
Abstract
It is necessary to understand the molecular nature of the virus population that persists in cellular reservoirs. To achieve this we planned to characterize the patterns of resistance of HIV-1 in CD14+ monocytes, CD4+ T cells, and plasma. Blood samples were collected from 42 patients treated for HIV: 32 were in virological failure and in 10 viremia was undetectable. CD14+ and CD4+ T cells were isolated using magnetic beads. Genotyping of the reverse transcriptase and protease gene of HIV-1 was undertaken using the fluorescent dideoxy-terminator method. Of the 32 patients in virological failure, 24 (75%) had resistance mutations in at least one compartment. The numbers and types of mutations from monocytes were the same as those detected in both CD4+ T cell-associated virus and plasma in only 8% whereas in 71% monocytes exhibited a different mutation pattern. In 21% of patients, the profile of drug-resistant mutations in the virus from blood monocytes was identical to that in plasma but differed from that in CD4. In the 71% of patients with virological suppression, the genotypic resistance pattern differed between monocytes and CD4+ T cells. Circulating monocytes may harbor a viral dominant population different from those viruses circulating in blood and archived in CD4+ T cells. Hence, monocytes and other cellular reservoirs might serve as an indirect source of a drug-resistant viral variant.
Introduction
S
Resting memory CD4 T cells have been proposed as the most significant cellular reservoirs for HIV-1, 1 –3,9 but HIV-DNA has also been found to persist in naive CD4 T cells 10 and in cells of monocyte/macrophage lineage, including blood-circulating monocytes. 11 –13 Monocytes are known to be released into the blood from bone marrow and to circulate in peripheral blood for 1–5 days before differentiating into immature dendritic cells and other tissue macrophages. Several studies have shown that HIV-1 can be detected in blood monocytes. 13 –15 However, it is not clear whether the virus is produced or maintained latently in monocytes. Recent studies have shown that blood monocytes contain HIV-1 variants that are genotypically and phenotypically distinguishable from those present in CD4+ T cells. 16,17 –20 Furthermore, it has been reported that in patients receiving suppressive ART, monocytes/macrophages are the major source of plasma HIV-1. 16,17,21 –23 These data suggest that HIV-1 circulating in blood monocytes represents the residual replicating HIV-1 that might escape ART and might be produced from tissue macrophages. The explanation of such viral replication in monocytes is that antiretroviral drugs may not block viral replication in these cells as efficiently as in CD4+ T cells. 24,25 Given the low level of replication in circulating monocytes and the reduced permeability of these cells to drugs, new HIV-1 infections of cells during therapy would be more likely to occur in monocytes rather than in CD4+ T cells. This might, in general, contribute to an increase in HIV diversity.
Within the framework of a study aimed at determining the influence of ART on the appearance of HIV-1 drug-resistant variants in monocytes, the mutational resistance patterns of HIV are described in different compartments of peripheral blood: that is, CD14+ monocytes, CD4+ T cells, and plasma samples from treated patients.
Materials and Methods
Patients
Blood samples were collected from 42 ART-experienced patients after informed consent. All were infected with the HIV-1 subtype B strain and had been receiving ART for a long period (mean, 11 years; range, 3–18 years). In particular, at the time of blood sampling, current ART was failing in 32 patients and viremia levels were undetectable in the remaining 10 patients.
With regard to the therapeutic histories of the 32 patients in virological failure, 11 included three classes of drugs—nucleoside reverse transcriptase inhibitors (NRTIs), non NRTIs (NNRTIs), and protease inhibitors (PIs); eight had received NRTIs and PIs, five were PI-naive but NRTI and NNRTI experienced, and eight had been treated with all three classes of drugs, together with raltegravir and/or enfuvirtide. Of the 10 patients responding to treatment, four had received three classes of drugs, one was NNRTI-naive and had received enfuvirtide, three had been treated with NRTI and PI, and two were PI naive.
At the time of analysis, the median CD4 cell count was 327 cells/μl (range 14–1115 cells/μl) and the median viral load was 27,914 copies/ml (range 800 to >500,000 copies/ml).
