
Abstract
Humanized Rag2-/-γc -/- mice (Hu-DKO mice) become populated with functional human T cells, B cells, and dendritic cells following transplantation with human hematopoietic stem cells (HSC) and represent an improved model for studying HIV infection in vivo. In the current study we demonstrated that intrasplenic inoculation of hu-DKO mice with HIV-1 initiated a higher level of HIV infection than intravenous or intraperitoneal inoculation, associated with a reciprocal decrease in peripheral CD4+ T cells and increase in peripheral CD8+ T cells. HIV infection by intrasplenic injection increased serum levels of human IgG and IgM including human IgM and IgG specific for HIV-1 gp120. There was a significant inverse correlation between the level of HIV-1 infection and the extent of CD4+ T cell depletion. Highly active antiretroviral therapy (HAART) initiated 1 week after HIV-1 inoculation markedly suppressed HIV-1 infection and prevented CD4+ T cell depletion. Taken together, these findings demonstrate that intrasplenic injection of hu-DKO mice with HIV is a more efficient route of HIV infection than intravenous or intraperitoneal injection and generates increased infection associated with an increased anti-HIV humoral response. This animal model can serve as a valuable in vivo model to study the efficacy of anti-HIV therapies.
Introduction
I
Deletion of the Rag2 recombinase and interleukin (IL)-2 receptor gamma chain [gamma common (γc) chain] genes severely compromises the immune system of Rag2-/-γc -/- mice (DKO mice) by preventing the maturation of functional T cells, B cells, and natural killer (NK) cells. 14,15 As a consequence of this severe immunodeficiency, DKO mice are highly tolerant for human cell transplantation and support the subsequent differentiation of human hematopoietic stem cells (HSC) into functional lymphoid and myeloid cells 16,17 after transplantation into sublethally irradiated newborn mice. 18 These humanized DKO mice, hu-DKO mice, form primary and secondary human lymphoid tissues, develop functional immune responses, 16,17 and are susceptible to HIV infection. 19 –24 However, studies are still limited regarding their capacity to develop immune responses against HIV after infection and their use to evaluate the efficacy of antiretroviral therapy.
In the present study, we demonstrate that after intrasplenic injection of HIV-1, hu-DKO mice develop productive HIV infection that is associated with reciprocal depletion of the human CD4+ T cell population and increase in the human CD8+ T cell population that correlates with the degree of HIV infection. The plasma of the HIV-infected hu-DKO mice contained human IgG and IgM directed to gp120. Provision of highly active antiretroviral therapy (HAART) soon after infection potently suppressed productive HIV infection and prevented CD4 depletion. Taken together, these results further validate the applicability of hu-DKO mice as a model for the development of HIV-specific therapies.
Materials and Methods
Isolation of hematopoietic stem cells from fetal liver
Human fetal liver was obtained from the Albert Einstein College of Medicine Human Fetal Tissue Repository in accordance with institutional review board regulations. Cells were isolated from the disrupted fetal liver by density gradient centrifugation over Ficoll-Paque Plus (GE Health Care, Piscataway, NJ). Some of the harvested mononuclear cells were enriched for CD34+ cells by positive selection after incubation with immunomagnetic beads (CD34 Progenitor Cell Isolation Kit, Miltenyi Biotec Inc., Auburn, CA) and passage through an autoMACS Separator (Miltenyi Biotec Inc., Auburn, CA). The cells were either immediately used for transplantation or frozen until newborn mice were available for injection.
Generation of hu-DKO mice
Rag2-/-γc -/- mice on a C57BL/10 background (DKO mice) were bred and maintained under biocontainment conditions. Newborn mice (within 1–7 days of birth) were sublethally irradiated (3.50 Gy) and within 6 h intrahepatically injected with fetal liver-derived human hematopoietic progenitor cells, either with CD34 enriched cells (0.1–2.5 × 106 cells) or unfractionated mononuclear cells (0.5–70 × 106 cells) as described. 16 All the studies were performed under a protocol approved by the Animal Institute for Animal Studies at Albert Einstein College of Medicine.
Analysis of cell populations by flow cytometry
Human leukocyte engraftment was quantified in the peripheral blood at indicated time points after transplantation to determine the level of human cell engraftment. Selected mice were sacrificed at different time points after infection, and the spleens, bone marrow, thymus, and lymph nodes were harvested. The population of human cells was quantified by staining with antihuman monoclonal antibodies (mAb) directed to CD34, CD45, CD8, CD4, CD19, CD11b, CD14, and CD69 (BD Biosciences, San Jose, CA) and measuring expression of these markers on an LSRII (BD Biosciences) using the Flowjo Analysis Program (Tree Star, Ashland, OR).
HIV-1 infection of hu-DKO mice and analysis
Rag2-/-γc -/- mice reconstituted with human HSC were infected with 100 μl of HIV-1JR-CSF viruses (800, 3600, or 8000 TCID50) at 9 weeks and older by intrasplenic injection as described. 25 A control mouse group was injected with heat-inactivated (30 min at 56°C) HIV-1JR-CSF virus by intrasplenic injection. 26 To compare the effects of infection by different injection routes, other groups of hu-DKO mice were injected with the same dose of HIV-1JR-CSF virus by either the intraperitoneal or intravenous route. Plasma HIV-1 RNA was quantified using VERSANT HIV-1 bDNA 3.0 (Bayer, Tarrytown, NY) according to the manufacturer's instructions. Frozen plasma from mice in the same experimental group was pooled and diluted in phosphate-buffered saline (PBS) to achieve the 1 ml volume required by the assay. The number of HIV-1-infected cells in the mouse spleens was quantified by limiting dilution coculture of mouse splenocytes with HIV-1-naive donor PHA/IL-2-activated human peripheral blood mononuclear cells (PBMCs) (1 × 106/well). HIV-1 p24 antigen in the supernatant was measured 7 days later by a modified enzyme-linked immunosorbent assay (ELISA) as previously described. 25
Identification of HIV-infected cells in the spleens of the HIV-challenged mice by intracellular staining for p24 antigen and flow cytometry
Spleen cells harvested from the mouse spleens were pooled in each experimental group and suspended in FACS buffer (PBS with 1% sodium azide). The cells were stained with antihuman CD45 and CD4 mAbs prior to cell fixation with 4% paraformaldehyde. Cells were then incubated for 4°C for 1 h, followed by washing with FACS buffer. Phycoerythrin (PE)-conjugated anti-HIV p24 antibody (clone KC57-RD1, Beckman Coulter, Inc., CA) was added at 1:1000 dilution in 200 μl per sample of staining/washing buffer (PBS containing 1% saponin and 20% FCS), and the cells were incubated for 20 min at room temperature. After incubation, the cells were washed twice with staining/washing buffer, followed by washing with FACS buffer. Cells were then evaluated on an LSRII and analyzed with the Flowjo Program.
