
Abstract
The Tax protein encoded by HTLV-1 plays a key role in the development of ATL in infected individuals. Our previous studies showed that tax transgenic mice develop disease that is almost identical to human ATL, with widespread organ invasion by lymphomatous cells and the development of leukemia. The same pathology develops rapidly in SCID mice engrafted with cells from the transgenic animals. In the present study, we used this SCID model to analyze the expression levels of several cytokines, growth factors, and adhesion molecules to determine their possible involvement in the development of disease. We showed that Tax expression was undetectable at the protein level in the tax-transformed cells used to inoculate the SCID mice and that these cells displayed constitutive NF-κB and Akt activity. We demonstrated significant differences in the levels of circulating PDGF-BB, TNF-α, sICAM-1, and sVCAM-1 in inoculated animals. Cell-surface staining of the tax transgenic cells showed that they do not express receptors for any of the upregulated growth factors. Significant differences were not found in the secreted levels of bFGF, MMP9, VEGF, or E-selectin, whereas IL-2, IL-15, IL-6, IL-1β, and IFN-γ expression was undetectable. Even though the number of factors analyzed is limited, our study identified TNF-α, PDGF-BB, and the adhesion molecules sICAM-1 and sVCAM-1 as factors that may contribute to the high levels of organ infiltration by leukemic cells in this tax transgenic SCID model.
Introduction
H
The pleiotropic function of Tax 1 is highly related to its ability to interact with a multitude of cellular proteins. Tax 1 interacts with >100 cellular proteins, including transcription factors, cell-signaling and cytoskeleton proteins, cell-cycle and PDZ domain–containing proteins, and transport proteins. 11 Through these interactions, Tax 1 modulates several cellular transcription pathways including NF-κB, Akt, cyclic AMP response element–binding protein/activating transcription factor (CREB/ATF), Sp1, AP-1, and serum response factor. 12 Even though activation of the NF-κB pathway by Tax 1 is essential for transformation, 13 Tax mutants that fail to activate NF-κB have also been shown to immortalize T cells, indicating that additional pathways may be involved in this process. 14 The PI3K/Akt prosurvival pathway has also been implicated in Tax-mediated transformation. 15 Several studies have shown that Akt is activated in HTLV-1–transformed cells and is linked to NF-κB activation, p53 inhibition, and cell survival. 16 Modulation of these and other pathways by Tax 1 has been shown to activate the expression of several cellular proteins, including cytokines such as interleukin-1α (IL-1α), interleukin-2 (IL-2), interleukin-6 (IL-6), granulocyte macrophage colony–stimulating factor (GM-CSF), tumor necrosis factor α (TNF-α), interleukin-15 (IL-15), 17 –20 growth factors such as platelet-derived growth factor (PDGF), 21 vascular endothelial growth factor (VEGF), 22 and intercellular adhesion molecule-1 (ICAM-1) 23 and vascular cellular adhesion molecule-1 (VCAM-1). 24 The overexpression of many of these factors in HTLV-infected individuals is speculated to contribute to the pathogenesis of both ATL and TSP/HAM.
The oncogenic potential of Tax 1 is clearly demonstrated by in vivo studies, which show that tax transgenic animals develop a range of neoplasias of lymphoid and nonlymphoid origins, depending on the cellular compartment that expresses Tax. 25 –27 In a previous study, we generated tax transgenic in which the expression of Tax 1 was targeted to developing thymocytes by using the proximal Lck promoter. 26 After prolonged latency periods, transgenic mice developed disease almost identical to human ATL, with widespread organ invasion by lymphomatous cells and the development of leukemia. The same pathology developed in mice with severe combined immunodeficiency (SCID) engrafted with cells from the transgenic animals after a relatively short period.
In the present study, we examined the expression levels of a number of cytokines, cytokine receptors, growth factors, and adhesion molecules in this SCID model to investigate their involvement in the pathogenesis of this disease.
Materials and Methods
Cells and inoculation of mice
A pre–T-cell line, derived from mice containing the HTLV-1 tax gene, 26 was cultured in RPMI containing 10% fetal calf serum (FCS), 100 units/ml penicillin/streptomycin, 10 μg/ml kanamycin, and 50 μM β mercaptoethanol. SCID mice (n = 6) were inoculated with 106 cells by intraperitoneal injection and were monitored for 4–6 weeks, after which blood was obtained by cardiac puncture under terminal anesthetic. Ascites cells from the abdominal cavity were subsequently used to inoculate further a second group of SCID mice (n = 8), and serum was obtained as described earlier. Noninoculated SCID mice were used as a control (n = 5 for both sets). SCID mice were purchased from Charles River Laboratories (Margate, UK). Animal experiments were approved by the Animal Research Ethics Committee in University College Dublin and by the Department of Health and Children, Ireland.
