
Abstract
Monocytes serve as a systemic reservoir of myeloid precursors for the renewal of tissue macrophages and dendritic cells (DCs). Both monocytes and dendritic cells can be infected with HIV-1. Circulating DCs are believed to be derived from a common precursor of monocytes, or, in the case of inflammatory challenge, from monocytes directly. Because there are fewer infected blood monocytes than infected cells after differentiation, we hypothesized that the majority of HIV-1 infection in circulating DCs occurs via direct viral binding to their CD4 and coreceptors after differentiation. We isolated monocytes at one time point and circulating dendritic cells at a second time point from the blood of HIV-1-infected patients. Proviral DNA was isolated from DCs and monocytes, and the C2-V4 region of the HIV-1 env gene was cloned and sequenced. Phylogeny, nucleotide distances, and glycosylation patterns of the env gene were performed. The phylogenetic trees revealed that viral forms from the monocytes clustered distantly from the quasispecies derived from circulating DCs. The nucleotide distances and differing glycosylation patterns suggest that the infection of DCs is independent of the infection of the monocytes.
M
Blood DCs are the most potent antigen-presenting cells derived from monocytes; once infected with HIV-1 they can transmit the infection to CD4+ T cells. 16 It has been shown that DC subtypes (myeloid DCs and plasmacytoid DCs) can transmit HIV-1 to T cells more rapidly than infection by the free virus via cis infection using the CD4 receptor as well as CCR5 and CXCR4 coreceptors. 17–18 However, another nonpredominant pathway of transmission exists. The trans infection occurs when HIV-1 enters a DC by the binding of gp120 to mannose C-type lectin receptors such as DC-SIGN. 16,19,20 These interactions allow the virus to be transferred to T cells so that productive viral replication can take place. 16
Blood dendritic cells and monocytes can share a common precursor or else monocytes can differentiate into dendritic cells in inflammatory conditions such as those often found with an uncontrolled HIV-1 infection. It is crucial to identify how DCs become infected with HIV-1 to better understand the role of DCs over the course of infection. Based on the dichotomy of a lower fraction of infected blood monocytes compared to the fraction of infected cells after differentiation, we hypothesized that the HIV-1 infection of circulating DCs is produced directly from viral binding to their CD4 and coreceptors and it is not acquired before differentiation. If the viral infection of DCs is independent of that of their precursors, control of viremia in dendritic cells will become less challenging. However, if monocytes infected with HIV can differentiate into DCs while maintaining the virus in their genome, another pathway of infection of DCs would be identified.
Three HIV-1-infected individuals were enrolled in this study. Blood samples from each IRB-consenting individual were obtained by peripheral phlebotomy at two time points. Peripheral blood mononuclear cells (PBMCs) were separated from 63 ml of whole blood by density gradient sedimentation on Histopaque-1077 (Sigma). In the first two patients (HF1, HF2) monocytes were purified from fresh PBMCs by using CD14 MicroBeads (Miltenyi Biotec Inc., Auburn, CA) and passed through the magnetic column following the Miltenyi protocol. For patient HF3 the monocyte isolation was performed by using a combination of the monoclonal antibodies CD14+ and CD16+ (BD Biosciences, San Jose, CA) and then sorted using a FACSCalibur Aria (BD Systems). The isolation of DCs was performed 16 weeks after the monocyte sample was obtained from patient HF1, 10 weeks for patient HF2, and 2.5 weeks for patient HF3.
The time points selected for the sample collections took into consideration that monocytes are in blood for a period of days before migrating to tissues whereas the circulating dendritic cells are final stage cells that survive for weeks to months. These samples were collected using time frames optimal for finding a relation between the proviral forms. The broad time frame selected for the first patient provided us with the maximum time frame to find a relation between these two groups of cells. This time frame was reduced for the other two patients to determine whether this relation was maintained or changed. These time frames were predicted to provide the opportunity to identify whether the viral infection persisted beyond differentiation. Circulating DCs were purified from fresh PBMCs according to the Dendritic Cell Isolation Kit II protocol (Miltenyi Biotec, Inc., Auburn, CA). CD4+ T cells were isolated using CD4+ T Cell Kit II (Miltenyi Biotec) for all time points, except for samples from HF1 and HF2 time 2, from which CD25 MicroBeads were also used to obtain the CD25– subset of CD4+ T cells (resting T cells). Isolated cell suspensions yielded average purities of 98% for monocytes, 94% for DCs, and 97% for T cells based on flow cytometry, using FACSCalibur and CellQuest software (BD Biosciences, San Jose, CA). To determine the presence of contaminating cells in the DC separation, the Lin marker was used. The Lin marker registers positive for CD3, CD14, CD16, CD19, CD20, and CD56, a population that includes monocytes and T cells. This marker showed an average of 4.0% for Lin-reactive cells.
