
Abstract
We have studied 65 HIV-1-infected untreated patients recruited in Caracas, Venezuela with TCD4 counts ≥350/μl. The reverse transcriptase and protease sequences of the virus were sequenced, aligned with reference HIV-1 group M strains, and analyzed for drug resistance mutations. Most of the viruses were subtype B genotype in both the protease and RT genomic regions. Five of the 62 virus isolates successfully amplified showed evidence of recombination between protease and RT, with their protease region being non-B while their RT region was derived from subtype B. Four strains were found bearing resistance mutations either to NRTIs, NNRTIs, or PIs. The prevalence of HIV-1 isolates bearing resistance mutations was therefore above the 5% threshold of WHO.
Introduction
H
HAART regimens have been adopted by the AIDS National Program in Venezuela since 1999. By the end of 2005, approximately 15,000 patients were receiving HAART in Venezuela, which represented 84% of adults in need of therapy according to the program guidelines. 8 The available HIV drug resistance reports from the country show that a 3-fold increase of resistant viruses to RTIs, now representing 10% of the clinical samples, has been observed between 2001 and 2007. 3,7 Certainly this increase implies a significant rate of HIV drug resistance transmission. The present study monitored the prevalence of genotypic drug resistance in a sample of asymptomatic untreated patients recruited at an outpatient network in Caracas according to the WHO survey threshold method recommendations. 9
Materials and Methods
All individuals enrolled in the study signed the informed consent form and conformed to the following criteria: adults with a positive HIV enzyme-linked immunosorbent assay (ELISA) within the last 3 years verified through Western blot, naive to antiretroviral treatment, and with a TCD4 lymphocyte count ≥350 cells/μl. Blood was collected in ethylenediaminetetracetic acid (EDTA) tubes and plasma was separated and stored at –80°C. Plasmas were thawed, homogenized, and applied to spots of Schleicher and Schuell 903 (now Whatman 903) filter paper cards with five spots of 50 μl of plasma. The samples were allowed to dry at room temperature for 1 h in a vertical position and were placed in plastic bags with desiccants. They were stored at 4°C before being carried at room temperature to Bordeaux, France during a 12–15 h journey. The spots were cut with scissors and eluted in 220 μl/spot of a buffer containing phosphate-buffered saline (PBS), Tween 20, and fetal calf serum as previously described. 10 The samples were then agitated at 4°C for 1 h before being vortexed and 140 μl of the final elution was used for RNA extraction using the QIAamp Viral RNA Mini Kit (Qiagen). The RNA was used in reverse transcription polymerase chain reaction (RT-PCR) of reverse transcriptase (RT) and protease (Prot), respectively, using two sets of primers in a GeneAmp PCR System 9700 (Applied Biosystems, Foster City, CA) thermal cycler. The outer and inner primers for RT and Prot are described in our previous publications. 10,11 The fragments obtained were sequenced on both strands using the CEQ DTCS Quick Start kit on an automated sequencer Beckman CEQ 2000 DNA Analyser System in the Virology Department of the University Hospital of Bordeaux as previously described. 10,11
The derived nucleotide sequences of the RT and Prot regions were aligned by the Clustal W 1.74 alignment program with reference sequences of HIV-1 M, N, and O groups (plus SIV-CPZ for the protease tree) from the HIV Database Subtype Reference Alignment (
Results
Seventy HIV-1-infected naive outpatients were recruited in Caracas; five patients were excluded from the study (two with no results of TCD4 counts, two with TCD4 counts below 350/μl, and one lost to follow-up); the results were therefore valid for 65 patients. The population studied was as follows: 51 men (78.46%), 14 women (21.54%), 18–58 (median 32.57) years old, TCD4 range from 350 to 1320 (average 551, median 531) cells/μl, viral loads ranging from 763 to 500,000 (average 53,164, median 19,337) RNA copies/ml, and an estimated first positive HIV ELISA ranging from 0 to 132 months (average 14.86, median 6 months).