RT-PCR
Total RNA was extracted from ∼5 × 106 cells, as indicated, by using an RNeasy Mini Kit (Qiagen, Crawley, UK) according to the manufacturer's protocol. All real-time PCRs were performed on the Roche LightCycler platform by using the QuantiTect SYBR Green PCR Kit (Qiagen). Primers directed against tax 1 28 and the 18s rRNA internal reference 29 were described previously. PCR-cycle parameters comprised an initial incubation at 95°C for 15 min, followed by 40 cycles of 95°C for 15 s, 55°C for 30 s, and 72°C for 30 s). Relative quantification was carried out according to the Pfaffl method, 30 normalizing expression levels within samples by using the 18s rRNA internal control and quantifying against expression levels seen in MT2 cells. IL-2 and IL-15 amplification was carried out as previously described. 31,32
Nuclear extraction and electromobility shift assay (EMSA)
Nuclear extracts were prepared from the tax transgenic mouse cell line and ascites cells by using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Scientific, Rockfield). EMSA, by using biotin-labeled NF-κB consensus oligonucleotides, was carried out on 3 μg of the indicated nuclear extracts by using the Pierce LightShift Chemiluminescent EMSA kit, according to the manufacturer's instructions.
Western blotting
The 30 μg of lysates (as determined by the BCA Protein Assay kit; Thermo Scientific) from the tax transgenic cell line, ascites cells, MT2 cells, and CEM cells was subjected to SDS/PAGE by standard protocols. Tax 1 expression was detected by chemiluminescence with the mouse anti-Tax monoclonal antibody 1A3 (Abcam, Cambridge, U.K.) followed by anti-mouse HRP. The β-actin was detected by using the mouse monoclonal AC-15 (Abcam). The unphosphorylated and phosphorylated forms of Akt were detected in lysates from cells incubated with 100 nM serine/threonine kinase inhibitor Calyculin A (Cell Signaling, Danvers) for 1 h before lysing the cells, by using the Rabbit monoclonal C67E7 (Cell Signaling) and the Phospho-Akt (Ser 473) rabbit monoclonal D9E (Cell Signaling).
Multiplex analysis
Simultaneous growth-factor detection was carried out by using the Mouse Growth Factor 4-Plex Kit (Invitrogen Life Science) for the measurement of mouse VEGF, PDGF-BB, bFGF, and G-CSF. The four-plex GF kit was used in combination with single-plex kits for TNF-α, IFN-γ, IL-6, and IL-1β (Invitrogen Life Science) for a combined eight-plex assay. Multiplex analysis of MMP-9, sICAM-1, sVCAM-1, and sE-selectin was carried out by using Mouse Cardiovascular Disease (CVD) Panel 1 LINCOplex Kit (Invitrogen Life Science). All assays were carried out twice, according to manufacturer's instructions.
Murine IL-15 ELISA
Serum levels of murine IL-15 protein were assessed by using the “Ready-Set-Go” Mouse IL-15 ELISA kit (eBioscience, San Diego, CA), according to the manufacturer's protocol. All serum samples were assayed in duplicate or triplicate.
Statistical analysis
Statistical analyses were performed by using Statview software version 5.0.1 (SAS Institute, Cary, NC). The Mann–Whitney U test was used to compare analyte levels between inoculated and control mice, and between Group 1 and Group 2 mice. Correlations among analyte levels, or between analyte levels and spleen weights, were determined by using Spearman's Correlation coefficient.
Flow cytometric analysis of cell-surface markers
FITC-conjugated anti-CD3 and anti-CD44, APC conjugated anti-CD25 and anti-CD69 antibodies, and the PE-conjugated anti-VEGF-R2, along with appropriate isotype controls, were obtained from BD Pharmingen (Oxford, UK). The VEGF-R1 IgG1 antibody (AbCam Cambridge, UK) or isotype control was used with TxRD anti-mouse IgG1 secondary antibody (Southern Biotech). FITC conjugated anti-VCAM-1 and ICAM-1 and PE-conjugated PDGFR-β were purchased from eBioscience. Sample acquisition was performed by using a BD FACS Calibur flow cytometer, and data were analyzed by using CellQuest software.