Measurement of HIV-1-specific antibodies
HIV-1-specific human IgM and human IgG in the mouse plasma were measured by ELISA. Briefly, a Maxisorp ELISA plate (NUNC, Rochester, NY) was coated with 5 μg/ml of HIV-1 proteins, recombinant HIV-1 IIIB p55Gag (NIH AIDS Research and Reference Reagent Program, Catalog number 3276), or HIV-1 CN54 gp120 (NIH AIDS Research and Reference Reagent Program, Catalog number 7749) 27,28 in 0.1 M NaHCO3 buffer. After washing and blocking the plates, diluted plasma samples (1:5) were added and incubated for 1 h at 37°C. Either horseradish peroxidase-conjugated antihuman IgG or IgM monoclonal antibodies (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA) were added and incubated for 1 h at 37°C. Substrate (o-phenylenediamine dihydrochloride, Sigma-Aldrich, St. Louis, MO) was added and incubated at room temperature for 30 min. The reaction was stopped with 4 N sulfuric acid and specific absorbance at 490 nm was determined using a microplate reader. Plasma from an HIV-infected human patient was used as a positive control and plasma from hu-DKO mice infected with inactivated HIV, HIV-uninfected hu-DKO mice, and HIV-inoculated DKO mice were used as negative controls.
Administration of HAART to the mice
Mice were treated with a well-described clinical triple combination regimen of two reverse transcriptase inhibitors, AZT and lamivudine, and a protease inhibitor, indinavir, by administering the drugs (45 mg/kg/day for each drug) dissolved in the sterile drinking water at a concentration that provides therapeutic levels of the drugs. 29 Antiretroviral therapy was initiated 1 week after infection with HIV. Seven weeks later the mice were sacrificed and analyzed for the level of HIV infection, human CD4+ and CD8+ T cell subsets, and viral load as described above.
Statistical analysis of data
SPSS version 13 for Windows XP statistical software package was used for the statistical analysis with Pearson correlation and Student's t-test. Differences were considered statistically significant for p-values < 0.05.
Results
Reconstitution of human lymphoid systems in hu-DKO mice transplanted with enriched CD34+ cells and unfractionated human fetal liver cells
DKO mice were sublethally irradiated and injected intrahepatically with human hematopoietic stem cells within 7 days of birth. By 8 weeks after transplantation, human leukocytes were detected in the peripheral blood of almost all of the transplanted mice. Representative FACS staining profiles of mouse bone marrow, spleens, and thymus at 8 weeks after transplantation, stained with antihuman mAbs, are shown in Fig. 1. The bone marrow of the mice contained a large population of human CD19+ B cells as well as a defined population of human CD34+ cells (Fig. 1A). The mice exhibited active human thymopoiesis in the mouse thymus as evidenced by the presence of human double-positive CD4+CD8+ and single-positive CD4 and CD8 human T cells (Fig. 1B). A discrete population of human cells stained by CD45 antihuman mAb was also observed in the spleen of the transplanted mice (Fig. 1C). Of 68 mice transplanted with human fetal liver cells intrahepatically within 7 days of birth, 97% of the animals achieved reconstitution with human CD45+ cells (>1%) at 7 weeks or later after transplantation (mean ± standard error = 14.21% ± 1.56%). There was no significant difference in the level of engraftment of the mice transplanted with highly purified CD34+ cells isolated from fetal liver or with unfractionated fetal liver mononuclear cells (Table 1).

Population of hu-DKO mice with human immune cells. Newborn DKO mice (1–7 days old) were irradiated (3.50 Gy) and injected intrahepatically with enriched CD34+ or unfractionated human fetal liver cells. Representative flow cytometric analysis profiles are shown. (
Within 7 days of birth, sublethally irradiated newborn DKO mice were transplanted by intrahepatic injection with either unfractionated mononuclear human fetal liver cells or highly purified CD34+ cells isolated from human fetal liver. About 2 months later, hu-DKO mice were infected by intrasplenic injection with HIV-1. The mean population of the peripheral blood by human CD45+ cells and the fraction of those cells that were either CD4+ T cells or CD8+ T cells before and 8 weeks after infection and the mean levels of HIV infection measured as TCID50 for the indicated groups of mice are shown. Data are presented as mean ± standard error for each group.
Productive HIV infection of hu-DKO mice
Hu-DKO mice reconstituted with human lymphoid tissues were inoculated by direct injection of HIV-1JR-CSF virus (800 TCID50) into the spleen. As a negative control, three groups of control mice were also included, humanized mice receiving no HIV, humanized mice injected intrasplenically with heat-inactivated HIV, and control DKO mice injected intrasplenically with HIV.
To quantify the level of productive infection in the HIV challenged mice, splenocytes were harvested and cocultured by limiting dilution with PHA/IL-2-activated HIV-naive donor human PBMCs at 8 weeks postinfection. The supernatant was collected 7 days later, and the levels of HIV p24 antigen were measured by ELISA. Of 25 humanized mice injected with HIV, 17 mice became productively infected as indicated by positive coculture with a mean TCID50 value of 1263 ± 377 infected cells per 106 splenocytes (Fig. 2). No productive HIV-1 infection was detected in any mice from the three control groups.
