
Abstract
We examine the distribution of viral genetic forms and the presence of antiretroviral drug resistance mutations in HIV-1 infections in the Republic of Dagestan, in the North Caucasus area of Russia, where a recent large increase in HIV-1 infections has been documented. Samples were collected from 41 HIV-1-infected individuals from Dagestan, most of them from the cities of Derbent (n = 21) and Mahachkala (n = 18). Thirty six were injecting drug users and five were infected by heterosexual contact. None was on antiretroviral drug treatment. HIV-1 protease and a segment of reverse transcriptase were amplified by RT-PCR from plasma RNA and sequenced, and phylogenetic trees were constructed via maximum likelihood. Forty (97.6%) of 41 samples were of subtype A, former Soviet Union variant (AFSU), of which 27 (67.5%) clustered with the subvariant containing the V77I substitution in protease (V77IPR). Within this cluster, 13 viruses formed a local subcluster, 10 of which were from Derbent. Four viruses clustered with the ASP2 subcluster, recently identified in St. Petersburg, two with a virus from Georgia and one with a virus from Azerbaijan. No mutations associated with antiretroviral drug resistance were detected. The results, therefore, show the relationship of the HIV-1 epidemic in Dagestan with that of other areas of Russia and of neighboring countries, and reveal the spread of the AFSU V77IPR variant in the North Caucasus area.
R
In recent years there have been reports on a rapid increase of HIV-1 infections in the North Caucasus area of Russia, including Chechnya and Dagestan 15 (Fig. 1). However, no data on the distribution of HIV-1 variants or on the prevalence of drug resistance mutations in this area of the Russian Federation have been published. In the Caucasus area, there are reports only from the neighboring South Caucasus countries of Georgia and Azerbaijan, in which AFSU is predominant, representing 70% and 90.2% of the infections, respectively, with subtype B representing 26% and 7.3% of the infections, respectively. 16,17
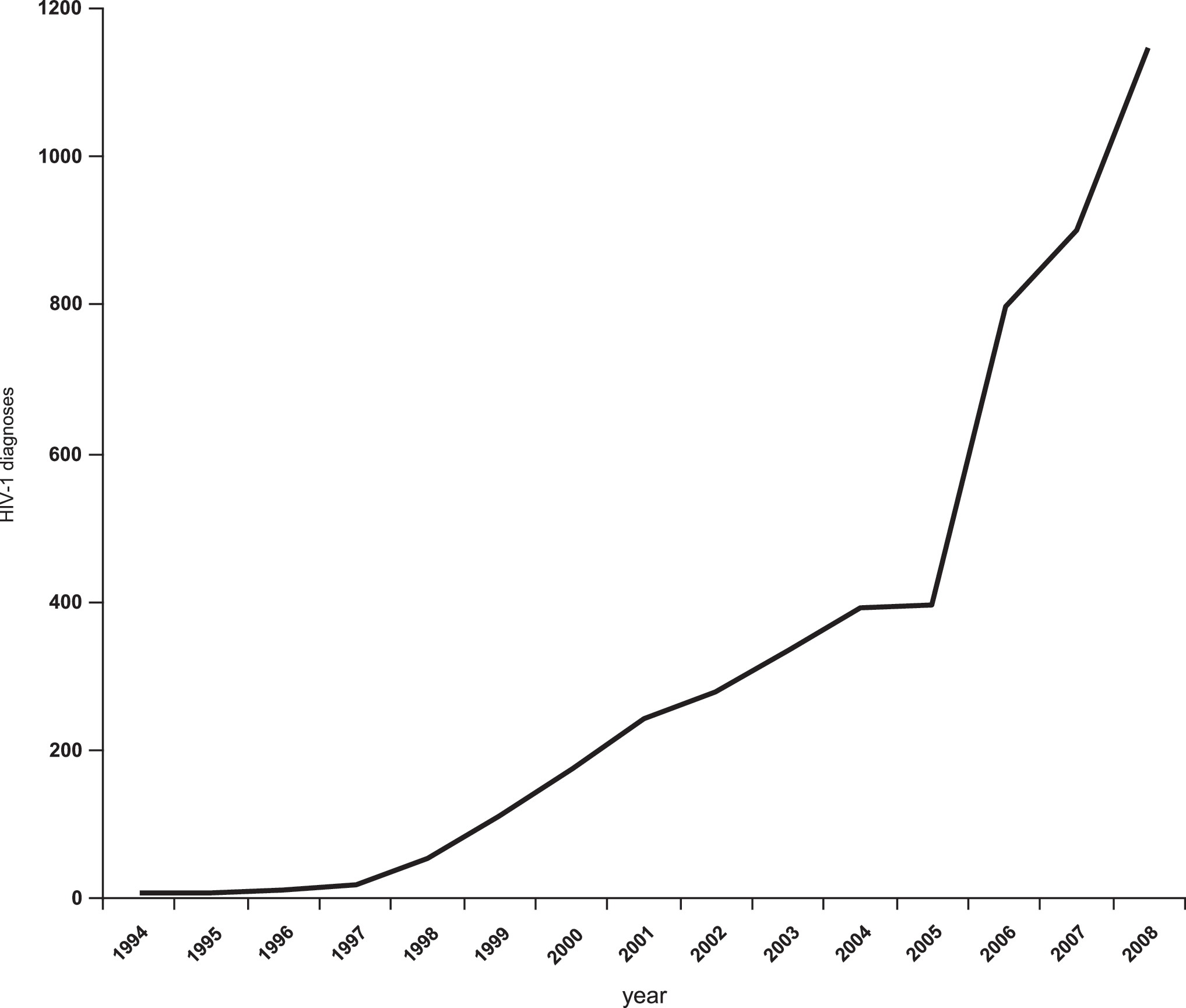
Chronological evolution of the HIV-1 epidemic in Dagestan. 15
In this study, we analyzed 41 samples from HIV-1-infected individuals collected in Dagestan in November 2006. Subjects were from the cities of Derbent (n = 21), Mahachkala (n = 18), Hasaburt (n = 1), and Buinaks (n = 1). Samples from Derbent were collected at the local prison and those from residents in Mahachkala, Hasaburt, and Buinaks were collected at the Republic's Center for Infectious Diseases, AIDS and Prevention in Mahachkala. Thirty six were injecting drug users and five were infected by heterosexual contact. Ages ranged from 20 to 52 years old (mean 28.1). None was on antiretroviral drug treatment. RNA extraction from plasma and reverse transcriptase polymerase chain reaction (RT-PCR) amplification and sequencing of protease and a segment of reverse transcriptase (PR-RT) were done as previously described. 