Results
Phenotypic analysis of the Lck tax transgenic cells
SCID mice were inoculated by intraperitoneal injection either with a cell line established from the original Lck tax transgenic animals (Group 1) or with ascites cells derived from the Group 1 animals (Group 2). Flow-cytometric analysis of the cell line and ascites cells (data not shown) showed that these cells expressed CD25, CD44, and CD69 on their cell surface, whereas CD3 was predominantly visible only when cells were stained after membrane permeabilization (Fig. 1). Analysis of Tax 1 expression in these cells showed low levels of expression at the RNA level and undetectable levels at the protein level compared with endogenous levels of Tax 1 expression in the HTLV-1–infected MT-2 cell line (Fig. 2). It is well established that Tax 1 modulates gene expression and associated cellular events by activating signaling pathways, including NF-κB and Akt. To determine the involvement of these pathways in the pathogenesis of disease observed in inoculated animals, we performed EMSAs by using an NF-κB probe and nuclear extracts from the tax transgenic cells and ascites cells. These assays showed that the NF-κB pathway was constitutively activated in both cell types, with the ascites cells showing higher levels of activity (Fig. 3A). In addition, Western blot analysis of lysates from the tax transgenic cell line, incubated with the phosphatase inhibitor calyculin A, showed the presence of the active phosphorylated (position 473) form of Akt (Fig. 3B).