Genomic DNA was isolated from DCs, monocytes, and CD4+ T cells using the QIAmp DNA Micro kit (QIAGEN, Inc., Valencia, CA). Amplification of the HIV-1 C2V4 env region was achieved through nested polymerase chain reaction (PCR), using HotStar Taq DNA Polymerase (QIAGEN, Inc.) with the external primer pair ED12 and ED31 (5′-AGTGCTTCCTGCTGCTCCCAAGAACCCAAG-3′ and 5′-CCTCAGTCATTA CACCAGGCCTGTCCAAAG-3′), respectively, and the internal primer pair CV3-R and CV3-F (5′-TGATGGGAGGGGTATACATT-3′ and 5’-CTGTTAAATGGCAGT CTAGC-3′), respectively. The final amplicon (525 bp) was electrophoresed along with a low-molecular-weight marker in 1.2% agarose, stained with ethidium bromide, and visualized under UV light for the determination of molecular weight.
HIV-1 RNA was isolated from plasma according to the procedure of the QIAmp Viral RNA Mini Kit (QIAGEN, Inc., Valencia, CA). Nested PCR was performed using the OneStep RT-PCR Kit and HotStar Taq DNA Polymerase (QIAGEN, Inc.) with the same primer combinations. PCR products were ligated and cloned using the PCR-TOPO TA Cloning Kit (Invitrogen, Carlsbad, CA). Plasmid DNA was purified using QIAprep Spin Miniprep columns (Qiagen Inc., Valencia, CA), and all HIV-1 env gene clones were sequenced using an Applied Biosystems 3130x Genetic Analyzer.
Nucleotide sequences were aligned and hand edited with BioEdit version 7.0.5.2.
21
Analyses of a total of 152 cloned nucleotide sequences from DCs, monocytes, CD4+ T cells, and plasma were carried out on an approximately 480-bp fragment on the env gene spanning the C2-V4 region. Pairwise nucleotide sequence distances employing the Kimura two-parameter model and a transition/transversion ratio of 2.0 were computed using MEGA version 3.1.
22
Phylogenetic reconstruction was generated using neighbor joining for 1000 bootstrapped data sets. The final graphic output was created with MEGA X.