18 Briefly, RNA was extracted from plasma using the reagents provided in the Nuclisens kit (Biomerieux, Marcy l'Etoile, France); RT-PCR was performed with a mixture of SuperScript III (Invitrogen, Carlsbad, CA) and Biotaq DNA polymerase (Bioline, London, UK), followed by nested PCR performed with Biotaq DNA polymerase, enzymatic purification of the PCR product using exonuclease I and shrimp alkaline phosphatase, and sequencing with the ABI Prism BigDye Terminator kit, with analysis in ABI PRISM 3700 automated sequencer (Applied Biosystems, Foster City, CA). The sequenced segment used for phylogenetic analysis is 1.2 kb long, comprising nucleotides 2253–3452 in the HXB2 proviral genome. Phylogenetic trees were constructed via maximum likelihood (ML) using Treefinder. 19 For tree construction, we included AFSU references, and within this variant, sequences of viruses representative of the V77IPR subvariant, and the recently identified St. Petersburg clusters (ASP1, ASP2, and ASP3). The best-fit model of sequence evolution was selected under the Akaike information criterion using jModeltest v.0.1.1. 20 The reliability of tree topologies was assessed using 100 bootstrap replicates. The presence of antiretroviral drug resistance-associated mutations was analyzed with the Stanford University HIVdb Program. 21
The ML tree obtained with the Dagestan and reference sequences is shown in Fig. 2. Forty sequences clustered with the AFSU variant, and only one sequence was of subtype B. Of the 40 AFSU sequences, 27 (67.5%) formed a cluster corresponding to the V77IPR subvariant, and 4 (10%) formed another cluster nested within the ASP2 subcluster, a minor AFSU variant of St. Petersburg. 11 Within the V77IPR clade, 13 (48.1%) viruses formed a local subcluster, 10 of which were from Derbent (Figs. 2 and 3). To examine the relationship of the Dagestan viruses with viruses from neighboring areas, sequences of all subtype A sequences from Georgia 16 and Azerbaijan 17 available at the Los Alamos HIV Sequence Database 22 were included in the alignment, which was analyzed with Treefinder. In the resulting tree, a virus from Dagestan (Dag42) clustered with a virus from Azerbaijan (02AZ098) and another two viruses of Dagestan (Dag03 and Dag35) clustered with a virus from Georgia (03GEMZ008), with 91% and 84% bootstrap values, respectively (results not shown). No antiretroviral drug resistance mutations were detected in any of the samples.