Phenotypic analysis of the tax-transgenic cell line. Surface expression of CD3, CD25, CD44, and CD69 was examined (
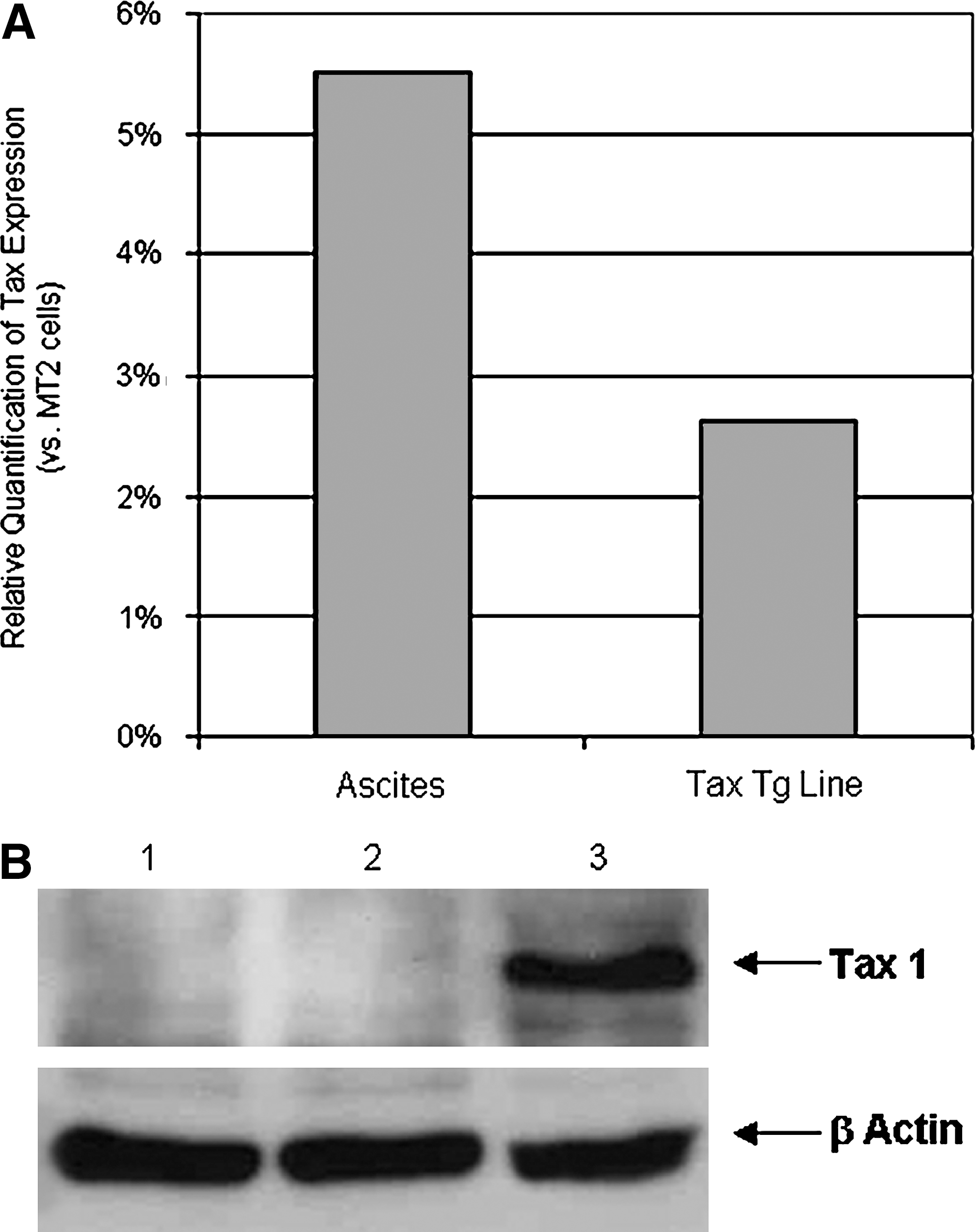
Tax 1 expression in the murine tax transgenic cell line and ascites cells. (
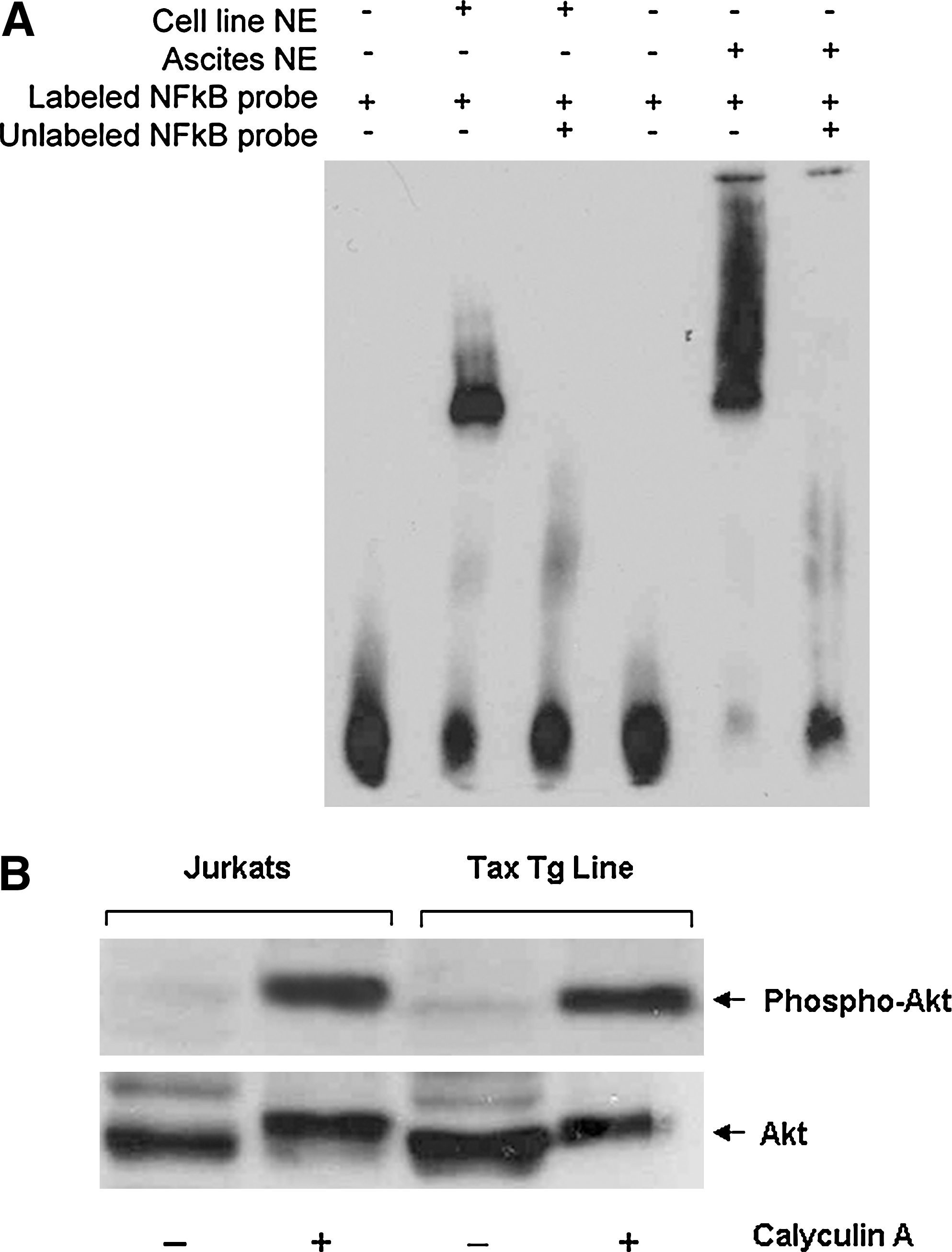
NF-κB and Akt activity in the tax transgenic cells. (
Lymphoma and leukemia development in SCID mice inoculated with a cell line derived from tax-transgenic animals
The most pronounced pathologic effect observed in inoculated animals was the development of marked splenomegaly (Fig. 4A), with the spleen weights of inoculated mice being significantly different from those of controls [median (inoculated) = 0.5 g; median (control) = 0.03 g; p = 0.005]. Peripheral blood smears from inoculated animals showed the presence of a large number of leukemic cells (Fig. 4B)

Gross pathologic findings in SCID mice inoculated with tax-transgenic cell line compared with controls. (
Absence of IL-2 and IL-15 expression by tax-transgenic cell line
To determine the possible involvement of IL-2 and IL-15 in the pathogenesis of disease in inoculated SCID mice, we examined the expression of both cytokines in the tax transgenic cell line with RT-PCR. As shown in Fig. 5, IL-15 mRNA levels were elevated in these cells in contrast to the levels of IL-2 mRNA, which was undetectable. cDNA from a T-cell library was used as a positive control for IL-2 and IL-15 amplification, and reactions with no templates, as negative controls. Despite the presence of elevated levels of IL-15 mRNA, secreted IL-15 could not be detected in the serum of inoculated animals (data not shown).
