
Abstract

E
A cohort of eight CRF07_BC-infected subjects (six men and two women with an average age of 32.9 years), who were defined as virologic noncontrollers (antiretroviral drug-untreated individuals with plasma HIV-1 RNA levels of >2000 copies/ml), 4 participated in this study after they signed an informed consent form. All the subjects were IDUs from Xinjiang Province of China. They had an average viral load of 6.6 × 105 copies of HIV-1 RNA/ml, as measured with HIV-1 nucleotide fluorescence quantitative assay kits (BD Biosciences, Franklin Lakes, NJ). 3 Their average CD4+ and CD8+ T lymphocyte counts analyzed with the FACS/Lyse kits provided by BD Biosciences 5 were 331.6 ± 187 and 1226.5 ± 521 counts/μl, respectively. Their peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Paque gradient (Amersham Biosciences, Uppsala, Sweden) for isolation of primary HIV-1 virions as previously described. 3 Briefly, PBMCs isolated from each of the HIV-1-infected patients were cocultured with phytohemagglutinin (PHA)-stimulated PBMCs from healthy donors. The cell cultures were maintained for 4 weeks in RPMI 1640 medium (Gibco) containing 20 U/ml of recombinant interleukin-2 (IL-2), 1% penicillin and streptomycin (P/S), 2 mM glutamine, and 10% fetal bovine serum (FBS) and the culture media were changed twice a week. The culture supernatants containing HIV-1 virions were collected for detection of p24 content and virus titer, then aliquoted and stored in liquid nitrogen until used.
A sensitive HIV-1 neutralization assay using TZM-b1 cells 6 was performed to measure neutralizing activity of the patient-derived plasmas against autologous virus (that was isolated from the same patient whose plasma was tested) and heterologous viruses (that was isolated from a different patient than the patient whose plasma was tested). The luciferase activity was measured using a luciferase assay kit (Promega, Madison, WI) and a Reporter Luminometer (Perkin-Elmer Life Sciences, Wellesley, MA). The neutralizing activity was recorded as the reciprocal of the dilution of plasma required to confer 50% inhibition (ND50) of infection by a virus.
As shown in Table 1, all the plasma samples were effective in neutralizing heterologous viruses of CRF07_BC with a mean ND50 of 91. However, six of eight plasmas showed no detectable neutralizing antibody responses against autologous viruses, while the remaining two plasmas exhibited marginal neutralizing activity (ND50: 11–13). These results suggest that the majority of the CRF07_BC viruses can induce a neutralizing antibody response but become resistant to autologous neutralization. It is well documented that isolate-specific neutralizing antibodies are rapidly developed after primary HIV-1 infection, but virus variants resisting neutralization by autologous sera emerge subsequently to escape the immune pressure exerted by neutralizing antibodies. 7,8 However, it is puzzling why the neutralizing antibodies are still highly effective against heterologous viruses of the same subtype (CRF07_BC). We speculate that the neutralizing antibody responses against heterologous viruses may protect against HIV-1 reinfection (or superinfection). It was reported that HIV-1 superinfection occurred in acute viral infection, 9,10 but was uncommon in advanced stages of HIV-1 infection. 11
The plasmas were tested at a series of 3-fold dilutions ranging from 1:8 to 1:648. For calculation, the ND50 <8 and >648 were treated as 8 and 648, respectively. The neutralizing titers of plasmas against the autologous virus are marked in
Strikingly, we found that the neutralization sensitivity of HIV-1 CRF07_BC isolates was inversely correlated with the neutralizing activity of the patient-derived plasmas (Fig. 1). The plasmas from some of the patients exhibited a high level of neutralizing activity, whereas the viruses isolated from these patients were highly resistant to neutralization by both heterologous and autologous plasmas. For example, the plasma from patient XJ1981 (XJ1981P) was most effective in neutralizing the heterologous viruses (mean ND50 = 239), whereas the virus from the same patient (XJ1981V) was least sensitive to neutralization by plasmas from HIV-1-infected patients (mean ND50 = 8.9). In contrast, plasma from patient XJ135 (XJ135P) was ineffective in neutralizing any of the viruses tested (ND50: < 8), whereas the virus from this patient (XJ135V) was most sensitive to neutralization by patient-derived plasmas (mean ND50 = 198). It is worthwhile to compare the antigenic structures of the envelope proteins of XJ1981V and XJ135V in order to identify the critical antigenic sites that can induce potent neutralizing antibody responses or those that protect against neutralizing antibodies. It is also interesting to comparatively investigate molecules in XJ1981P and XJ135P to identify the serum factors that may be associated with immune suppression against virus-specific immunity. This study will provide important information for the design of novel anti-HIV vaccines.