Maximum likelihood phylogenetic tree of PR-RT sequences from Dagestan. The analyzed segment comprises nucleotide positions 2253–3452 in the HXB2 proviral genome. Names of viruses from Dagestan start with Dag. AFSU references are underlined. Sequences preceded by ASP1, ASP2, and ASP3 represent recently described clusters of St. Petersburg. The tree was constructed with Treefinder 19 using the TVM + Γ + I substitution model selected by jModeltest 20 under the Akaike information criterion. A subcluster of viruses comprising viruses mainly from Derbent is bracketed on the right. Only bootstrap values of 70% or greater of relevant clusters are shown.
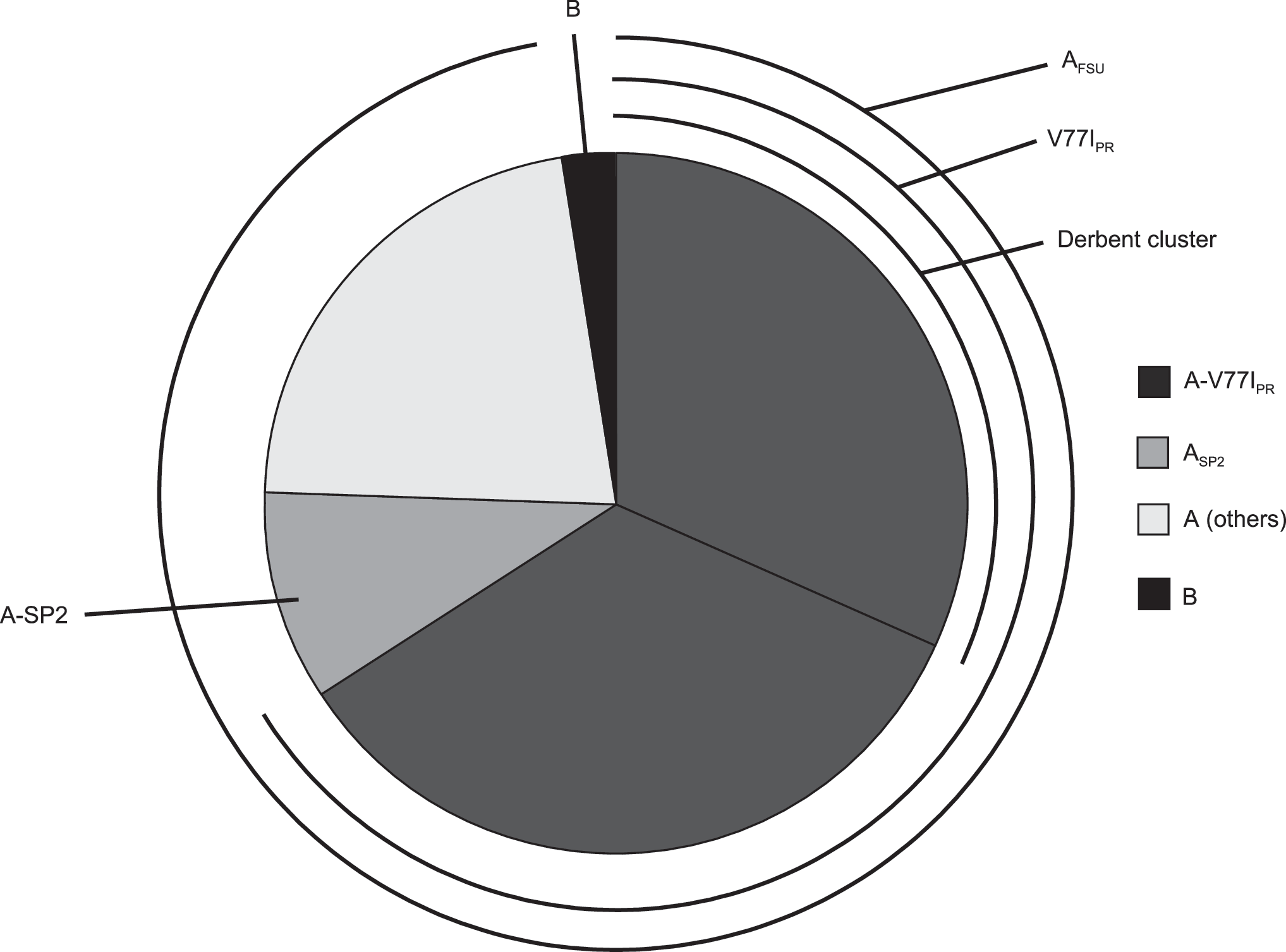
Distribution of HIV-1 clades in Dagestan.
