1) the depletion of sulfur in AIDS patients, (2) the special requirement of lymphocytes for sulfur factors, and (3) the HIV-suppressing effect of cystamine in vitro. These three areas are summarized and correlated below. The emerging relationships may have clinical application and it is hoped that this summary will stimulate research in this area.
Sulfur Metabolism in AIDS
Numerous reports have documented a depletion of cysteine and glutathione in plasma and leukocytes of AIDS patients (for example, see reference 1). The cause of the sulfur depletion may be related to the finding that the activity of the enzyme, cystathionase, was greatly decreased in the livers of three AIDS patients. 2 This is an important finding needing confirmation in more cases and in more tissues. A generalized decrease in cystathionase activity could have important consequences in AIDS patients: (1) the block in the transulfuration pathway would deprive the cells of methionine-derived cysteine, (2) cystathionine would be lost in the urine contributing to the observed sulfur depletion, and (3) the decreased generation of sulfane sulfur from cystine through the action of cystathionase would deprive lymphocytes of an important regulatory factor (as described below).
Sulfur Regulatory Factors for Lymphocytes
Lymphocytes seem to have a special requirement for sulfane sulfur and this is influenced by defects in the enzymes of sulfur metabolism. This is a large subject and, for brevity, the information relevant to cystathionase and to cystamine is condensed below into a series of essential findings:
Sulfur-dependent lymphocytes. Broome and Jeng were the first to show that cystamine (2-mercaptoethylamine disulfide, CA–S–S–CA) or 2-mercaptoethanol disulfide (ME–S–S–ME) at micromolar concentrations permitted the proliferation of certain malignant murine lymphocyte cell lines in vitro.
3
These cells did not proliferate in vitro in the absence of the sulfur factors.
Related Enzyme Defects. The malignant cell lines that are dependent on these growth factors in vitro have complete absence of the enzyme, methylthioadenosine phosphorylase,
4
and are greatly deficient in the enzyme cystathionase.
5,6
Normal Lymphocytes Are Also Stimulated. In addition to the absolute requirement for sulfur growth factors in malignant enzyme-defective lymphoid cells, many in vitro functions of normal lymphocytes are strongly stimulated by these sulfur compounds. This effect was first demonstrated with murine lymphocytes in the mitogen response,
7
the primary immune response to sheep red blood cells,
8
mixed leukocyte culture,
9
and B lymphocyte colony formation
10
and later in normal human lymphocytic,
11
hematopoietic,
12
and embryonic
13
cells. Other cells types do not appear to respond to these factors.
Disulfide Form Required. Cystamine and ME–S–S–ME must be in the disulfide form to support growth of dependent cells in vitro. When the reduced or sulfhydryl forms, cysteamine (CA–SH) or 2-mercaptoethanol (ME–SH), were added to dependent cell cultures and maintained in the reduced form by 1 mM glutathione, they did not support growth in the absence of fetal serum.
6
Sulfane Sulfur (So) Involved. Cystamine is a substrate for the enzyme diamine oxidase
14,15
and ME–S–S–ME is a substrate for alcohol dehydrogenase.
6
In both reactions a carbonyl group is created in a beta position to a C–S bond. The carbonyl group labilizes the C–S bond and causes the release of So (see Table 1). Other systems that slowly generate sulfane sulfur also support growth of the dependent cells; these include the beta elimination of cystine by pyridoxal as described by Cavallini et al.,
16
sulfide-treated proteins, and viscose dialysis tubing.
6
Cystine and Cystathionase. Cystathionase catalyzes the beta elimination of cystine yielding sulfane sulfur.
17,18
During the reaction, the So becomes attached to the cystathionase protein and is subsequently transferred to other sulfane sulfur carrier proteins.
19
Cystathionase has a greater affinity for cystine than for cystathionine (K
m = 0.07 mM and 15 mM, respectively).
20
It has been estimated that as much as 50% of terminal sulfur metabolism occurs by this route in liver and kidney
21
and this is probably the principal source of So in humans.
C–S Cleavage of Cysteine Alkyl Disulfides. Cysteine alkyl disulfides support growth of the sulfur-dependent cells.
6
These cysteine derivatives undergo deamination with C–S bond cleavage and the resulting R–S–SH product releases So
6
(see Table 1). At least 11 mammalian transaminases (e.g., kynurenine, glutamine, aspartate, alanine) or amino acid oxidases have cysteine C–S lyase activity of this type.
22
–24
Preformed cysteine methyl disulfide (cys–S–S–CH3) added to cell cultures and physiologically derived CH3–SH (from methionine) can also give rise to So by this mechanism.
Functions of So. Sulfane sulfur has many known functions in biological systems. It activates some enzymes, inhibits others, is involved in posttranslational modification of tRNA, and it is the source of sulfur for the synthesis of FeS clusters, molybdenum cofactors, biotin, lipoic acid, and thiamine.
25
–27
The exact function(s) in lymphocytes are not known.
Potency of So. A remarkable feature of So in biological systems is its potency. In in vitro biochemical systems it is active at micromolar concentrations and in cell culture systems, it is active at nanomolar concentrations.