23
Phylogenetic analyses were performed using the program MacClade.
24
Potential N-linked glycosylation sites were determined using the N-Glycosite tool from the Los Alamos HIV Sequence database,
Viral loads and CD4+ T cells counts were measured from blood samples. Each patient and time point had detectable viral loads (Table 1).
Phylogenetic analyses of the HIV-1 C2V4 env region from patients HF1, HF2, and HF3 showed separate, distinct monophyletic groups of nucleotide sequences for each patient with 100% bootstrap values, excluding the possibility of cross-contamination. The phylogenetic trees revealed that viral forms in resting CD4+ T cells clustered consistently with the forms found in circulating DCs in patients HF1 and HF2 (Figs. 1 and 2). In addition, plasma viral RNA quasispecies and monocytes clustered separately from those derived from resting CD4+ T cells and DCs. Similarly, in patient HF3, from whom the two samples were obtained 2.5 weeks apart, the phylogenetic tree showed viral forms from the monocytes did not cluster with the form of the DCs. For this patient, these forms from the monocytes clustered more closely to those from CD4+ T cells and the DCs cluster even more closely to the forms from plasma (Fig. 3). For patient HF3, CD16+ monocytes were isolated using a double positive selection for CD14+/CD16+. CD16+ monocytes have a higher presence of CD4+ and HIV coreceptors and represent a potential harbor for the virus. 26,27
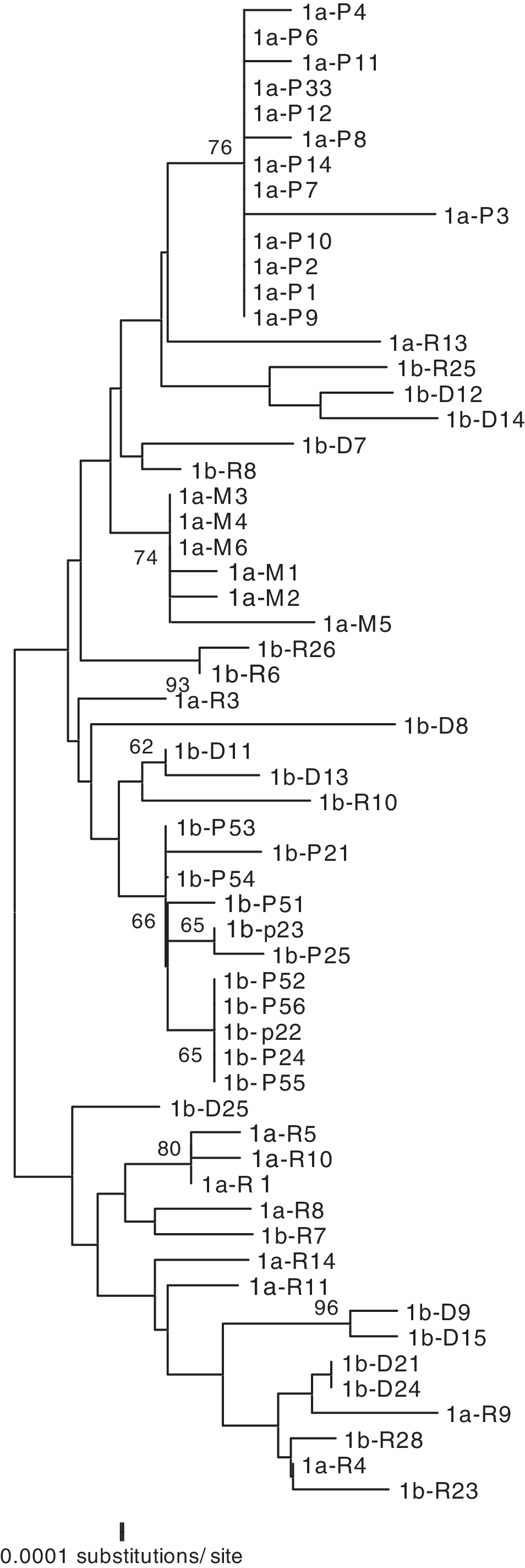
Phylogenetic reconstruction of HIV-1 gp120 env C2-V4 nucleotide sequences from patient HF1. Reconstruction was generated by the NJ method. Clones were labeled
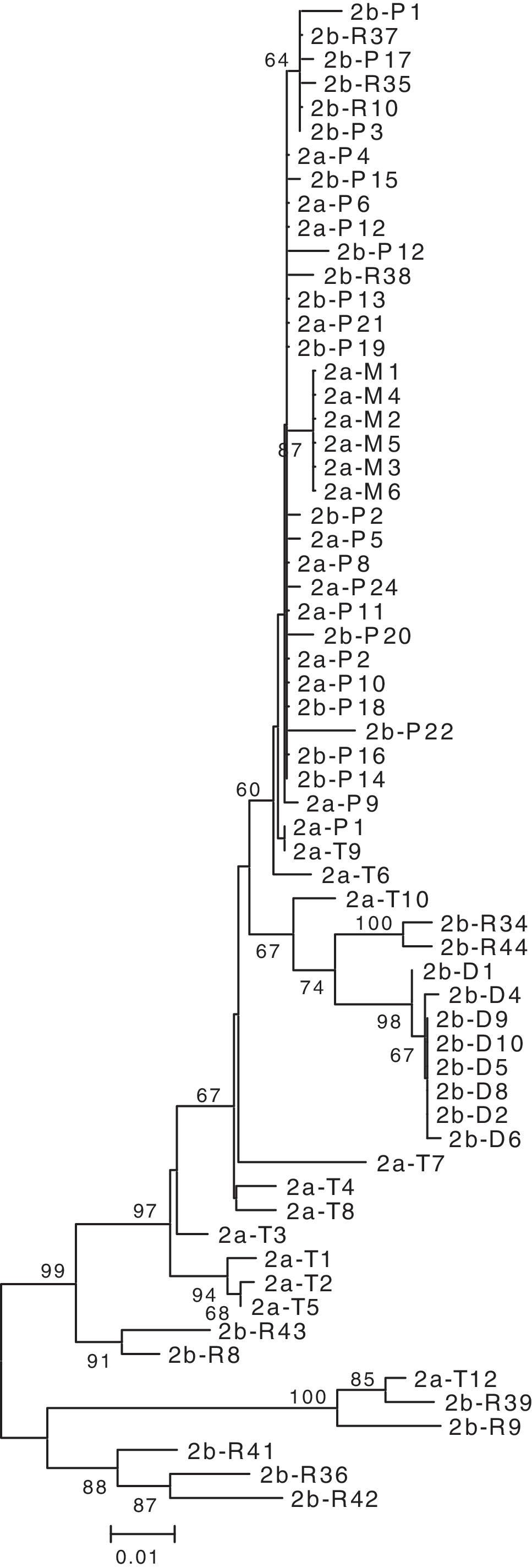
Phylogenetic reconstruction of HIV-1 gp120 env C2-V4 nucleotide sequences from patient HF2. Clones were labeled as described in the legend to Fig. 1.