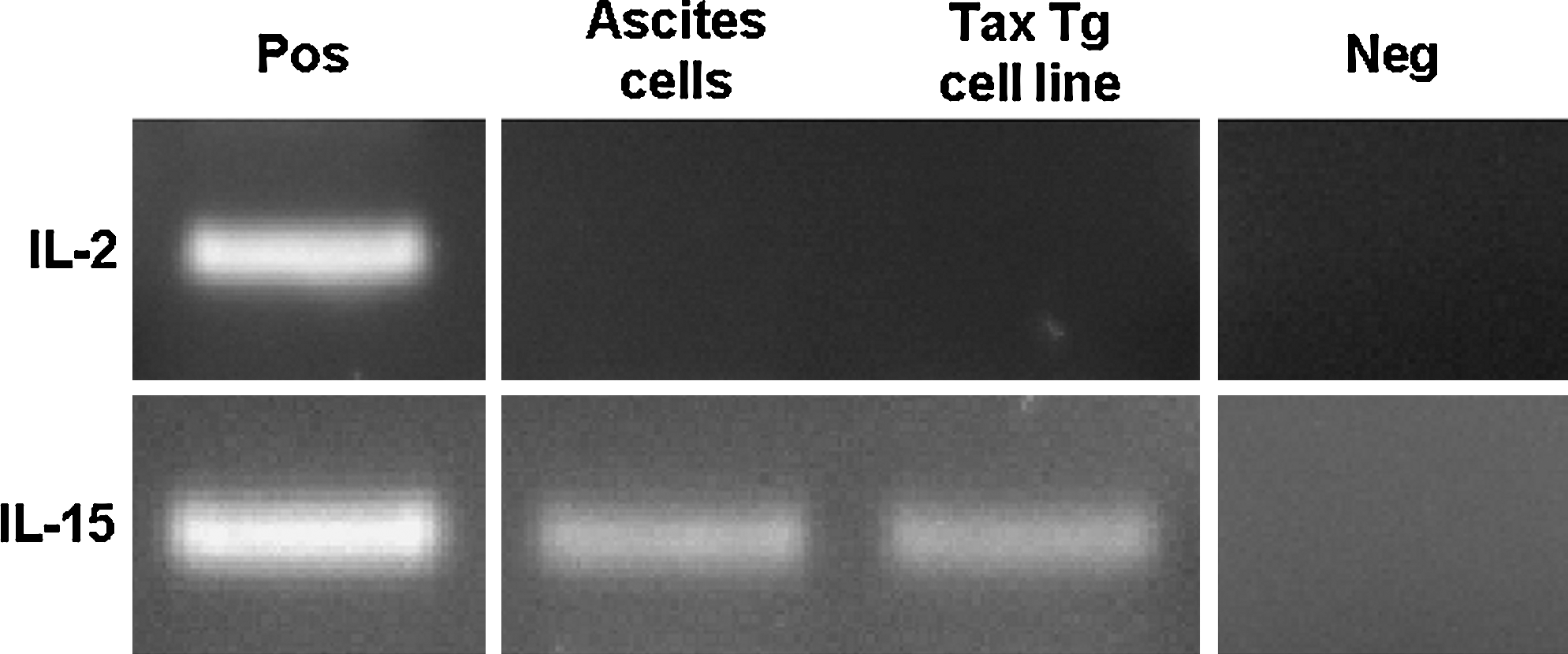
Expression of IL-15 but not IL-2 mRNA in tax-transgenic cell line and ascites cells from inoculated SCID mice. Expression of IL-2 and IL-15 mRNA in cells obtained from ascites fluid, or the murine tax cell line, was determined by RT-PCR. A T-cell cDNA library was used as a positive control in each reaction. Reactions without template were included as negative controls.