Isolation of CD14+ monocytes and CD4+ T lymphocytes
After separation of plasma, peripheral blood mononuclear cells (PBMCs) were isolated from venous blood samples and separated on Lympholyte-H (Cedarlane Laboratories Limited, Canada) gradients. The isolated cells were centrifuged and pelleted, washed twice with phosphate-buffered saline, and then counted in Turk staining. CD14+ monocytes were purified from 30 × 106 PBMCs by positive selection using micromagnetic beads anti-CD14 monoclonal conjugated antibodies (Miltenyi Biotec, Bologna, Italy). CD4+ T lymphocytes were purified from monocyte-depleted cells by positive selection using micromagnetic beads anti-CD4 monoclonal conjugated antibodies (Miltenyi Biotec, Bologna, Italy).
After magnetic separation, cells were checked for purity by fluorescent-activated cell sorting (FACS) analysis, the purity of isolated cells being analyzed by flow cytometry using FACS (FacsCalibur–Becton Dickinson) after the immunofluorescent staining of samples with monoclonal antibody to the following cell surface molecules: CD14, CD4, CD3, and CD19.
HIV genotyping
RNA was extracted from 140 μl of plasma using a QIAmp Viral RNA kit (Qiagen, Milan, Italy) following the manufacturer's instructions. The HIV-1 reverse transcriptase (RT) and protease genes were amplified and sequenced using a TruGene HIV-1 Genotyping kit and an OpenGene automated DNA sequencing system (Siemens Healthcare Diagnostics, Milan, Italy). The results were interpreted using GuideLine Rules 12.0. The entire protease gene (codon 1–99) and codon 37–248 of the HIV-1 RT gene were sequenced. Viral RNA was converted to 1.3 kb cDNA and amplified by polymerase chain reaction (PCR) in a single tube RT-PCR reaction. The reaction products were then added, without purification, to a set of sequencing reaction tubes. Sequencing was undertaken using CLIP, a DNA sequencing technique for direct sequencing of small quantities of amplified template (TruGene HIV-1, Siemens Healthcare Diagnostics).
DNA was extracted from CD14+ monocytes and CD4+ T cells using a Dneasy Tissue kit (Qiagen, Milan, Italy); to check the DNA extraction, the β-actin gene was amplified as described previously. 26 The RT-PCR step provided in the TruGene HIV-1 was modified for viral DNA. Specifically, 2 μl of the extract, containing approximately 0.3 μg of DNA, was added to the PCR reaction. Samples were denatured for 5 min at 94°C, followed by 20 cycles at 94°C for 30 s, 57°C for 30 s, and 68°C for 90 s, then by 17 cycles at 94°C for 30 s, 60°C for 30 s, 70°C for 2 min, with a final incubation at 70°C for 7 min. Following the manufacturer's instructions, the PCR products were sequenced by TruGene HIV-1.
Statistical analysis
To evaluate the association of the number of mutations with a different time of treatment, a Spearman correlation was undertaken. We calculated the proportion of monocytes samples that showed numbers and/or types of drug-resistant mutations different from those detected in either the plasma virus or the CD4+ T cell-associated virus, with the corresponding 95% confidence intervals.
Results
Blood monocytes, CD4+ T cells, and plasma were obtained from 42 samples of PBMCs as described in Materials and Methods. Of these, 32 were from patients failing ART and 10 from virologically suppressed individuals.
The blood monocyte suspensions were 94–96 % pure (with 1–3% CD3+ and 1–2% CD19). CD4+ T cell preparations were 95–99% pure (with 0.7–1% CD14+ and 0.23–1% CD19+) (Fig. 1).
Flow cytometric purity control of CD14+ and CD4− (
The analysis of mutation patterns associated with drug resistance detected in plasma HIV-RNA and in HIV-1 from CD14+ monocytes and from CD4+ T cells revealed that 8 of the 32 samples analyzed showed virological failure (indicated by the rebound of viral load), without showing any resistance mutations in either cellular compartments or plasma. Careful anamnesis by clinicians showed that these patients were not adherent to ART.