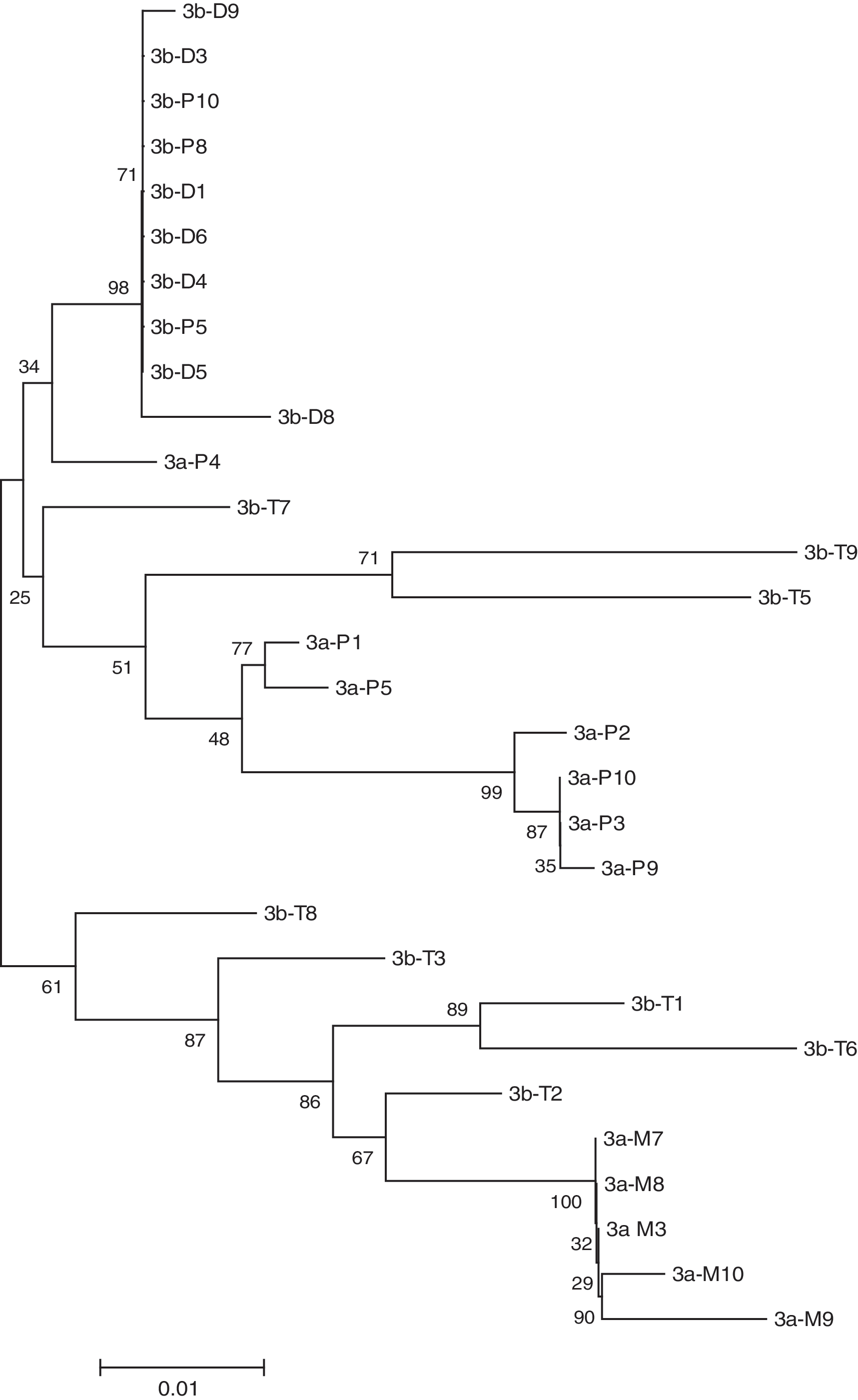
Phylogenetic reconstruction of HIV-1 gp120 env C2-V4 nucleotide sequences from patient HF3. Clones were labeled as described in the legend to Fig. 1.
