
Abstract
HIV-1 clade C is the major subtype circulating in India and preferentially uses CCR5 during the entire disease course. We have recently shown that env clones from an Indian patient; NARI-VB105 uses multiple coreceptors for entry and was presented with an unusual V3 loop sequence giving rise to high net V3 loop positive charges. Here we show that env clones belonging to subtype C obtained from an AIDS patient, NARI-VB52, use CXCR6 and CCR8 in addition to CCR5 for entry. However, unlike the NARI-105 patient, the env clones contained a low V3 loop net charge of +3 with a conserved GPGQ motif typical of CCR5 using subtype C strains, indicating that residues outside the V3 loop contributed to extended coreceptor use in this particular patient.
H
Glushakova et al. 20 have shown that there is a strong association between CXCR4 use and cytopathicity in lymphoid tissue; nevertheless, Li et al. 21 provided evidence that at late disease stage in patients harboring R5 variants, the progression of disease is influenced by increased genetic diversity leading to increased replicative capacity. In a typical clinical setting, early in the infection, the HIV-1 envelope exclusively uses CCR5, and with progression toward symptomatic stages preferably tends to switch to exploit CXCR4 or become dual tropic in about 50% of cases at least with subtype B strains. 3,22 The switch is believed to play an important role in the exacerbation and apoptosis of CD4+ T cells, leading to opportunistic infection and eventually death. 17,22 –24 With the switch from CCR5 to CXCR4, with viral isolates changing from non-syncytium using (NSI) to syncytium using (SI), viral variants are believed to exploit a broader range of cell types leading to efficient killing of T cells. 25
In clade C infections, CXCR4-using variants have rarely been detected in individuals in the late stages of disease, 26 –30 though two studies indicated a relatively higher frequency of X4 variants in patients who received highly active antiretroviral therapy (HAART). 31,32 Nevertheless, in HIV-1 clade C infections, disease progression leading to acquired immunodeficiency syndrome (AIDS) and death predominantly occurs by R5 variants.
Isaacman-Beck et al. 33 have recently shown that clade C variants obtained from Zambian patients use alternate coreceptors. In the present study, we investigated the coreceptor binding properties of HIV-1 clade C env clones obtained from a female AIDS patient from Western India with a CD4 count of 51 cells/mm3. The patient had a history of sexually transmitted diseases (STD) and presented with genital ulcer disease, oral candidiasis, and herpes genitalis.
gp160 was PCR amplified from low passaged HIV+ peripheral blood mononuclear cells (PBMCs) using 5′-CAC CGGCTTAGGCATCTCCTATGGCAGGAAGAA-3′ as forward primer
34
and 5′-TAACCCTTCCAGGTACCCCCTTTT CTTTTA-3′ as reverse primer with a proofreading Platinum Taq DNA polymerase (Invitrogen Inc.). A KpnI site was inserted into the reverse primer and the amplicon obtained from maximally diluted HIV+ PBMCs was ligated into pSVIIIenv.
35
Clones with gp160 inserts were screened by restriction digestion to confirm proper orientation. Hypervariable loop sequences of env clones were checked against known and published sequences available in the HIV-1 database (
To determine viral coreceptor use patterns we tested a number of gp120 clones obtained from patient NARI-VB52 in a cell–cell fusion assay. Thus, env clones of NARI-VB52 were tested for their ability to exploit additional coreceptors other than CCR5 by cell–cell fusion assay between env+ producer cells and CD4+ target cells expressing different coreceptors as described previously. 30,35 Briefly, env+ 293T cells were cocultivated with GHOST (4 × 104 cells per well) cells expressing CCR5, CXCR4, CCR2b, CCR3, GPR1, GPR15, CXCR6, CX3CR1, APJ, and CD4+ NP2 cells expressing CCR8 (4 × 104 cells per well) and monitored for induction of multinucleated giant cells or syncytia. Cells were subsequently washed in phosphate-buffered saline (PBS) and fixed in methanol containing 1% methylene blue (to stain cytoplasm) and 0.25% basic fuchsin (to stain nucleus) for 10 min. Cells were washed with PBS multiple times and examined under an inverted microscope to measure syncytia formation. As shown in Fig. 1, NARI-VB52.J9, J19, J29, and J30 were found to use CXCR6 and CCR8 in addition to CCR5.