In 24 of the 32 patients' samples (75%) drug-resistant mutations were seen in at least one of the compartments analyzed. In two (8.0%) of these 24 samples the mutation pattern of HIV from blood monocytes was identical to those detected in both CD4+ T cells and plasma. In 17 of the 24 samples, the virus found in monocytes showed numbers and/or types of drug-resistant mutations that were different from those detected in either the plasma virus or the CD4+ T cell-associated virus (71%; 95% confidence interval, 53–89). In five patients (21%), the profile of drug-resistant mutations in the virus from blood monocytes was identical to that in plasma, but different from that found in CD4+ T cells (the results are summarized in Table 1).
PL, plasma.
In virological success, patients' data are referred to CD4+ lymphocytes and monocytes.
In 50% of samples, the circulating monocytes appear to harbor a viral population with fewer mutations [mean value (±SD) = 8.3 ± 7.7] than were found in plasma [mean value (±SD) = 14.7 ± 7.4] and in CD4+ T cells [mean value (±SD) = 13.7 ± 7.6], although the difference was not statistically significant (CD14 vs. PL, p = 0.061; CD14 vs. CD4, p = 0.113). The smaller number of mutations detected in monocytes leads to a significantly higher genotypic sensitivity score than that for plasma virus (GSS-CD14 = 13.6 ± 5.5; GSS-PL = 8.7 ± 5.2; p = 0.044) and for CD4-associated virus (GSS-CD4 = 8.9 ± 5.0; p = 0.049) (Table 2).
Genotypic sensitivity score (GSS) monocytes vs. GSS CD4+ T cells and vs. GSS plasma, p < 0.05.
With regards to the 10 patients with virological suppression, sequence analysis was performed only on cell-associated viruses, as plasma viremia was undetectable. In three of these patients no drug-resistant mutations were found in either cellular compartment. Seven patients exhibited drug-resistant mutations in HIV-1 sequences from CD4+ T cells and/or from blood monocytes. Specifically, the mutations at the codon recognized as important in the genotypic resistance pattern differed between monocytes and CD4+ T cells in five patients (71%), whereas only two (29%) showed the same numbers and types of mutations in both compartments.
Correlation between number of years of treatment and numbers of drug-resistant mutations in the distinct blood compartments