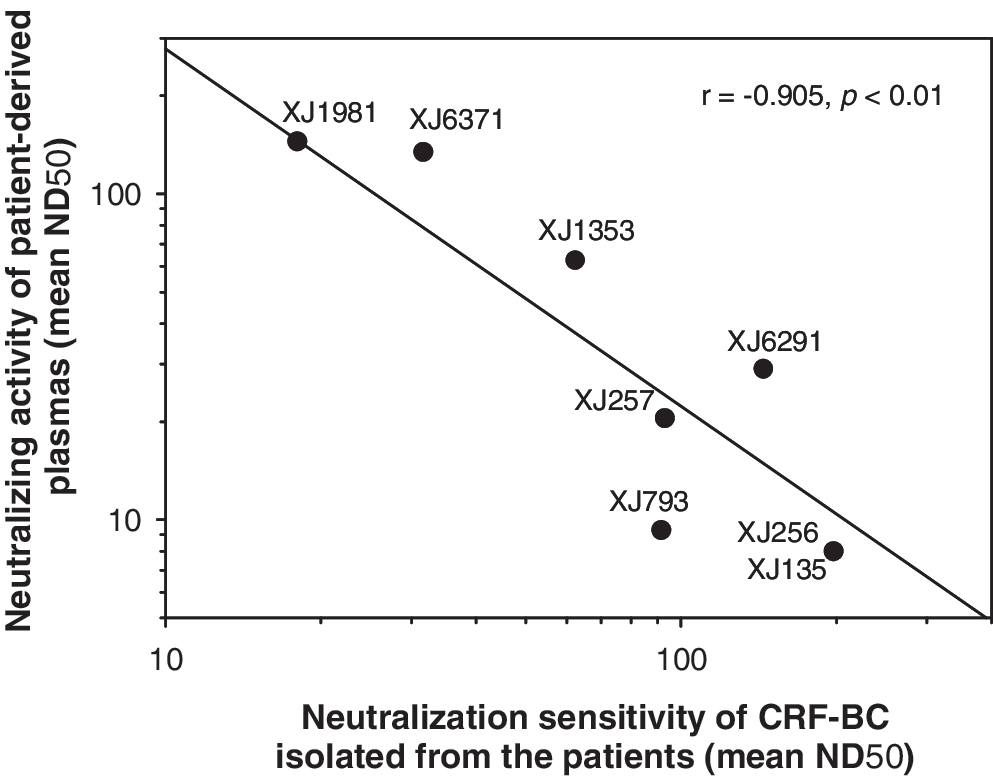
Neutralizing activity of plasmas from HIV-1 CRF07_BC-infected patients was inversely correlated with neutralizing sensitivity of the viruses isolated from these patients.
Footnotes
Acknowledgments
This work was supported by the Ministry of Science and Technology of China (2005CB523103 to L.M., 2007DFC30230 to Y.S.), National Key Science and Technology Special Project (2008ZX10001-010 and 2008ZX10001-013 to Y.S.), State Key Laboratory for Infectious Disease Prevention and Control (2008SKLID101), and National Nature Science Foundation (30872232 to L.M.).
Author Disclosure Statement
No competing financial interests exist.