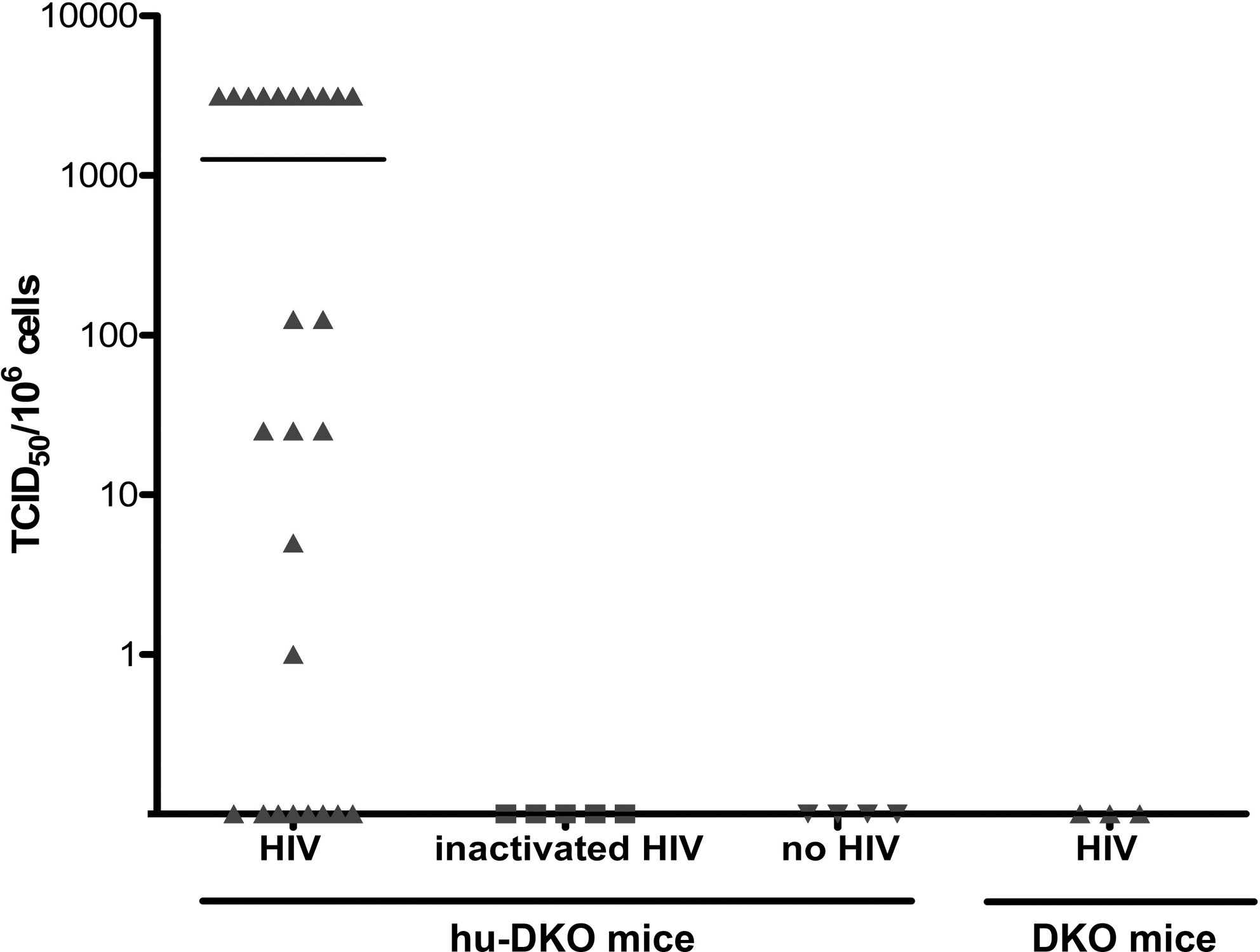
Productive HIV infection of hu-DKO mice. Eight weeks after intrasplenic HIV-1 injection of hu-DKO mice (n = 25 mice), limiting dilution coculture of splenocytes with activated HIV-naive donor human PBMCs was performed. One week later, the p24 antigen levels in the culture supernatant were measured and TCID50 values were calculated. The negative control groups consisted of hu-DKO mice injected with inactivated HIV (n = 5), hu-DKO mice not injected with HIV (n = 4), and DKO mice injected with HIV (n = 3). Data are presented as TCID50 per 1 million spleen cells. Each dot represents one mouse. The mean level is shown as a solid horizontal line.
We further evaluated productive HIV infection in the hu-DKO mice by quantifying plasma levels of HIV-1 mRNA using the branched DNA (bDNA) assay, an FDA-approved signal amplification nucleic acid probe assay that is widely used for the measurement of HIV viral load in humans. 30,31 This assay requires at least 1 ml of a plasma sample for each reaction. However, the amount of plasma available from individual mouse is extremely limited. To overcome this limitation, we pooled the samples from mice in the same experimental group. Although no HIV RNA was detected in the plasma of control mice, the plasma from the humanized mice injected with HIV had a viral load of 47,950 copies/ml. The extent of HIV-1 infection measured by TCID50 was independent of the numbers of human CD45+ cells in the mouse peripheral blood before the infection. The level of productive HIV infection did not differ significantly whether the hu-DKO were reconstituted with unfractionated mononuclear fetal liver cells or with highly purified CD34+ cells isolated from fetal liver (Table 1).
Productive HIV-1 infection in hu-DKO mice depletes peripheral CD4 T cells
A common attribute of acute phase HIV infection in humans is a decline in CD4+ T cell counts and an increase in CD8+ T cell levels in peripheral blood. 32 –34 To determine if this sequela is recapitulated in hu-DKO mice, the CD4 and CD8 lymphocyte population in the PB of the infected mice was analyzed pre- and postinfection. As shown in Fig. 3A, 17 hu-DKO mice productively infected with HIV showed a significant reduction (p = 0.001) of almost 50% in the peripheral blood CD4+ T cell population at 8 weeks postinfection. This was associated with a significant reciprocal almost 2-fold increase (p = 0.009) in the CD8+ levels in mice productively infected with HIV-1 (Fig. 3B). No significant changes occurred during this time period in the human CD4 and CD8 population of the control mice (Fig. 3A and B).