Cell–cell fusion assay. 293T cells expressing the NARI-VB52.J9 clone were cocultivated with GHOST cells expressing CCR5 and CXCR6 and CD4+ NP2 cells expressing CCR8. Multinucleated giant cells representing syncytia were observed by staining with methylene blue and basic fuchsin as described in the text. Note that similar phenotypes were observed with other NARI-VB52 env clones. In the mock well, untransfected 293T cells were mixed with GHOST-CCR5 cells.
We next tested the infectivity of both the primary isolate and the pseudoviruses obtained from NARI-VB52 (J9, J19, J29, and J30) to infect GHOST cells expressing CCR5, CXCR6, and NP2-CD4-CCR8. Pseudotype viruses carrying the patient envelope were produced by cotransfection of pSVIIIenv carrying different NARI-VB52 env clones and env-defective HIV-1 backbone vector (pSG3ΔEnv) 37 into 293T cells in a six-well tissue culture tray (Corning Inc.) using calcium phosphate (Promega Inc) following the manufacturer's protocol as described previously. 35 Pseudoviruses with equal focus-forming units (FFU) were added at different dilutions to GHOST cells expressing CCR5, CXCR6, and NP2 cells expressing CCR8 and further incubated for 72 h in a humidified CO2 incubator at 37°C. Cells were fixed and permeabilized with chilled methanol and acetone (1:1 v/v), and infectivity was assessed by staining intracellular p24 using anti-p24 Hybridoma supernatant 183-H12-5C as primary antibody as described before. 35
As shown in Fig. 2, both the primary NARI-VB52 isolate and the pseudoviruses made from env clones were found to use CCR5 and CXCR6 maximally with CCR8 utilized least. Additionally, the primary isolate obtained from this patient was found to replicate efficiently in PBMCs in the presence of a saturated concentration of the CCR5 antagonist TAK-779 (data not shown), further substantiating our data. To further indicate that NARI-VB52 used CCR5, CXCR6, and CCR8, we infected PBMCs preincubated with TAK-779 (CCR5 antagonist), vMIP-I (antagonist to CXCR6, CCR8, CCR3, and GPR15), and I-309 (CCR8 antagonist) (R & D systems, Inc.) with the NARI-VB52 primary isolate and tested virus growth by measuring p24 levels. Briefly, 1 × 106 PHA/IL-2-stimulated PBMCs were preincubated with 1000 nM vMIP-I, 38 100 ng/ml of I-309, 39 and 10 μM TAK-779, respectively, for 1 h at 37°C, and subsequently infected with 50,000 relative luminescence units (RLU) relative to TZM-bl cells of the NARI-VB52 primary isolate. Cells were washed after 3 h and the plate returned to a humidified 37°C incubator supplemented with 5% CO2. At days 7 and 14, cell-free viruses were clarified by centrifugation and virus growth was monitored by measuring p24 in an ELISA following the manufacturer's protocol (Vironostica HIV-1 antigen kit, Biomerieux Inc.). As shown in Fig. 3, none of the antagonists was found to inhibit NARI-VB52 replication in PBMCs, suggesting that the virus indeed used multiple coreceptors for entry. The use of CCR8 had not previously been reported with subtype C infection.