Spearman's correlation analysis was used to evaluate whether the numbers of mutations detected in the three distinct blood compartments were associated with different times of treatment. The results, shown in Fig. 2, showed a significant positive correlation between the number of years of therapy and the number of drug-resistant mutations in blood monocytes (r = 0.626; p = 0.002) and in plasma (r = 0.488; p = 0.022). Against this, the correlation between the numbers of mutations in CD4+ T cells and periods of treatment did not reach statistical significance (r = 0.409; p = 0.06).

(
Analysis of the frequency of specific mutations
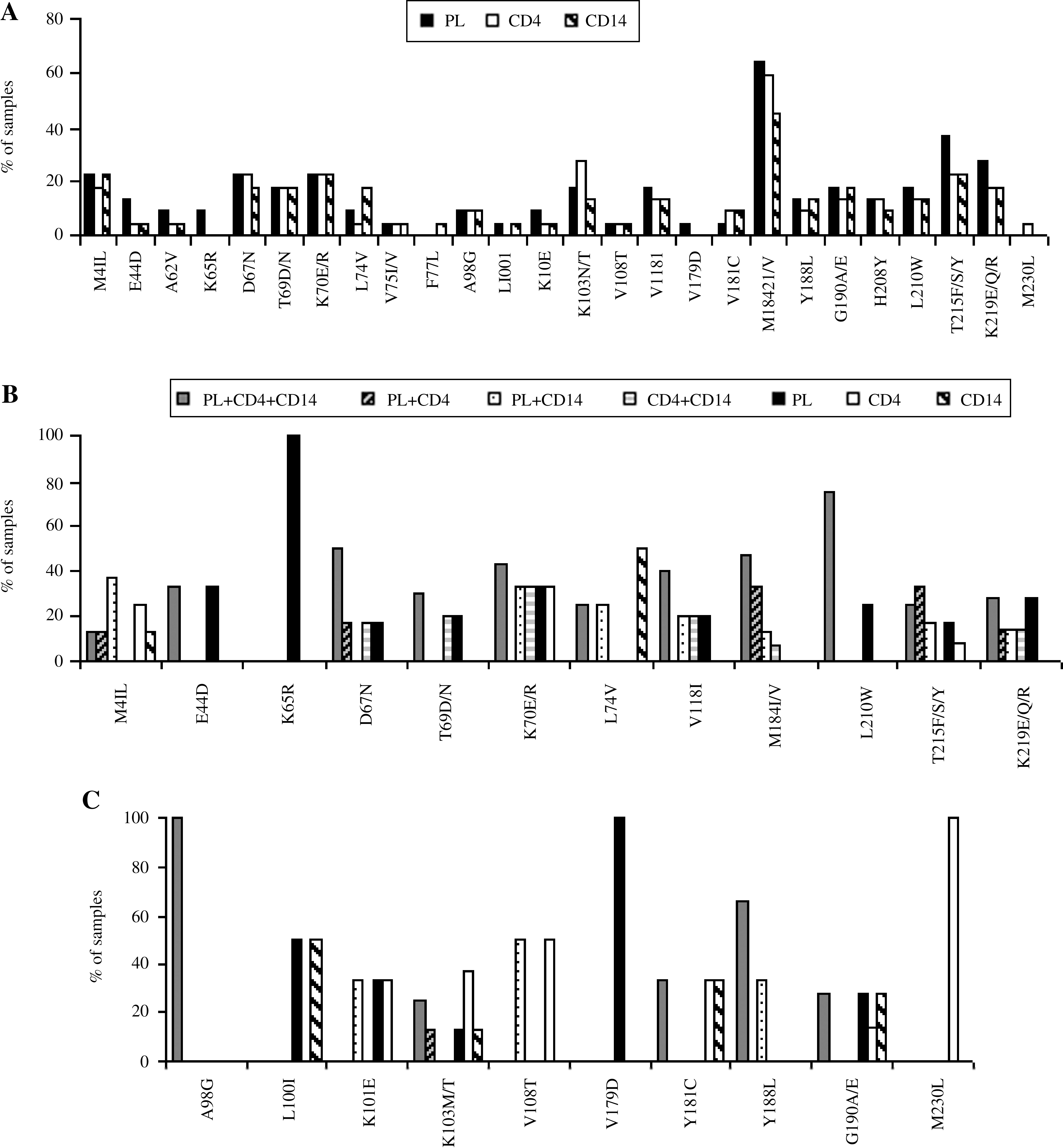
Analysis of the frequency of mutations associated with NRTI and NNRTI resistance (see Fig. 3A) showed that the resistance mutations most frequently detected in all three compartments were M184V (plasma samples, 64%; CD4+ T cells, 59%; monocytes, 45%) and T215Y/F/S (plasma samples, 36%; CD4+ T cells and monocytes, 23%). Although some mutations, such as M184V, K219E/Q/R, and E44D, were more frequently detected in plasma samples, others, such as K103N and Y181C, were more frequently observed in cellular compartments. The results are divided according to the class of the drug.
(
To evaluate the distribution of each mutation in the three compartments, we confirmed the number of compartments in which the mutation was found for all samples.
The results relating to the main mutations associated with NRTI resistance are shown in Fig. 3B. It can be seen that in most patients the mutations were spread across all three compartments. However, the K65R mutation was detected in only one plasma sample and, in some patients, mutations such as L74Vand M41L were detected in blood monocytes only. Interestingly, patients in whom the viral variant M41L mutation was found only in CD14+ monocytes had been treated with thymidine analogues for long periods, but these compounds were not included in the therapeutic regimen administered at the time of the analysis. In two patients the L74V mutation was found only in circulating monocytes; an analysis of their therapeutic histories revealed that one had been previously treated with didanosine and/or abacavir for 4 years and the other with an abacavir-containing regimen for 2 years.