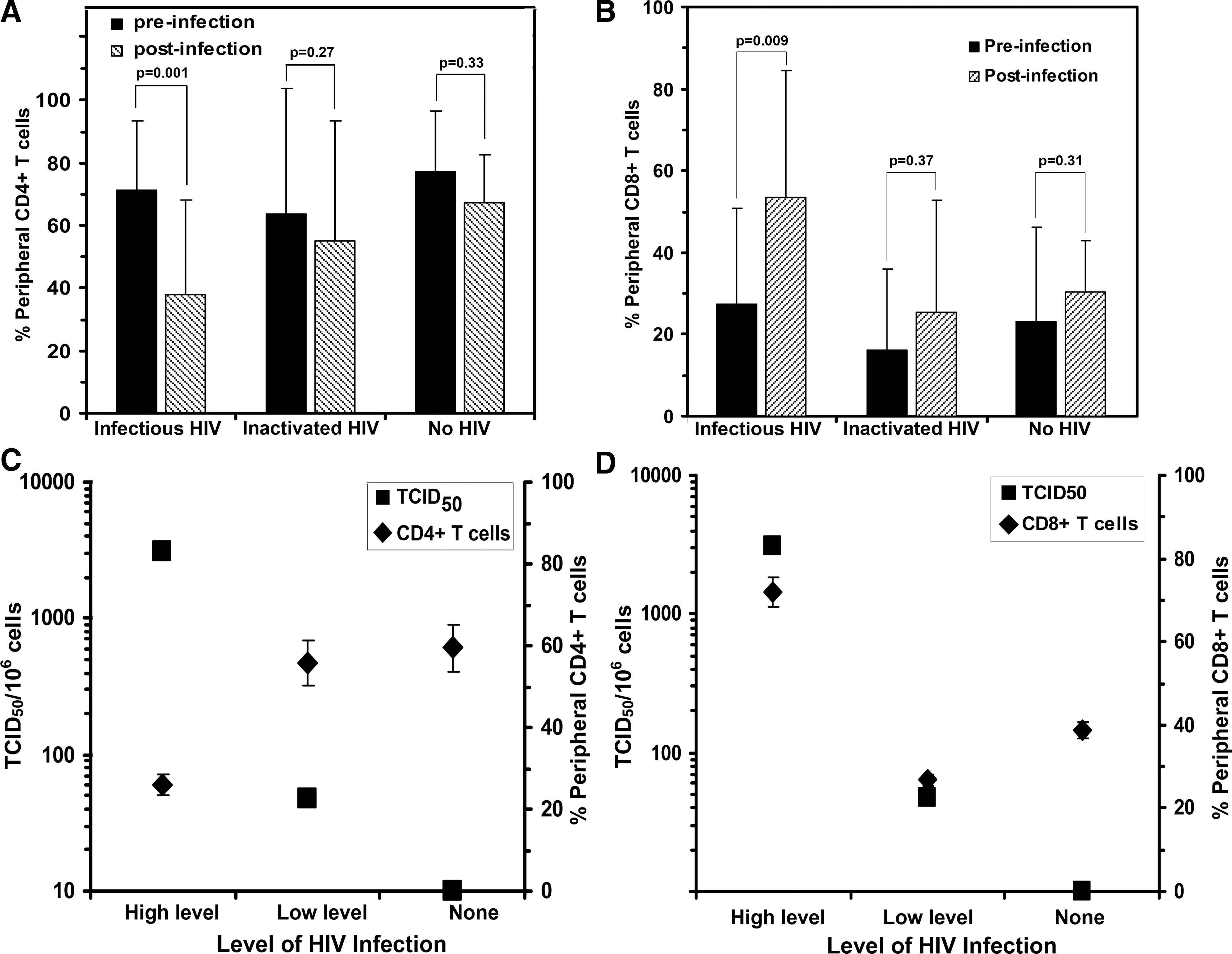
HIV-infection of hu-DKO mice has reciprocal effects on the levels of peripheral human CD4+ and CD8+ T cell subsets. Levels of (
There was an inverse correlation between the degree of CD4+ T cell depletion and the level of HIV infection (Fig. 3C). Significantly more (p = 0.003) CD4+ T cell depletion occurred in mice whose spleens had the highest number of HIV-infected cells. In parallel, there was a significant correlation (p < 0.01) between a high level of HIV infection in mouse spleens and an increase in the CD8+ T cell population after infection (Fig. 3D). These results demonstrate that the productive HIV infection in hu-DKO mice is associated with CD4+ T cell depletion and a reciprocal increase in CD8+ T cells in the peripheral blood that recapitulates the characteristic T cell population changes observed in HIV-infected humans.
To determine the effect of HIV infection on lymphoid tissues, the human thymocytes in the mouse thymus were analyzed 8 weeks after HIV injection, and the fractions of single positive CD4+, single positive CD8+, and double positive CD4+CD8+ human T cells were assessed in the mouse thymus. The fraction of single positive CD4+ cells in the thymuses of the productively infected hu-DKO mice (31.98% ± 5.20, n = 17) was significantly lower (p = 0.01) than the single positive CD4+ population in the uninfected hu-DKO mice (62.49% ± 12.54, n = 5) and markedly lower than the single positive CD4+ thymocyte population in hu-DKO mice injected with inactivated HIV (44.44% ± 8.21, n = 5). In contrast, the thymuses of the productively infected hu-DKO mice displayed an increased fraction of single positive CD8+ thymocytes (33.42% ± 6.82) and double positive CD4+CD8+ thymocytes (39.21% ± 7.25) as compared to the thymuses of the uninfected hu-DKO mice (13.05% ± 5.34) and (24.46 ± 15.20), respectively, and the hu-DKO mice injected with inactivated HIV (18.95% ± 2.64) and (36.61% ± 9.65), respectively.
IgG plasma IgM humoral responses against HIV-1 in hu-DKO mice
While the presence of human immunoglobulin in the plasma of hu-DKO mice has been reported, 17,19,23 reports of HIV-1-specific antibodies present in the serum of these mice after HIV infection in this model are limited. 20 To evaluate the capacity of hu-DKO mice to produce human immunoglobulins and HIV-specific antibodies after infection, human IgG and IgM in the mouse plasma were measured by ELISA. The plasma of HIV-infected humanized mice contained higher levels of both human IgG and human IgM as compared to uninfected humanized mice (Fig. 4A and B). In addition, IgG specific for gp120 was detected in the plasma of the hu-DKO mice inoculated with HIV (Fig. 4C), but not in uninjected hu-DKO mice, hu-DKO mice injected with inactivated HIV, or control mice injected with HIV. The presence of plasma IgG specific for gp120 correlated significantly (p < 0.001) with the levels of HIV-1 infection in the spleen (Fig. 4D), as mice with high levels of HIV infection contained the highest level of gp120-specific IgG.