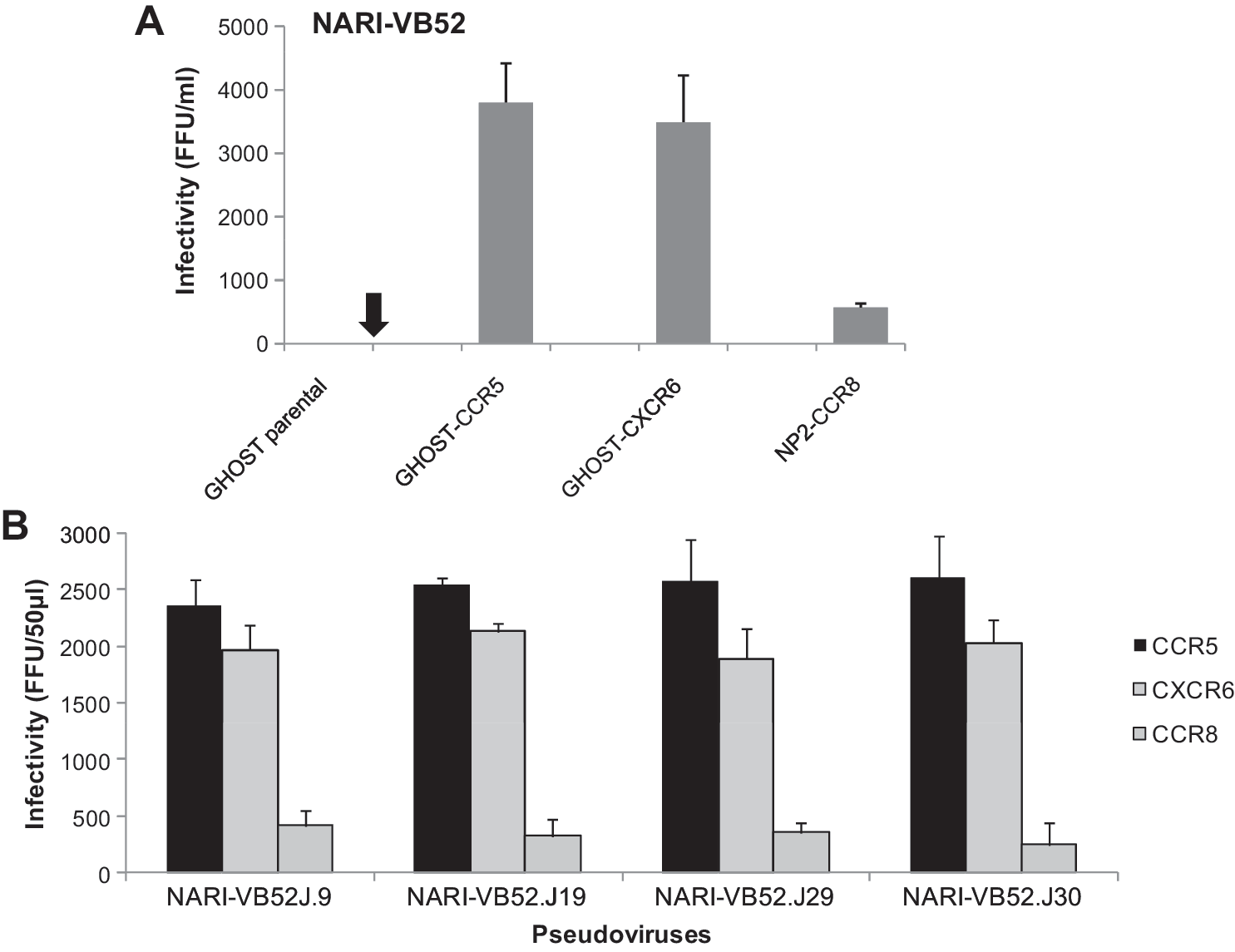
Cell-free infectivity of primary and pseudoviruses bearing different NARI-VB52 envelopes in indicator cell lines. GHOST cells expressing

Growth kinetics of the NARI-VB52 primary isolate. PHA/IL-2-stimulated peripheral blood mononuclear cells (PBMCs) were preincubated with antagonists to CXCR6 (vMIP-I), CCR8 (I-309), and CCR5 (TAK-779) as described for 1 h at 37°C in a humidified chamber supplemented with 5% CO2, and subsequently infected with 50,000 RLU of the NARI-VB52 primary isolate and further incubated for an additional 3 h under similar conditions. PBMCs were then washed and replenished with growth media and incubated further. The kinetics of virus replication was assessed by measuring p24 in cell-free virions.
To investigate the genetic properties of NARI-VB52 env clones, deduced amino acid sequences of all the env clones and the sequence of the consensus PCR product (primary isolate sequence) were aligned using MEGA 4.0.1 and compared with the HIV-1 consensus C sequence (obtained from the HIV Los Alamos database) as well as those of Indian clade C env obtained from patients who were found to be strictly CCR5 using. 30 The sequence of the primary virus was found to be genetically very closely related to all the NARI-VB52 env clones (Fig. 4A). As shown in Fig. 4B, no unusual V3 loop sequence was seen in all these clones, which were presented with the GPGQ motif typical of subtype C carrying a net positive charge of + 3, suggestive of CCR5 use. In addition, no significant differences were observed in the number of potential N-linked glycosylation sites (PNLG) and variable loop lengths between the NARI VB52 env clones and other env clonal sequences that were strictly CCR5 using. Interestingly, compared to the consensus subtype C sequence, NARI-VB52 env clones were found to contain a longer V1 loop as well as additional PNGTKS and EESN sequences in the V4 and V5 loops, respectively. However, since these differences were not found in env clones that were strictly CCR5 using, they may not possibly be influenced by the extended coreceptor tropism. Nonetheless, our data indicate that non-V3 domains most likely confer in expanded coreceptor use by the NARI VB52 env clones.