As far as the mutations associated with NNRTI resistance were concerned, the one most frequently observed was K103N/T (8/24 patients) followed by G190A (7/24). As shown in Fig. 3C, some mutations, such as L100I, K103N, Y181C, and G190A, were detected only in blood monocytes. It is of note that patients with these mutations in monocytes had taken NNRTI in previous therapeutic regimens.
Analysis of PI mutations showed that L63P, I13V, and M36I were observed in 75%, 58%, and 54% of plasma virus, respectively. In CD4+ T cells, the L63P, I62V, and L10F mutations occurred in 75%, 66%, and 46% of samples, respectively, while in blood monocytes the most frequently found mutations were L63P, M36I, and I62V, with a frequency of 71%, 50%, and 42%, respectively (Fig. 4A and B).

(
An analysis of the distribution of major mutations associated with PI resistance showed that in most samples, primary mutations were found in all three compartments, or in at least two compartments—plasma and CD4 cells (Fig. 4C). PI major mutations never occurred only in blood monocytes.
In patients with virological suppression (HIV-RNA <50 copies/ml), only cell-associated viruses were analyzed.
The distribution of mutations in CD4+ T cells and blood monocytes showed that 3 out of 10 patients had no drug-resistant mutations and seven displayed drug-resistant viral variants with mutations expressed differently in the two compartments. Some mutations, such as M41L, K103N, M184V, and T215Y, were found only in monocytes (Fig. 5A). Again, the presence of mutations correlated with the previous therapeutic regimen.
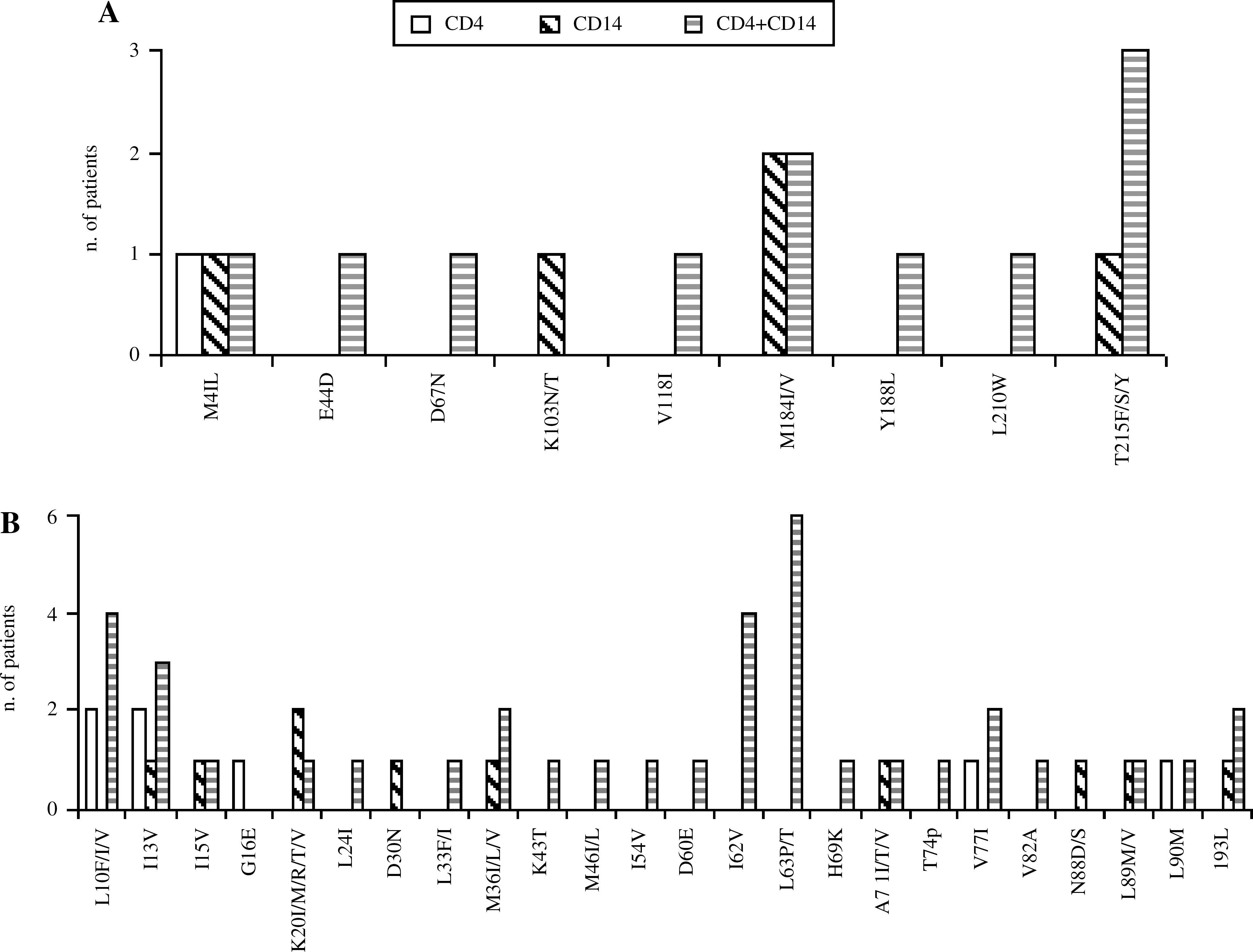
Numbers of virologically suppressed patients harboring mutations in CD4+ cells, CD14+ monocytes, or in both compartments. (
With regard to PI mutations, these were mostly found in both cellular compartments. However, in some patients major mutations, such as D30N and N88S, were detected only in monocytes (Fig. 5B).
Discussion
Previous studies have shown that peripheral blood monocytes can harbor genetically diversified HIV-1 variants that are distinguishable from those present in blood CD4+ T cells. 13,17 –19 Furthermore, others have reported that HIV-1 strains in monocytes are also diverse in phenotype. 20
In the present study, we have demonstrated that monocytes can harbor HIV variants with a pattern of drug-resistant mutations different from those detected in CD4+ T cells and in cell-free virus.