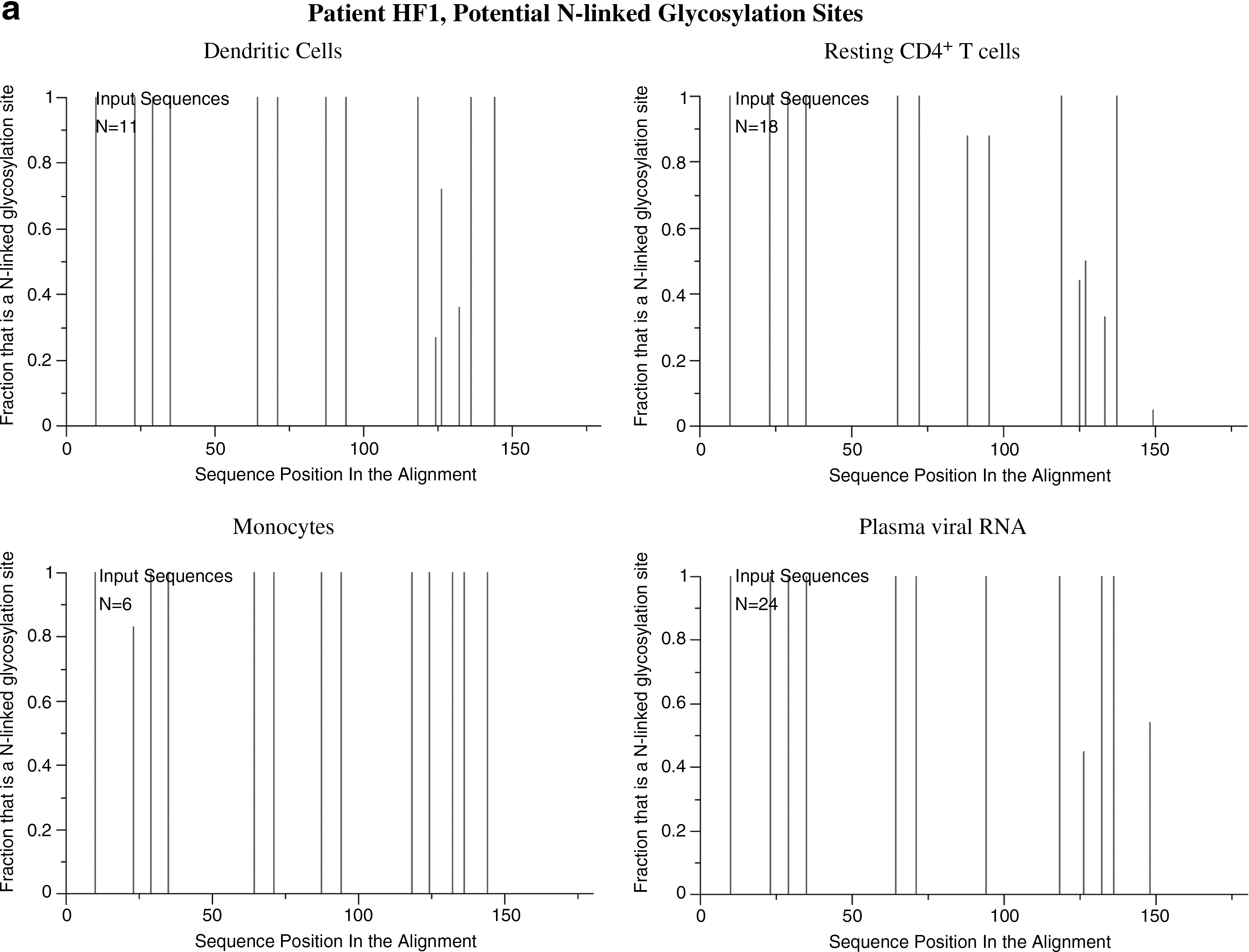
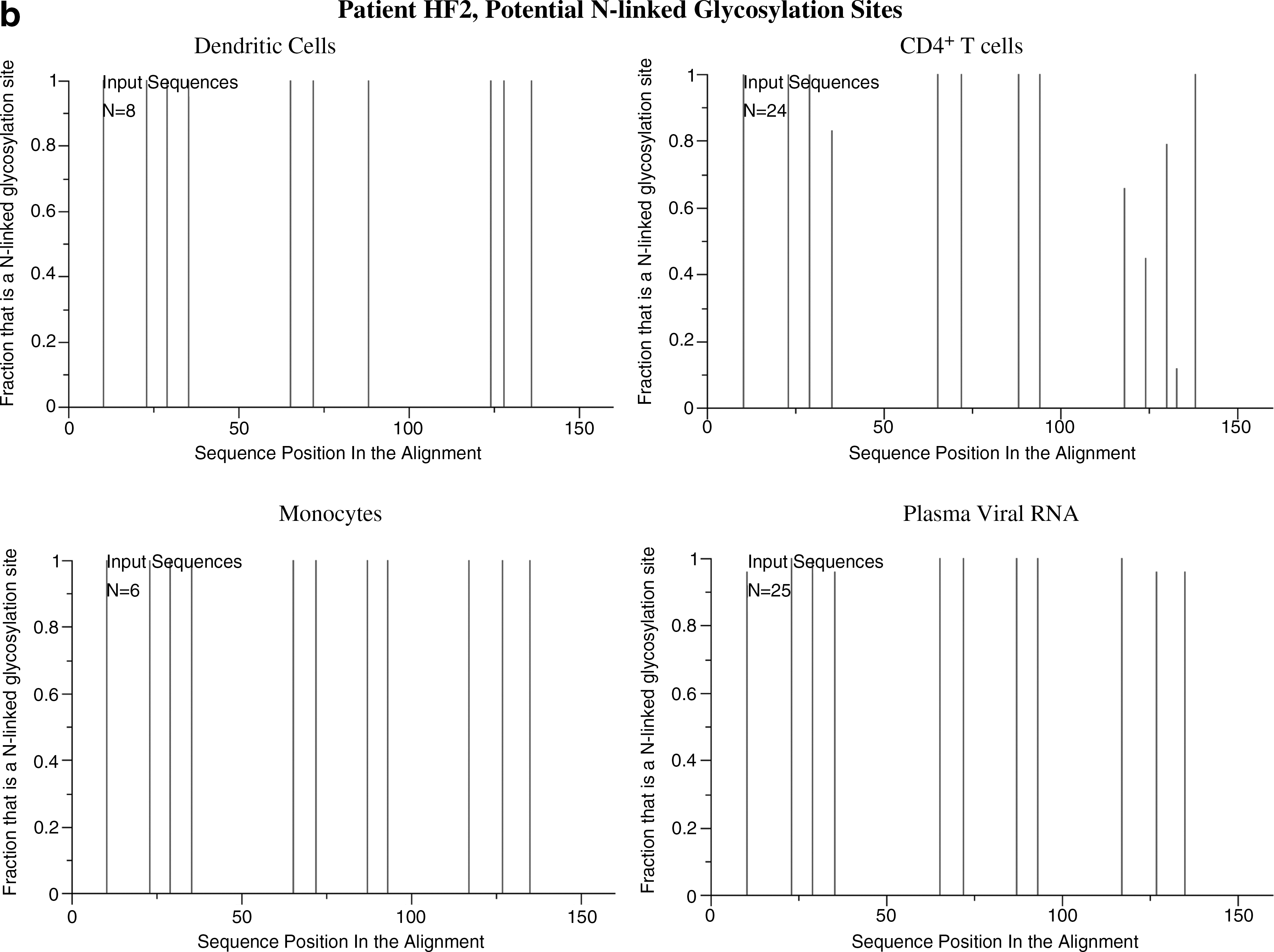
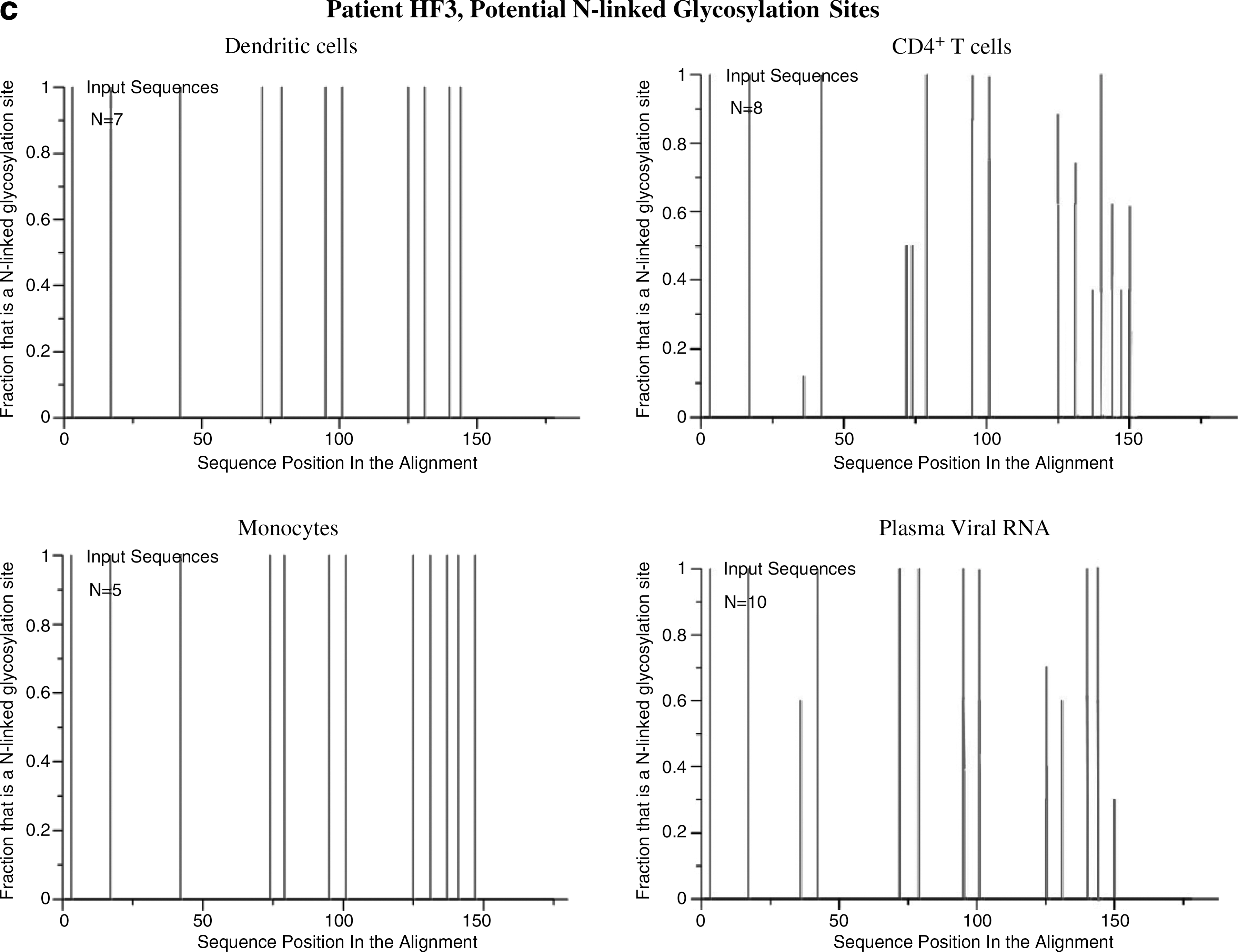
The results were similar after the separation of CD16+ monocytes suggesting this form of monocyte may harbor the virus. In all patients the intrasample viral quasispecies nucleotide distances from the monocytes and DCs showed minimal variability. Only the CD4+ T cells showed a large internal variation in nucleotide distances (Table 2). The nucleotide distances between the viral forms derived from various sources support the findings presented in the phylogenetic trees. There are viral forms from the dendritic cells more closely related to those from plasma and T cells than to those from monocytes for each patient (distance ranges, Table 3). The glycosylation patterns showed differences between each group of cells (Fig. 4). Viral forms from monocytes and DCs revealed little variation in glycosylation patterns, in contrast to the multiple changes observed in quasispecies of CD4+ T cells and plasma RNA in all patients (Table 4).
(
Intrasample Kimura two-parameter nucleotide distances. Values are expressed in percent.