Elevated levels of soluble TNF-α, PDGF-BB, but not IL-1β, G-CSF, IL-6, IFN-γ, bFGF, or VEGF are associated with the pathogenesis of disease in inoculated SCID animals
To investigate the factors that may be involved in the high levels of tissue infiltration by cells in the SCID animals, the Luminex xMAP system was used for the simultaneous analysis of the in vivo expression levels of VEGF, bFGF, PDGF-BB, and G-CSF growth factors, the cytokines TNF-α, IL-1β, IL-6, and IFN-γ in serum from animals inoculated with a cell line established from a tax-transgenic mouse (n = 6), and control mice (n = 5) (Group 1; Fig. 6). Levels of PDGF-BB, TNF-α, and G-CSF were all significantly upregulated in the inoculated mice compared with the control mice (p = 0.005, p = 0.005, and p = 0.009, respectively) (Fig. 6A–C). Levels of IL-1β, IL-6, IFN-γ, or VEGF were below the level of detection of the assay, suggesting that these factors may not be involved in the infiltration of cells in this model. A significant difference was not found for the expression of bFGF between the control and inoculated mice (Fig. 6D).

Levels of growth factors and adhesion molecules in the serum of SCID mice inoculated with the tax-transgenic cell line compared with controls. SCID mice were inoculated with a cell line derived from Lck-tax transgenic mice. Approximately 28 days after inoculation, blood was obtained from the mice, and the levels of (
Cells obtained from the ascites in the abdominal cavity of the first group of mice were then used to inoculate a second group of mice (n = 8) in an attempt to determine if a second round of amplification in mice would alter the growth-factor expression profile of these cells in vivo. Multiplex analysis was repeated on serum from seven of the eight inoculated mice (Fig. 7). Overall, the growth-factor expression patterns between the two groups followed a similar pattern, with some notable differences. Specifically, the levels of expression of PDGF-BB and TNF-α were significantly increased in Group 2 inoculated mice compared with Group 1 (p = 0.007 and p = 0.006, respectively) (compare Fig. 7A and B with Fig. 6A and B). Consistent with the first group, levels of both growth factors were significantly elevated in serum from the inoculated mice in Group 2 compared with the control mice (p = 0.003 in both cases) (Fig. 7A, B), and no significant difference was found in bFGF levels (Fig. 7D). However, whereas the first group of inoculated mice had shown a significant increase in levels of G-CSF (p = 0.009), no difference was observed in the second group (Fig. 7C). Interestingly, VEGF was detected in four of the seven animals inoculated with the ascites cells, but this result was not statistically significant (Fig. 7I), whereas VEGF was not detected in the first group of animals. Similar to the first group, the levels of IL-1β, IL-6, and IFN-γ were all below the level of detection of the assay.

Growth factor and adhesion molecule levels in SCID mice inoculated with ascites cells from mice in Fig. 6. The levels of (
Elevated levels of sICAM-1 and sVCAM-1 but not MMP-9 or sE-selectin in inoculated SCID mice
The expression levels of MMP-9, sICAM-1, sVCAM-1, and sE-selectin were also examined in serum samples from the inoculated and control mice. The difference in MMP-9 levels between inoculated and control mice was not significant in Group 1 animals; however, levels were approaching significance in Group 2 mice versus controls (p = 0.08) (compare Fig. 6E and Fig. 7E). A significant difference in the levels of sICAM-1 and sVCAM-1 was observed between the inoculated and the control mice in both Group 1 mice (p = 0.009 and p = 0.006, respectively, Fig. 6F, G) and Group 2 mice (p = 0.005 in both cases, Fig. 7F, G). Conversely, levels of sE-selectin were comparable between the two groups, with no significant difference found between control and inoculated mice in either case (Figs. 6H and 7H).
Correlations of growth-factor levels
Spearman correlation coefficients were calculated between pairs of upregulated growth factors and adhesion molecules identified in both groups of animals. Statistically significant correlations were found between only sICAM-1 and sVCAM-1 in Group 1 animals (p = 0.05; data not shown). Conversely, all growth-factor correlations in Group 2 were significant (Table 1). Positive correlations were found for TNF-α expression with PDGF-BB (R = 0.972; p < 0.04), VEGF (R = 0.979; p < 0.03), sICAM-1 (R = 0.822; p < 0.05), and sVCAM-1 (R = 0.847; p < 0.04). In addition, PDGF positively correlated with VEGF (R = 0.992; p < 0.03), sICAM-1 (R = 0.918; p < 0.05), and sVCAM-1 (R = 0.900; p = 0.04). VEGF demonstrated positive correlations with sICAM-1 (R = 0.753; p < 0.04) and sVCAM-1 (R = 0.871; p < 0.04). As with the Group 1 data, levels of sICAM-1 also demonstrated positive correlation with sVCAM-1 (R = 0.798; p = 0.02).
p < 0.05.
p < 0.04.
p < 0.03.
p < 0.02.