Similar results were obtained when plasma virus was compared with viruses archived in total PBMCs, 27 –29 but few data are available on the resistance pattern of viral populations within individual blood cell types. 30 Potter and colleagues reported that in some individuals, distinct viral populations containing specific drug-resistant mutations were present in different blood compartments. The results reported here are in agreements with these findings, as in 71% of patients failing ART there was a different distribution of drug-resistant mutations in the three analyzed compartments. A number of factors may influence the compartmentalization of viral variants in different blood cell types during art. The penetration of antiretroviral drugs into different cell types may be variable and could alter the selective pressure responsible for the emergence of drug resistance. It should also be taken into account that some physiological factors, such as the expression of some ABC-proteins that can be differently expressed by the diverse cell types, 31 might affect the uptake of drugs, thus determining what mutations and variants emerge. Furthermore, it is also well-known that the immunological system may exert a strong selective pressure on viruses, leading to the emergence of viral variants. 32,33
In general, in failing patients, the virus derived from monocytes contained fewer resistance mutations than were detected in both plasma and CD4+ T cells, suggesting that the development of drug resistance in monocytes may occur at a lower rate. Similar results have been obtained by Aquaro and co-workers in an in vitro study in which the development of resistance in monocytes/macrophages was reported to be lower. 34 There is some evidence that activity of antiretroviral drugs in monocytes is substantially different from that in actively replicating lymphocytes. 35 –37 The decreased probability of the development of resistance in monocytes could be due to a different selective pressure imposed by ART, although the rate of viral turnover may also contribute.
The data also showed that the numbers of drug-resistant mutations found in monocytes and in plasma virus correlated positively with periods of treatment. This was an expected result, as the selection of drug-resistant variants is known to depend strictly on the selective pressure exerted by the drugs. A longer treatment time is usually associated with the wider use of all classes of drugs and with a greater possibility of selecting different mutations
The lack of significant correlation between treatment times and numbers of mutations in CD4+ T cells may suggest that in these cells, the presence of the resistant variant is less related to the period of treatment, as the reservoir of these resistant variants may be established early and maintained for a long time.