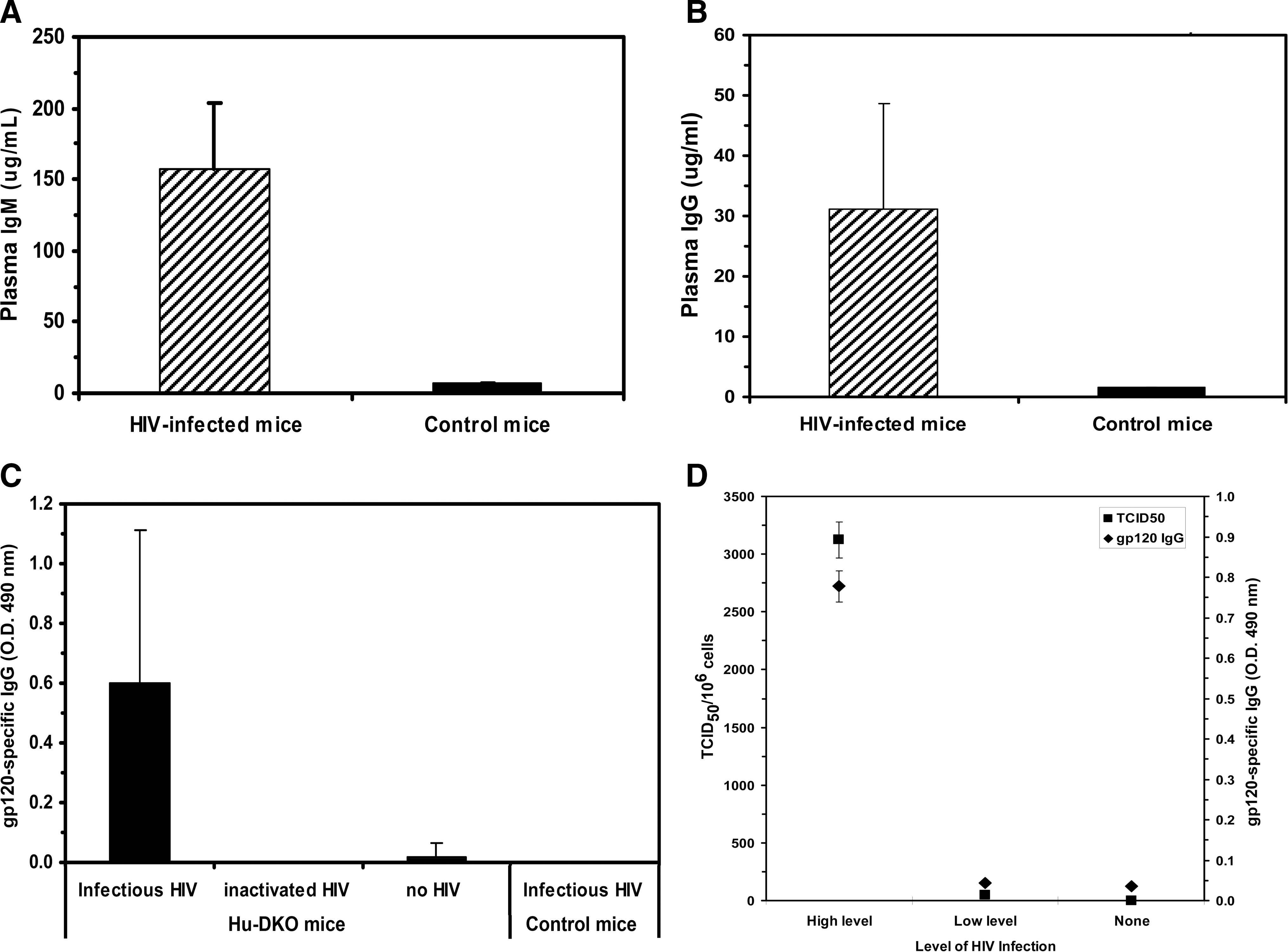
Measurement of total human IgM and IgG levels and HIV-1-specific IgG production in the peripheral blood of HIV-infected hu-DKO mice. (
Hu-DKO mice infected by the intrasplenic route develop increased levels of HIV infection and HIV-specific antibody production than HU-DKO mice infected by the intraperitoneal or intravenous route
To investigate the effect that the route of infection of hu-DKO mice has on the resultant level of HIV infection and HIV-specific humoral responses, we compared the degree of HIV infection and humoral responses in hu-DKO mice injected with the same dose of HIV viruses (8000 TCID50) either intrasplenically, intravenously, or intraperitoneally.
HIV infection in the hu-DKO mouse spleens was evaluated by flow cytometry for intracellular p24 antigen staining. As shown in Fig. 5A, the fraction of p24 antigen-positive cells in the spleens of hu-DKO mice inoculated intrasplenically was about 2-fold and 18-fold higher than splenocytes of hu-DKO mice inoculated intravenously or intraperitoneally, respectively. Flow cytometric analysis indicated that the majority of mononuclear cells containing HIV p24 antigen were human CD45+CD4− lymphocytes. Since these were also CD8− (data not shown) it is likely that these represent CD4+ T cells whose surface expression of CD4 was reduced by HIV infection, particularly by the effect of Nef. 35

Detection of HIV p24 antigen in the spleen cells of hu-DKO mice injected with HIVJR-CSF. (
We further investigated the extent of HIV infection by quantifying the number of cells in the mouse spleen capable of initiating productive infection by limiting-dilution coculture. Eight weeks after inoculation, while all of the HIV-injected groups were infected, the number of HIV-infected cells in hu-DKO mice intrasplenically injected with HIV was 12-fold and 7-fold higher than hu-DKO mice inoculated intravenously or intraperitoneally, respectively (Fig. 5B).
The increased level of HIV infection observed in the intrasplenically inoculated hu-DKO mice was paralleled by increased production of HIV-specific antibodies (Fig. 6A). The level of gp120-specific IgM in the plasma of intrasplenically inoculated hu-DKO mice was nearly 20-fold or 10-fold higher than that of hu-DKO mice infected intravenously or intraperitoneally, respectively; the plasma of hu-DKO mice infected intrasplenically had approximately 5-fold or 10-fold higher levels of gp120-specific IgG than did hu-DKO mice infected intravenously or intraperitoneally, respectively (Fig. 6A). Gag-specific IgM levels were markedly higher in the plasma of intrasplenically infected hu-DKO mice as compared to intravenously or intraperitoneally infected hu-DKO mice (Fig. 6B). The production of IgG against Gag was barely detectible in intrasplenically infected hu-DKO mice (Fig. 6B). There was a statistically significant positive correlation between the level of IgM production against gp120 and the level of HIV infection, as well as the level of IgG against Gag and HIV infection (data not shown).