Both Prot and RT were successfully amplified for 62 HIV-1 isolates of the 65 patients remaining in the study. For subtyping purposes only, nucleotide sequences of RT or Prot fragments from the three other viral isolates were also considered in the phylogenetic analysis. All analyzed RT sequences clustered with subtype B (Fig. 1). All but five Prot sequences were of subtype B; the five non-B Prot sequences were assigned as B/F1 recombinant (two sequences), CRF06_cpx (2), and CRF01_AE-like recombinant (1) (Fig. 2). As CRF06_cpx is rarely reported other than in Africa, we investigated whether the two identified strains could have a common ancestor and/or a particular recombination pattern. However, no unexpected breakpoints were seen when scanning both protease sequences and the highest blast scores corresponded to different CRF06_cpx reference sequences (data not shown). We then reanalyzed both RT subtype B fragments in the context of previously known B strain sequences from Venezuela and found that they fall in distinct clusters of a maximum likelihood tree (data not shown). Therefore the two B/CRF06_cpx genomes were proven to be unlinked isolates, which is also consistent with patient's records of unrelated infections. More extensive data would be necessary to classify these mosaic genomes. However, we can conclude that the variability of HIV-1 in Venezuela is increasing together with recombination of different strains within a background of subtype B.
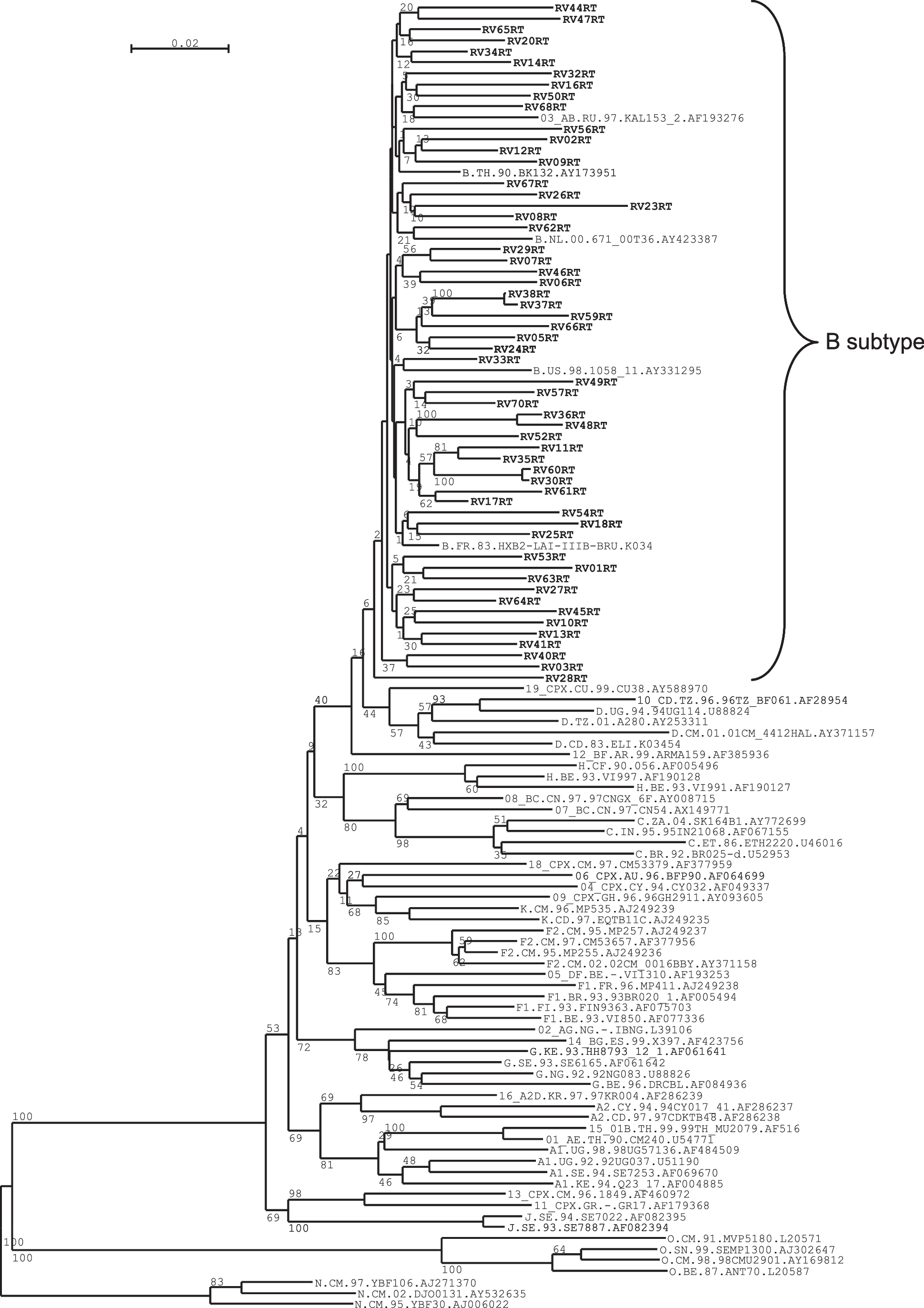
Neighbor joining tree of RT sequences. Subtype reference strains were included and the N strain was used as the root.