Intrapatient Kimura two-parameter nucleotide distances between sequences derived from various cell types and plasma. Values are expressed in percent. Sequences were derived from DCs, dendritic cells; M, monocytes; P, plasma viral RNA; T, CD4+ T cells.
The total number of N-linked glycosylation sites present in the C2–V4 sequences derived from the various cellular and plasma aliquots and the number of those N-linked glycosylation sites that showed heterogeneity.
Based on the phylogenetic analyses, we concluded that viral forms in circulating DCs are not closely related to the quasispecies found in blood monocytes from an earlier time point. This implies that the viral form in the DCs come from other sources and not from the circulating monocytes, suggesting an independent process of infection. The change in the isolation time frame created some minor shifts in the proviral phylogeny. Nevertheless, the clear distinction between monocytes and DCs persisted. Viral forms in CD4+ T cells comprise a heterogeneous population of quasispecies, as suggested by the broad range of nucleotide distances. Such a huge population of cells has the capacity to harbor viral quasispecies along the whole course of infection. However, viral forms from DCs in all patients showed nucleotide distances that are longer than those obtained from previously isolated monocytes.
For patient HF1, the viral forms from monocytes clustered with forms from CD4+ T cells and plasma RNA revealing short nucleotide distances between them. For patient HF2, the quasispecies between monocytes and plasma also showed short nucleotide distances. This suggests that a possible viral trafficking exists between these cell types and plasma. In contrast, monocytes and DCs showed longer distances between them, excluding the possibility of a recent common ancestor. In all patients the forms in DCs as well as in monocytes reflected intracellular homogeneous populations of quasispecies. Analysis of the phylogenetics trees using MacClade points to a strong relationship between the viral forms from the DCs and the T cells but shows no relation between those from the DCs and the monocytes. It appears that the virus moves from the DCs to the T cells but not from the monocytes to the DCs.