In summary, we amplified env clones from a terminal patient with AIDS, infected with clade C HIV-1, and studied both coreceptor use and the corresponding env sequences. Earlier we found env clones from a late stage patient (NARI-VB105) with the capacity to exploit several coreceptors, including CCR5 and CXCR4, and that contained an unusually high net positive charge in the V3 loop. 30 However, unlike NARI-VB105 envs, NARI-VB52 env clones were found to contain a low net positive charge and a typical GPGQ motif in the V3 loop despite using alternate coreceptors, CXCR6 and CCR8, indicating that regions outside the V3 loop in envelope possibly have influenced alternate coreceptor use. The PNLGs in envelope have been shown, by different investigators, to modulate tropism. 40 –42
Blay et al. 43 reported that the majority of PNLG changes fall within limited windows, or “hot spots” of the HIV-1 envelope, and they include the N-terminal side of V1, a small region on the C-terminal side of V2, most of V4, and a small region of V5. They concluded that within these “hot spots” variation in the positioning of glycan residues is associated with coreceptor switching and an altered immune response. It was reported that clade B envelopes, in contrast to clade A and C envelopes, do not necessarily accumulate PNLGs over time from infection, whereas clade A and clade C envelopes tend to have a reduced numbers of PNLGs early in infection and gradually acquire more as infection progresses. 44,45
Coetzer et al. 26 recently showed that clade C envelopes obtained from two South African patients who underwent a coreceptor switch showed an increase in the overall length of the V1 loop. In our study, although NARI-VB52 env clones obtained from a late stage patient were found to contain similar loop lengths in gp120, there was only one PNLG site in the V1 loop of NARI-VB-52.J29 and J30 clones in comparison to four PNLGs found in clones J9 and J19. Nonetheless they were all found to use CXCR6 and CCR8 in addition to CCR5, indicating that differences in PNLG patterns in the V1 loop found in NARI-VB52 clones did not influence coreceptor switch.
Though considerable variations in genotype and tropism have been studied with HIV R5 variants belonging to subtype B, 46 –48 not much is known about the relationship between the genetic properties and tropism of subtype C viruses, particularly at the late stage of disease. The complex relationships between factors governing disease progression and onset of AIDS are not clearly understood. Further studies toward understanding the viral determinants influencing the expanded coreceptor tropism in clade C strains modulating the rate of transmission and disease progression will help in providing important insight toward the comprehensive characterization of the replication of HIV-1 subtype C circulating in countries such as India, where CCR5 using variants are predominant.
Nucleotide Sequence Accession Numbers
The GenBank accession numbers for NARI-VB52 env clones are EU760888, EU521727, EU760889, and EU760890, respectively. The sequence of the primary virus (consensus PCR product) is GU945306.
Footnotes
Acknowledgments
We are grateful to Dr. Paul Clapham, UMASS Medical School, Worcester, MA for providing GHOST and NP2 cell lines as well as the pSVIIIenv vector. We thank Dr. Hiroo Hoshino, University of Gunma, Japan for permitting the use of NP2 cells. This work was supported in part by research grants from the Department of Biotechnology, Government of India (BT/PR7829/Med/14/1133/2006 and BT/PR7747/Med/14/1097/2006) to J.B. L.G. and R.R. are supported by Senior and Junior Research Fellowships from the Department of Biotechnology, Government of India, and Indian Council of Medical Research, Government of India, respectively. We gratefully acknowledge the NIH AIDS Research Reagent and Reference Program for making available pSG3Δenv and anti-p24 hybridoma supernatant 183-H12-5C for p24 immunostaining. We thank Dr. R.S. Paranjape, Director, NARI for support.
Author Disclosure Statement
No competing financial interests exist.