The results, therefore, indicate an overwhelming predominance (97.6%) of the AFSU variant in the Dagestan epidemic. This is similar to the prevalence reported in Azerbaijan (90.2%), 17 and higher than that reported in Georgia (70%). 16 Among the AFSU viruses, the V77IPR subvariant is predominant, representing approximately two-thirds of the viruses. This subvariant has also been detected in Georgia, where it represents 36.4% of AFSU viruses. 16 Subcluster analysis has not been reported in Azerbaijan, 17 but our own analysis with sequences retrieved from databases shows that 44.8% AFSU viruses from this country are of the V77IPR subvariant (results not shown). Further phylogenetic structure was detected in the V77IPR subvariant, with 10 viruses from Derbent and 3 from Mahachkala forming a subcluster nested within it.
The results reported here and elsewhere 4,6,10,11 show that the AFSU variant has undergone multiple successive splits along its expansion in FSU countries, representing successive introductions at different geographic areas and at different times. Some subvariants have remained largely confined to a geographic area, such as those recently identified in St. Petersburg, although sporadic introductions have occurred into distant areas, 11 including, as here reported, Dagestan. On the other hand, one of the AFSU subvariants, V77IPR is widely circulating at different geographic areas of Russia and other AFSU countries, representing a substantial proportion of viruses in some of them. 4,10,11,16,17,23 We recently reported its presence in 17.4% of AFSU samples from St Petersburg, 11 and in a previous study on samples from diverse FSU countries, the great majority of them from Russia, we found V77IPR in 49.4% subtype A viruses. Outside of Russia, it is also circulating in Georgia (36.4%), 16 Azerbaijan (44.8%), 17 and Kazakhstan (58.8%). 23 The current study now reports its presence in Dagestan, where it represents 67.5% of AFSU viruses in this Republic of the Russian Federation. This subvariant has undergone further splits, as shown here in the cluster identified in Derbent. The molecular epidemiological study of the HIV-1 epidemic in FSU countries, therefore, reveals the progressively increasing complexity of the HIV-1 evolution along its expansion, with successive splits of HIV-1 clades into different subclades and subsequent formation of intraclade recombinant forms (unpublished results). The increasing complexity of the HIV-1 evolution along its expansion, as evidenced in the AFSU epidemic, should be taken into consideration in a future revision of the nomenclature system of HIV-1 genetic forms, which is not properly addressed by the current classification. 24
The identification of large proportions of viruses grouping into phylogenetic clusters within the AFSU variant is in contrast to findings in most countries where subtype B is predominant, in which, with some exceptions, 25,26,27 no such structured phylogenies are found, except for relatively minor clusters of recent origin. 28,29 One of the reasons for this difference may be that in longstanding epidemics, such as those of subtype B in Western countries, the distinction among clusters may be blurred by extensive intrasubtype recombination, which accumulates progressively over time and interferes with the reconstruction of the original phylogenetic structure of the viruses by using the usual phylogeny inference methods. In the case of the subtype AFSU epidemic, being relatively young, the original phylogenetic structure may still remain relatively intact, allowing reconstruction of phylogenetic clusters with commonly used programs.
In conclusion, the HIV-1 epidemic in Dagestan is largely dominated by the AFSU variant, with a predominance of the V77IPR subvariant, within which approximately half of the viruses belong to a local subcluster, most of them from the city of Derbent. Sporadic relationships were found with viruses from St. Petersburg, Georgia, and Azerbaijan. The phylogenetic characterization and the knowledge on the distribution of HIV-1 variants circulating in Russia and other FSU countries will contribute to tracking the expansion of the epidemic in this geographic area and may be important in examining correlations of HIV-1 clades with immune responses relevant for vaccine development and with biological features of the virus.
Footnotes
Acknowledgments
We thank Pablo Martínez, Aurora de Miguel, and Ana Parejo from the Genomic Unit, Centro Nacional de Microbiología, Instituto de Salud Carlos III, Majadahonda, Madrid, Spain, for technical assistance in sequencing. This work was funded through a technical service agreement with WHO-UNAIDS, OD/TS-07-00446. Anna Vinogradova was supported through a scholarship by the HIV Research Trust.
Sequences have been deposited in GenBank, with accession numbers GQ867596 through GQ867636.
Author Disclosure Statement
No competing financial interests exist.