This study also included a small group of patients with undetectable levels of plasma viremia. In most of them (70%), drug-resistant HIV strains were detected in both CD4+ T cells and monocytes. As observed in samples from ART-failing patients, in responding patients the profile of drug-resistant mutations in the virus from monocytes was different from that in CD4+ T cells, which suggests that these cells, as well as CD4 T-lymphocytes, may contribute at the level of cellular drug resistance.
Interestingly, in some patients, both those failing and those responding to therapy, monocytes harbored viral variants, probably selected by previous treatment. Given that blood monocytes circulate in the peripheral blood for only a few days, the drug-resistant viral variant detected in monocytes should be from sources of replicating virus that have recently infected blood monocytes. One potential source of monocyte HIV-1 may be tissue macrophages that are productively infected and serve as a source of virus. 14,38 –41 HIV-1 might be produced in tissue macrophages released into the blood and then, preferentially, into infected blood monocytes. 17,18,22 Alternatively, precursors such as CD34+ progenitor cells in the bone marrow could serve as target cells and may be infected by drug-resistant variants, carrying the virus through differentiation into blood monocytes. 42 It is plausible that circulating monocytes could become infected at this stage and carry the virus through differentiation into tissue macrophages. Hence, the presence in monocytes of mutations selected by previously administered drugs could be due to a recycling of virus from precursor to macrophage. Recent findings 20 indicate that blood monocytes can contain a macrophage-specific CCR5 HIV-1 strain, able to infect only monocyte-derived macrophages; hence, it can be further speculated that mutations found only in monocytes belong to a viral variant capable of replication mainly in the monocyte–macrophage lineage.
It is of note that the presence of detectable HIV-DNA in monocytes during suppressive ART is in agreement with some previous findings, 13,15,16,18,30,38 although others 11 have reported that in suppressive ART, neither monocytes nor monocyte-derived macrophages harbored HIV, which suggests that the HIV infection of monocytes is directly related to the success of the therapy. This discrepancy could be caused by the characteristics of the population analyzed; our findings are based on monocytes derived from patients who have a long therapeutic history with more than one virological failure. It is presumed that in these patients who harbored resistant viral variants, ART was able to suppress the replication in plasma virus but did not reduce the HIV-DNA load below the limits of detection. This hypothesis is supported by the findings of Zhu and co-workers, who showed that in patients on effective ART the decay of HIV-DNA in CD14+ monocytes was slower than that in activated or resting CD4+ T cells. 22
The limitations of the present study include the small number of patients and the use of a procedure based on direct sequencing of a PCR product that does not allow the detection of minor viral variants. Nevertheless, the data presented here indicate that CD14+ monocytes may harbor a dominant viral population with a drug-resistant mutation pattern different from that detected in CD4+ T cells and in cell-free virus. Although recent studies indicate that HIV-1 replication occurs in blood monocytes in vivo, 16,43 it remains unclear to what extent HIV-1 can infect and replicate in blood monocytes. Blood monocytes are able to migrate to all tissues, such as lungs, gastrointestinal tract, kidney, urogenital tract, primary and secondary lymphoid organs, and the central nervous system. Any viral variants harbored in monocytes could be carried into these tissues where they could replicate, thus contributing, together with the resistant viruses harbored in CD4+ T cells, to the persistence of the drug-resistant virus.
It is important to note that we were unable, in this study, to reach any definite conclusions about the role played by the monocytes, compared with CD4+ T cells, in acting as a source of archived resistant viruses. Rather, our data seem to indicate that the establishment of an archive in monocytes, and probably in CD4+ T cells, is, in some way, unpredictable because it may depend not only on drug pressure, but also on the host immune pressure and on the dynamic network of cytokines and activated immune cells.
Footnotes
Acknowledgments
This work was supported by grants from “Sapienza” University of Rome, Italy, to G.A. and to O.T.
Author Disclosure Statement
No competing financial interests exist.