VEGF-R1, VEGF-R2, or PDGFR-β is not expressed by tax-transgenic cells
Many studies have shown that the concomitant expression of growth factors and their respective cell-surface receptors can result in the activation of signaling pathways that facilitate the tissue invasion by malignant cells by either autocrine or paracrine mechanisms. Even though VEGF was detected in only some animals, we examined whether the VEGF receptors VEGF-R1 or VEGF-R2 were expressed on the tax-transgenic cells. As shown in Fig. 8D and E, neither receptor was expressed. Similarly, the PDGF-BB receptor PDGFR-β was not detected (Fig. 8C). In addition, we analyzed the cell-surface expression of the ICAM-1 and VCAM-1, as previous studies showed that enhanced levels of the soluble forms of both these adhesion molecules may arise because of shedding of the proteins from the cell surface. 33 In contrast to VCAM-1, which was not expressed, ICAM-1 was expressed on the surface of the majority of cells (Fig. 8A, B).
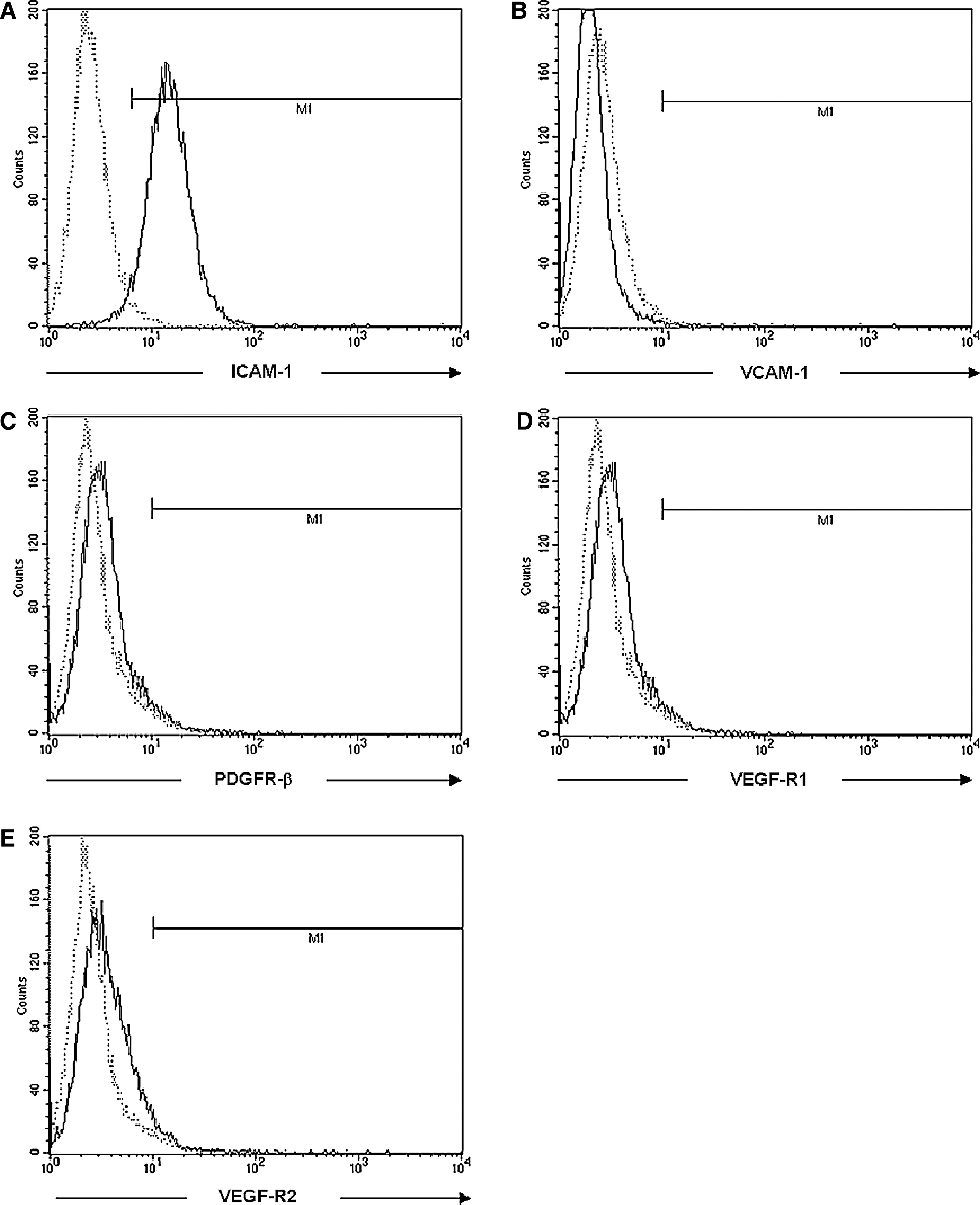
Flow cytometric analysis of cytokine receptor and adhesion molecule expression on leukemic tax-transgenic cells. The surface expression of (
Discussion
In this study, we examined the expression levels of several cytokines, growth factors, and adhesion molecules in serum from SCID mice inoculated with either a cell line derived from tax-transgenic animals or ascites cells from inoculated SCID mice. Phenotypic analysis of cells used to inoculate animals in the present study showed that they are positive for the expression of iCD3, CD25, CD44, and CD69, but not CD4− and CD8−, consistent with the original thymus-derived pre–T-cell phenotype of the lymphoma cells from the tax-transgenic mice. 26 In vivo, these cells displayed high levels of proliferation, as shown by accumulation of ascites cells in the abdominal cavities of inoculated SCID animals and the high levels of organ infiltration, as shown by the development of splenomegaly. The pathology of disease observed in inoculated SCID animals closely resembles that seen in the original tax-transgenic animals, 26 with features that are almost identical to those of human ATL. Our analyses of cytokine and growth-factors expression in inoculated animals show that of the panel of molecules analyzed, TNF-α, PDGF-BB, sICAM-1, and sVCAM-1, were the only factors that were produced in significantly high amounts and suggest that these factors may contribute to disease development in this animal model. Levels of IL-1β, IL-2, IL-6, IL-15, IFN-γ, bFGF, MMP9, VEGF, and E-selectin were not significantly different in inoculated and control animals.