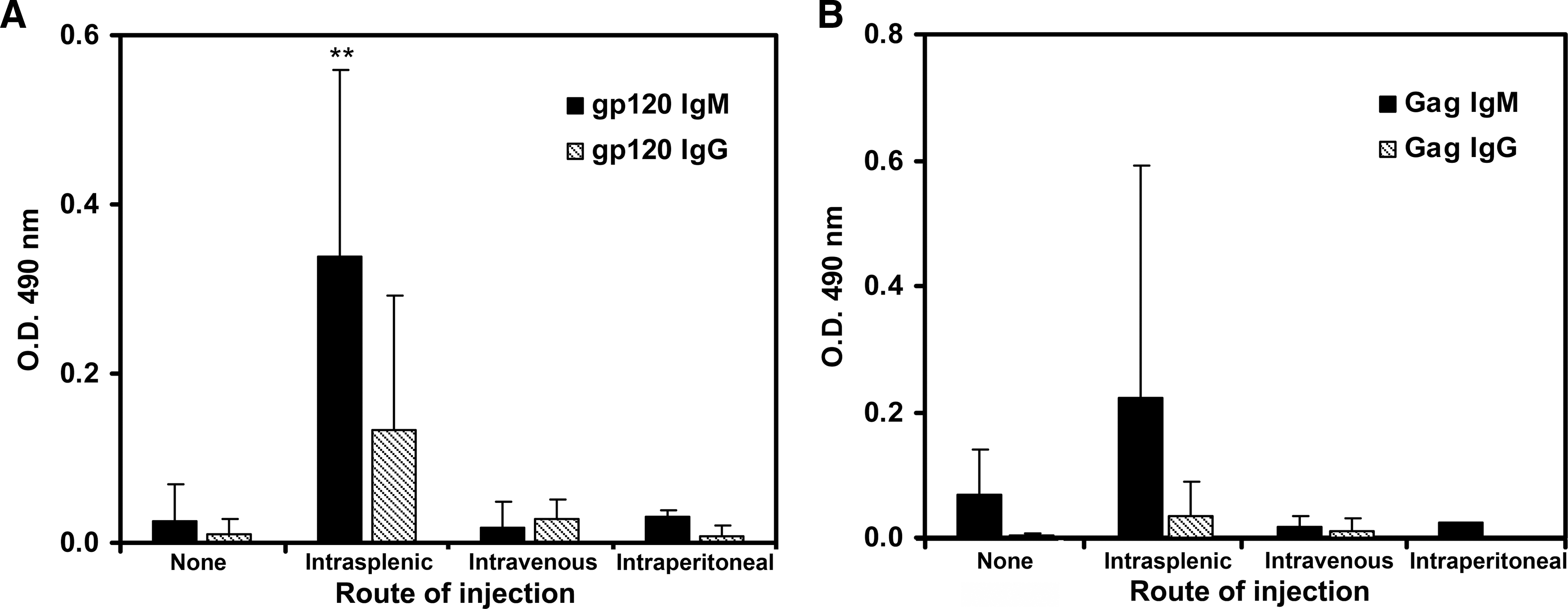
Comparative analysis of HIV-1-specific IgM and IgG production against gp120 and Gag in HIV-injected hu-DKO mice by different routes of injection. At 8 weeks postinfection, HIV-1-specific IgM (solid bars) and IgG (diagonal lined bars) production was determined by ELISA in the plasma of hu-DKO mice injected with HIVJR-CSF intrasplenically (n = 6), intravenously (n = 2), or intraperitoneally (n = 2), and hu-DKO mice not injected with HIV (none, n = 4). Data presented are mean specific absorbance at 490 nm ± standard errors of each group for the indicated ELISA assay. (
Evaluation of the peripheral blood of HIV-infected individuals has indicated that T cells are activated by HIV infection as evidenced by their increased expression of the CD69 activation marker. 36 –38 We examined whether a similar response was induced by HIV infection in hu-DKO mice by analyzing CD69 expression by peripheral human T cells after HIV infection. Although few CD69+ T cells were detected in the peripheral blood of uninfected hu-DKO mice or intraperitoneally infected hu-DKO mice, over 10% of the peripheral T cells expressed CD69 in hu-DKO mice infected intrasplenically with HIV, which was almost 3-fold higher than the fraction of CD69 expression observed in mice infected intravenously with HIV (Fig. 7). Taken together, these results demonstrated that intrasplenic injection of HIV viruses led to more potent HIV infection in hu-DKO mice than intravenous or intraperitoneal injection, which was associated with increased T cell activation and HIV-specific humoral responses.
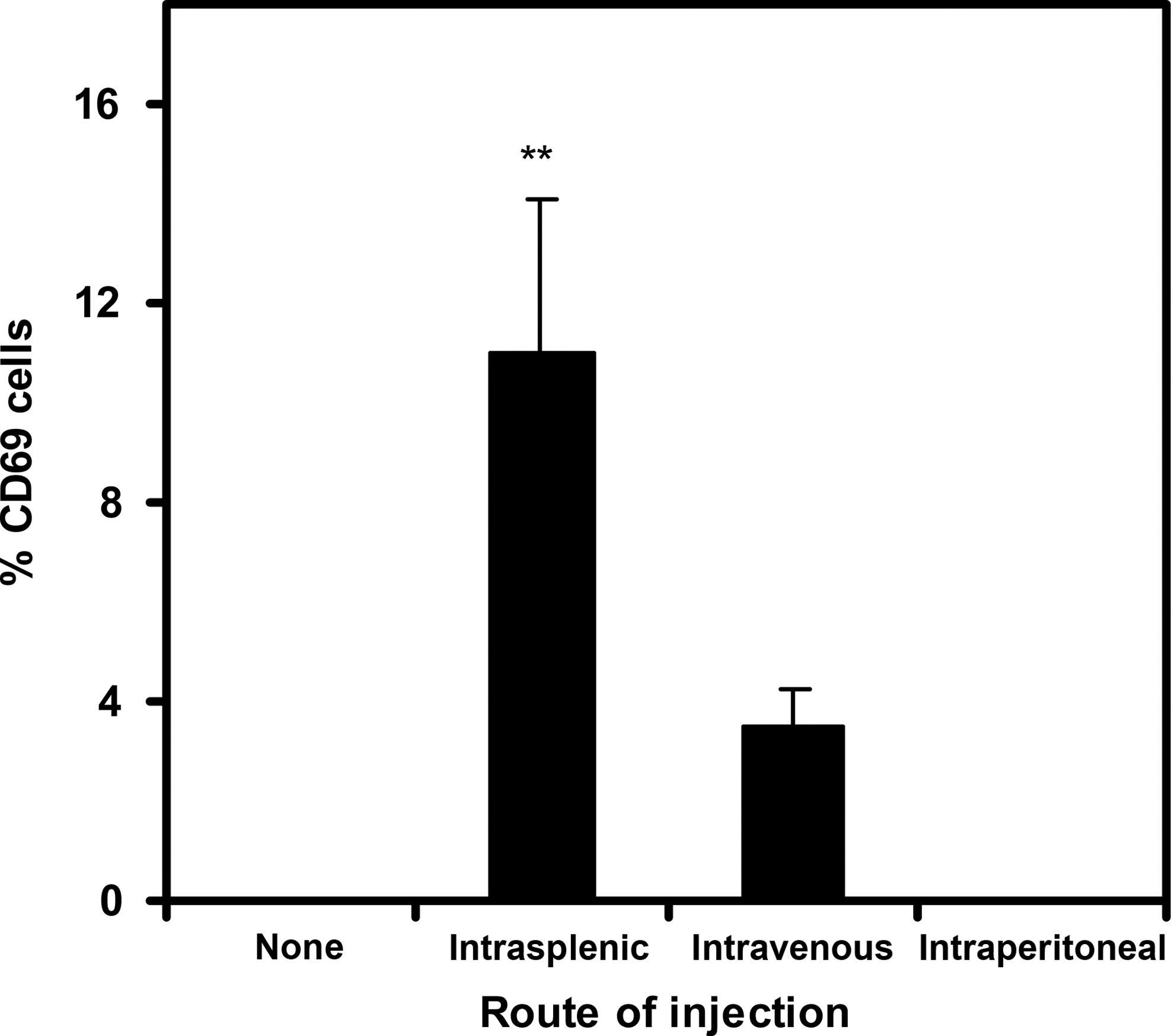
Activation of T cells in the peripheral blood of HIV-challenged hu-DKO mice. At 6 weeks postinfection, the human T cell population in the peripheral blood was assessed by flow cytometry for the expression of the activation marker, CD69, in hu-DKO mice injected with no HIV (none; n = 4), and in hu-DKO mice injected with HIVJR-CSF intrasplenically (n = 5), intravenously (n = 3), or intraperitoneally (n = 3). Data presented are means ± standard errors. **Denotes statistically significant differences between intrasplenic and no HIV (p = 0.03), intrasplenic and intraperitoneal (p = 0.049), intravenous and no HIV (p < 0.01), and intravenous and intraperitoneal (p < 0.01).