Neighbor joining tree of PROT sequences. Subtype reference strains were included and the N strain was used as the root.
Analysis of the RT sequences exhibited polymorphic substitutions at positions that, according to the ANRS algorithm, are not associated with resistance (E122, D123, I135, K173, T200, E204, Q207, R211, V245); amino acid substitutions at positions of resistance were quite rare; more interestingly, two resistance mutations to nonnucleoside reverse transcriptase inhibitors (NNRTIs) (K101E and K103N) were noted in separate isolates of subtype B while a third subtype B was found bearing two resistance mutations to nucleoside reverse transcriptase inhibitors (NRTIs) (D67N and K219Q) (Table 1).
RT, reverse transcriptase; Prot, protease.
Analysis of Prot sequences revealed frequent polymorphic amino acid substitutions at some positions of secondary resistance (M36, L63, V77); only one isolate (which did not bear resistance mutations to NRTIs and NNRTIs) exhibited major resistance mutations to PIs (L33F, M46I, V82T, I84V, and L90M) (Table 1).
Discussion
Most of the HIV-1 isolates are of subtype B; these results are consistent with previous data from the country showing that B viruses are largely predominant, but novel recombinant forms to the local epidemic that need more extensive characterization have been observed.
Some studies of resistance mutations have been done in Venezuela using proviral DNA from both treated and untreated patients, but all of them included a small number of samples from untreated individuals and the results must be considered qualitative and not quantitative even if the authors gave percentages of prevalence of isolates with drug resistance mutations. Delgado et al. 3 estimated 3% RT mutations in untreated patients and reported a case of resistance to PIs. Dieudonne et al. 6 emphasized the frequency of polymorphism at positions of seconday resistance in Prot of both treated and untreated patients; they found resistance mutations to NRTIs and NNRTIs in one untreated individual of their study (7.7%). Bouchard et al. 7 within our collaborative group confirmed the polymorphism of Prot, particularly at position 77, and estimated to 10% the percentage of isolates with resistance mutations to NRTIs and NNRTIs.
The study here exhibits some differences: (1) It has been conducted using plasma samples, whereas those cited above have been carried out on peripheral blood mononuclear cells (PBMCs) and proviral DNA (it must be pointed out that either compartment seems suitable for the detection of mutations) 12,13 and (2) it has been undertaken following the WHO criteria and the results should therefore be considered more reliable. When we studied the RT and Prot sequences, we observed polymorphic substitutions, particularly in the Prot region, as already reported. 6,7 Four isolates (all of B subtype) exhibited drug resistance mutations: two to NNRTIs, one to NRTIs, and one to PIs . Overall, these data show that the prevalence of HIV-1 isolates bearing resistance mutations to the drug classes used in the country is above the threshold of 5% and that a longitudinal survey of transmitted resistance will be necessary in a next future.
Footnotes
Acknowledgments
This work has been supported partly by the Fundación Polar from Venezuela, the Sociedad Venezolana de Infectología, and the French Ministry of National Education and Research through the quadriennal contract to EA 2968. We are grateful to all volunteers who participated in the study.
Disclosure Statement
No competing financial interests exist.