N-linked glycosylation of HIV env can enable evasion from neutralizing antibodies. 28 Analyses of shifts in sequence-predicted glycosylation can be used as a tool to evaluate the homogeneity or heterogeneity of viral populations within each analyzed host source, to determine whether a correlation existed between forms during the course of infection and as an indirect evaluation of immune pressures. 29 The N-glycosylation analysis was limited to the sequenced portion of the viral envelope and showed only the possibility of glycosylation at those specific sites. Viral forms in DCs and in CD4+ T cells from patient HF1 showed multiple changes in their glycosylation patterns (Fig. 4). This implies that immunological pressures were exerted on these populations over a long time. Among DC populations, only DCs from patient HF1 showed variants in the glycosylation of virus sequences. This could be due to the broader range of viral forms from the DCs from this patient with sequences distances with variants differing by up to 3.7% compared to 0.4% and 1.4% for patients HF2 and HF3, respectively.
When comparing glycosylation patterns of the populations in combination, no evident association was found between forms in monocytes and those in DCs. On the other hand, forms from monocytes showed the same glycosylation patterns as some viral quasispecies in plasma RNA, further supporting the possibility of trafficking between these two compartments. As the time between collections of blood samples for patient HF3 was less than 3 weeks, the glycosylation patterns between the DCs and monocytes appear to be related because they were under similar immune pressure.
The blood dendritic cells are a small portion of the PBMCs constituting only about 0.5–2% of the entire PBMC population. 30 Both myeloid and plasmacytoid types of dendritic cells are susceptible to infection with HIV. 31 Most dendritic cells reside outside of the blood compartment, with murine studies suggesting they may have differentiated from monocytes after the monocytes migrated to inflamed tissues. 26 It is not practical to sample these DCs, but inasmuch as blood is the most accessible tissue it can provide us with a limited window into the dynamics of the infection between monocytes and dendritic cells. Because the majority of dendritic cells are tissue associated, it is possible they may be infected from their precursors.
The viral forms obtained from the blood DCs of patients HF2 and HF3 each displayed a narrow range of variation. This could be due to the successful amplification of a limited number of infected cells and could skew the results by producing viral sequences that are not representative of infected blood DCs. However, with the large blood volume (63 ml) used, there should have been in the neighborhood of 106 blood dendritic cells. Even with a small fraction of this population being infected, there would have been thousands of proviral DNA templates.
There are two possible sources for circulating DCs. 32,33 Circulating DCs can share a common precursor with monocytes and then follow different pathways in their development. With this consideration in mind, the infection of this cell type could happen at any stage of their development and can come from different sources. However, when a patient is experiencing proinflammatory challenges as can occur in the case of uncontrolled HIV disease, monocytes directly differentiate into circulating dendritic cells. In this case, the different viral forms would mean that the circulating DCs were infected after differentiation from their monocyte precursors.
Although we found different viral forms in the monocytes and circulating dendritic cells, this study looked at only three patients with varied time frames. The varied time frames were employed to determine whether the viral forms from the short-lived monocytes would appear and remain in the longer-lived circulating dendritic cells; the range of times did not provide multiple samples from the same conditions. These patients may not be typical. HAART is often successful in maintaining viral loads below detection limits and in doing so can narrow the cell types that maintain a reservoir of virus. Each of the patients in this present study had substantial viral loads at each time point (range 319–21942 virions/ml).
In summary, in three patients, we found distant monophyletic clusters between monocytes and DCs in their phylogenetic trees (see Supplemental Figs. 1–9; see
The nucleotide sequences from monocytes, T cells, and plasma included in this report have been published previously 7 and were given GenBank accession numbers (DQ387660–DQ387853). Additional sequences from DC and CD16+ monocytes were given accession numbers (DQ647317.1)
Footnotes
Acknowledgments
We acknowledge Pablo López for assistance in flow cytometric analysis and Drs. Iván Meléndez and Gladys Sepúlveda and their clinical staff for patient recruitment, blood collection, and follow-up of the patients. We are also indebted to the patients who donated blood for this study. This work was supported by NIH Grants MBRS S06-GM008239 (to E.L.) and RCMI 2G12RR003050-20 (PSM Molecular Biology Core). Manuel Vázquez and Irvin M. Maldonado contributed equally to this work.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.