Antiretroviral therapy suppresses HIV-1 infection and prevents CD4 depletion in hu-DKO mice
Multidrug therapy combinations such as azidothymidine (AZT), lamivudine, and indinavir have been well established and termed HAART; HAART effectively induces a marked decline in plasma viremia and an increase in the peripheral CD4+ T cell population. 39,40 To determine the utility of using hu-DKO mice as a model to investigate the effectiveness of antiretroviral therapy, we examined the capacity of HAART to inhibit HIV infection. One week after intrasplenic inoculation with HIV-1JR-CSF virus (3600 TCID50), mice were treated with an antiretroviral cocktail consisting of AZT, lamivudine, and indinavir in the drinking water. 29 After 7 weeks of treatment, the mice were sacrificed to evaluate the development of productive HIV infection.
All of the untreated hu-DKO mice (n = 4) were extensively infected with HIV, to the maximum measurable TCID50 value of 3150 HIV-infected cells/106 splenocytes. In contrast, treatment of the HIV-infected hu-DKO mice with HAART (n = 4) resulted in an almost 300-fold (p < 0.001) lower number of HIV-infected cells in the mouse spleen (Fig. 8A). The potent inhibition of infection in the spleen cells of HAART-treated hu-DKO mice was associated with markedly lower plasma viral loads with an over 300-fold reduction in plasma HIV-1 RNA levels (69 copies/ml) (p < 0.001) compared to plasma HIV RNA levels in the untreated group (24,793 copies/ml) (Fig. 8B). HAART-mediated inhibition of HIV infection correlated with preservation of the peripheral CD4+ T cell population. The postinfection level of CD4+ T cells in the peripheral blood of HAART-treated hu-DKO mice was almost unchanged, compared to a nearly 40% reduction in the CD4+ T cell population of the untreated hu-DKO mice (Fig. 8C). This indicates that CD4 depletion caused by HIV infection was prevented by HAART.
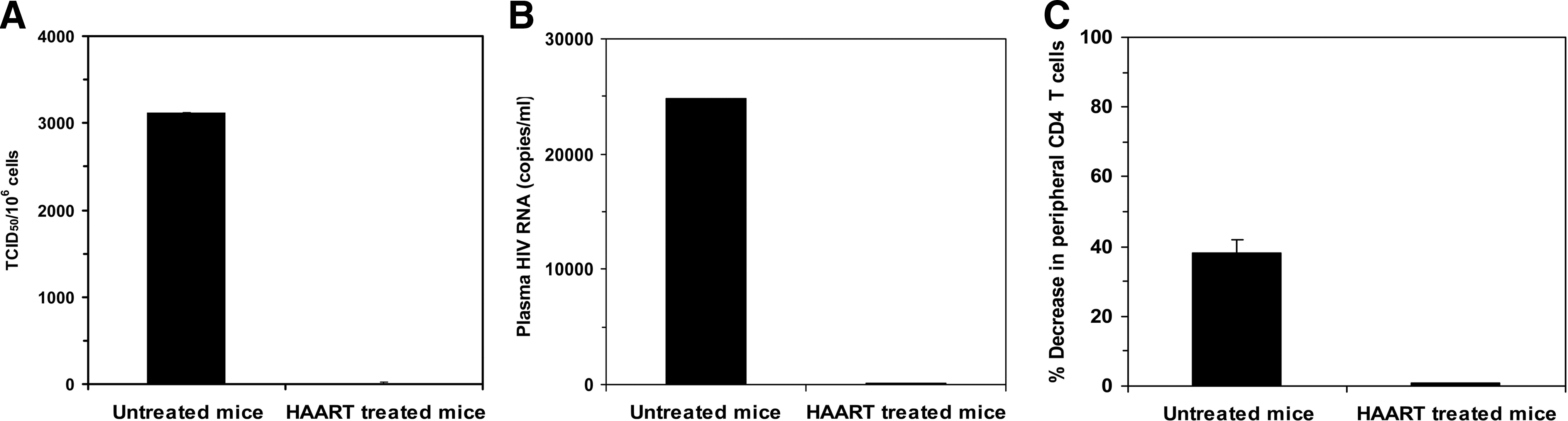
HAART suppresses HIV infection and prevents CD4 depletion in hu-DKO mice. One week after intrasplenic HIV injection, HAART consisting of AZT, lamivudine, and indinavir was administered in the drinking water to hu-DKO mice for 7 weeks. (
Discussion
Humanized mice generated by reconstituting DKO mice with HSCs are an emerging mouse model for studying HIV infection. 18 The severe immunodeficiencies of the DKO mice permit these mice to be extremely tolerant of human HSC transplantation 16,17 and support the subsequent maturation by the transplanted human HSCs into a large repertoire of human immune cells including T cells, B cells, monocytes, and dendritic cells. 18,41 Reconstitution of a functional human immune system in this model provides an experimental system that may be employed to evaluate vaccine and therapeutic strategies against HIV infection.