25
Enzymatic Systems That Generate Sulfane Sulfur from Disulfides
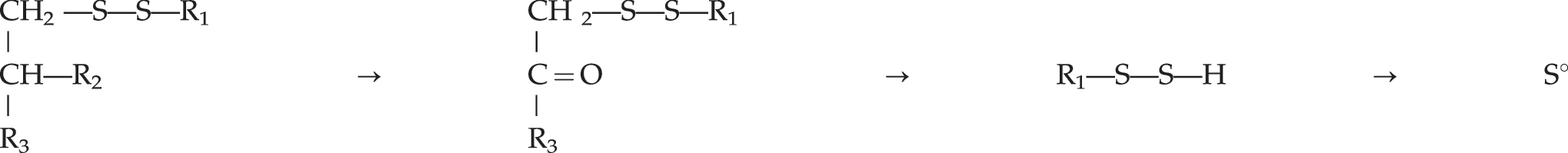
Cystamine Inhibits HIV Replication in Vitro
Based on the growth factor and immune enhancing properties of sulfane sulfur, I speculated that it might protect immune cells from the damages of HIV infection. The So- generating system, which is the easiest to handle, is cystamine/diamine oxidase since cystamine does not have an objectionable smell and since diamine oxidase is already present in serum. Therefore, in April, 1987, I submitted cystamine for testing in the National Cancer Institute In Vitro Anti-HIV Drug Screening Program. The results shown in Fig. 1, although incomplete, reveal two interesting findings. First, the CEM lymphocyte host cells used in the system were potently stimulated by cystamine resulting in 3- to 4-fold increase in cell proliferation above the expected level. Second, there was a pronounced antiviral effect approaching 100% at about 500 μM.

Effect of cystamine on uninfected and HIV-infected CEM-6 human lymphocytes. Testing was carried out by the protocol described by Weislow et al. 36 at the Developmental Therapeutics Branch of the National Cancer Institute. The vertical axis represents the number of viable cells relative to control cultures that were not virus infected and not treated with the agent (expressed as percent). This control parameter is represented by the dashed horizontal line. The upper line (dashed) shows the effect of increasing concentrations of cystamine on cells that were not infected with the virus. There was a marked stimulation of cell proliferation. The lower line (solid) shows the effect on HIV-infected cells. At the left of the chart, there was 100% killing of the host cells. As the cystamine concentration increased, the approach of the lower line to the upper line indicates that the killing effect of the virus was eliminated. It is noteworthy that cystamine promoted cell proliferation beyond the control level even in the virus-infected cells.
In 1994, Bergamini et al. published results showing that cystamine caused suppression of HIV replication in cultured human lymphocytes and macrophages. 28 Also in 1994, Kalebic and Schein reported that the cystamine prodrug, WR-151327, suppressed expression of HIV in chronically infected U1 cells. 29 Ho et al. published papers showing similar results in normal human peripheral monocytes and chronically infected promonocytic cell lines 30 and in cord blood monocytes and lymphocytes. 31 It is important to note that Ho et al. showed that while the disulfide form, CA–S–S–CA, was effective in suppressing HIV, the sulfhydryl form, CA–SH, was not effective. This finding correlates with the requirement for the oxidized disulfide form for growth factor activity in lymphocyte cultures (cf. item 4 above).
Follow-Up
Following these reports, a clinical trial might have been anticipated. Instead cystamine disappeared from the HIV literature. The pharmacokinetic data for a clinical trial are already in place since cystamine has been tested for radioprotection and because cysteamine is now being tested in clinical trials for treating nephropathic cystinosis, Huntington's disease, neuronal ceroid lipofuscinosis, and nonalcoholic steatohepatosis (website of Raptor Pharmaceutical Corporation,
Another aspect of this subject that needs development is the inactivation of cystathionase in tissues of AIDS patients. Since the cystine/cystathionase system is a source of So and since So seems to be especially stimulatory to lymphocytes, it is conceivable that inactivation of cystathionase is a crucial element in the lymphocytopathic effect of HIV. There may be a precedent for this with another retrovirus; Livingston et al. reported that thymic tumor induced in AKR mice by the retrovirus type C (M-MuLV) contained 5- to 10-fold lower specific activity of cystathionase than control tissue. 5 The decreased cystathionase activity undoubtedly disturbs sulfur metabolism in cells and deprives them of So as well as cysteine and, indeed, a requirement for both cysteine 5 and So 6 has been demonstrated in cystathionase-deficient cells. There may be a natural means of protection from this damage. Thus, several genetic variants of cystathionase are known 32 and probably many more exist. These mutations markedly affect the kinetic properties of the enzyme. 33 Certain variants may resist inactivation during HIV infection and this, in turn, could explain the variable progression of HIV infection or even HIV resistance.
The mechanism of cystathionase inactivation in retrovirus-infected cells also needs to be examined. This might require only one gene product. For example, cyanoalanine is a potent cystathionase inhibitor 34 that can be formed from asparagine by the enzyme asparagine hydro-lyase (EC 4.2.1.65, nitrile hydratase) in plants, bacteria, and fungi. 35 The encoding of this enzyme in viruses seems not to have been examined.
Footnotes
Disclosure Statement
No competing financial interests exist.