Varying levels of HIV infection have been reported in hu-DKO mice using different doses of HIV virus injected either intraperitoneally 19,20,22,23 or intravenously 24 that were associated with minimal HIV-specific humoral responses. Baenziger et al. demonstrated in their study that only 1 out of 25 mice produced IgG, but no IgM, against HIV-1 p34, gp41, p52, p58, and gp120. 20 An et al. did not detect HIV-1 specific IgG and IgM against viral proteins prepared from virions or recombinant pr55 Gag 19 while Gorantla et al. failed to detect any HIV-specific antibodies. 23
We hypothesized that using an alternative approach to infecting the humanized mice, directly introducing HIV into the spleen, which is a lymphoid tissue populated with human CD4+ and CD8+ T cells, monocytes, and dendritic cells, could lead to more potent infection and elicitation of HIV-1-specific humoral responses. Using this approach, we detected plasma human IgG against gp120 following HIV-1 infection. The successful induction of humoral responses may be due in part to the direct inoculation of HIV virus in the spleens of hu-DKO mice. The route of antigen administration contributes to the variability of immune responses in HIV-1 infection. 2 Mucosal or lymph node inoculation of SIV antigens was more effective for the induction of immune responses than peripheral infection. 42,43 Intrasplenic inoculation as a route to initiate HIV infection has been demonstrated by us using thy/liv-SCID-hu mice 44,45 and hu-PBL/SPL-SCID mice 46 and by others in hu-PBL-SCID mice. 47 In the hu-PBL NOD/SCID mouse model, intrasplenic injection with HIV-infected human PBMCs induced HIV-specific antibody production. 48 Although the production of human immunoglobulins in hu-DKO mice has been reported, 17,19,20,23 our study is the first report of the production of anti-HIV human immunoglobulins after intrasplenic inoculation in this model.
In accordance with several recent studies, 19,20,22 –24 we achieved successful engraftment of human T, B, and myeloid cells in primary and secondary lymphoid tissues in human HSC-transplanted DKO mice and HIV infection after intrasplenic inoculation of the humanized mice with HIV-1JR-CSF virus. We demonstrated productive infection by quantifying the number of cells in the spleen capable of initiating productive in vitro HIV infection. As reported in other studies of HIV infection in hu-DKO mice, the productive HIV-1 infection was associated with peripheral CD4+ T cell depletion, an increase in CD8+ T cells, and plasma viremia. We observed a significant inverse relationship between the level of HIV infection and the degree of CD4+ T cell depletion and a reciprocal direct correlation with the level of infection and the increase in the level of CD8+ T cells.
HIV challenged mice were monitored for total human immunoglobulin levels in peripheral blood. Plasma human IgG and IgM was detected in all hu-DKO mice, with higher levels of plasma IgM than plasma IgG in all of the hu-DKO mice. Plasma human IgG levels were comparable to that detected with hu-DKO mice immunized with tetanus toxoid (TT), 17 and plasma human IgG and IgM levels are comparable to those described by Watanabe et al. in the HIV-1-infected NOD/SCID/γc -/- model. 49 Recent studies of HIV-1 infection in hu-DKO mice demonstrated either no production of HIV-1-specific human immunoglobulins 19,23 or minimal production. 20 We detected measurable levels of human IgG gp120 in mouse plasma. We speculate that the direct inoculation of HIV-1 virus into spleens that contain large populations of human dendritic cells may promote better antigen presentation and thereby induce immune responses, as compared to other routes of inoculation used including intraperitoneal and intravenous injection. This was supported by our comparative analysis of HIV infection and immune responses after intrasplenic, intravenous, or intraperitoneal inoculation of hu-DKO mice with HIV; intrasplenic injection of HIV resulted in significantly higher levels of HIV infection than other injection routes and induced the most potent HIV-specific IgG and IgM antibody responses against two HIV proteins, gp120 and Gag. There was a high degree of correlation between the level of plasma anti-gp120 IgG and the level of HIV infection in the mouse spleens as measured by TCID50; mice exhibited a higher level of HIV infection mounting a more potent HIV-1-specific antibody responses. Taken together, these results indicate that functional B cells derived from transferred HSCs develop and are able to produce human immunoglobulins including antigen-specific antibodies, suggesting that effective antigen presentation and recognition can occur in the reconstituted immune tissues of hu-DKO mice.
In addition to evaluating the humoral responses, we also examined the development of cellular immune responses against HIV-1 infection using an interferon (IFN)-γ ELISPOT assay. Unfortunately, as reported by others, 19,49 after in vitro restimulation of T cells with HIV-1 antigen we did not detect antigen-specific T cell responses. It is unclear why T cell immune responses do not develop in hu-DKO mice despite their population with human T cells expressing a broad TCR repertoire in the reconstituted mice. 16,17,23 This may be due partially to impaired thymic selection for HIV-specific human T cells, impaired development of the lymph node microenvironment, 18 or lack of human cytokines and chemokines required for appropriate T cell expansion. 41 The lack of HIV-specific CD8+ T cell responses may also be due to inefficient T cell activation in mouse lymphoid tissues or by HIV-mediated reduction of the population of human CD4+ T cells that is required for the expansion of the HIV-specific CD8+ T cell population. Further improvements of the model need to be made, including developing approaches to generate higher and consistent engraftment of the mice with human cells, and prolonged and higher levels of HIV infection, to induce more robust humoral responses and cellular immune responses.
In the present work, we have also demonstrated evidence of the efficacy of antiretroviral therapy in this mouse model. The mice treated with a well-established clinical antiretroviral regimen consisting of AZT, lamivudine, and indinavir showed a significant suppression of HIV infection when compared to the untreated animals, as measured by the decreased number of infectious cells and plasma viremia. We did not observe an occurrence of virus rebound since this established treatment completely suppressed the virus infection. Our observation is in contrast with the recent demonstration of viral rebound and the emergence of antiretroviral drug-resistant mutants in hu-DKO mice by Choudhary et al. 50 We speculate that the more potent treatment regimen we used prevented the emergence of resistant isolates. HAART also prevented HIV-induced reduction of the CD4+ T cell population when compared to HAART untreated mice. These results suggest that the hu-DKO mice may recapitulate the effects of antiretroviral therapy seen in humans.
Thus we have successfully demonstrated the production of HIV-specific humoral responses induced by intrasplenic immunization and the efficacy of antiretroviral therapy in hu-DKO mice reconstituted with a human immune system. Our results further support the application of this model to serve as a valuable in vivo tool to study the pathogenesis, immunology, and virology of HIV infection, and to promote the development of a therapeutic design against HIV infection.
Footnotes
Acknowledgments
This work was supported by the National Institutes of Health (National Institute of Allergy and Infectious Diseases AI67136 and the Einstein-Montefiore Center for AIDS Research AI51519). H. Goldstein is the Charles Michael Chair in Autoimmune Disease. The following reagents were obtained through the NIH AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: HIV-1IIIB p55 Gag and HIV-1 CN54 gp120 from DAIDS, NIAID.
Author Disclosure Statement
No competing financial interests exist.