The abnormal expression of several cytokines and growth factors is speculated to contribute to the pathogenesis of ATL and HAM/TSP in humans as well as disease development in tax-transgenic animals. Tumor cells from tax-transgenic mice in which the expression of tax is restricted to mature T-lymphocytes/NK cells by using the granzyme B (GzmB) promoter, produce elevated levels of IL-1α, IL-1β, IL-6, IL-10, IL-15, IFN-γ, GM-CSF, ICAM-1, VCAM-1, which was linked to the pathogenesis of disease in that model. 25,32 However, in contrast to our study, GzmB tax animals did not develop the characteristic T-cell lymphoma and leukemia associated with ATL, but instead developed tumors at various peripheral sites and osteolytic bone lesions. In common with that study, we found that IL-2 was not produced in vivo in inoculated animals and showed that the tax-transgenic cell line used in the present study expressed high levels of surface ICAM-1. However, our data show that significant levels of IL-1β, IL-6, IL-15, IFN-γ, or GM-CSF were not produced in inoculated SCID animals and, as such, are unlikely to have contributed to the pathogenesis of disease. These data suggest that the different pathologies observed in both these animal models, together with the different growth-factor expression patterns, specifically relate to the unique characteristics of the different tax-transgenic cells used in each study.
TNF-α was the only cytokine that was significantly produced by these cells in vivo. Elevated TNF-α expression is speculated to contribute to the pathogenesis of HAM/TSP by enhancing the transmigration of HTLV-1–infected cells into the central nervous system. 18,34 Our data also show that, of the growth factors analyzed, PDGF-BB was the only one present at significantly higher levels in the serum of inoculated animals compared with controls. Even though the role of PDGF-BB in the pathogenesis of ATL is unclear, previous studies have shown HTLV-1–infected cells express soluble PDGF and the PDGF receptor, PDGFR-β. 21,35,36 We found that PDGFR-β was not expressed on the cells used in the present study, suggesting that the biologic effects of PDGF may be indirect.
It is well established that Tax 1 modulates several signaling pathways, such as the NF-κB and Akt pathways, altering cellular gene expression. 12 Analysis of Tax 1 expression in the cells used in the present study showed low levels of expression at the mRNA level and undetectable levels of Tax expression at the protein level. This is consistent with studies that show that Tax 1 expression in ATL cells can be extremely low or, in some cases, absent, because of genetic changes in the viral genome. 37,38 These studies suggest that even though Tax 1 is required for the transformation of T cells, it may be dispensible for the maintenance of transformed cells at later stages in the disease. However, irrespective of Tax expression, ATL cells exhibit constitutive NF-κB activity. 39 Our data show that the NF-κB pathway was constitutively active in the tax-transgenic cells used in the present study, with higher levels of activity being observed in the ascites cells. It is interesting to note that this coincided with enhanced levels of both TNF-α and PDGF (three- and ninefold, respectively) in SCID mice inoculated with ascites cells compared with animals inoculated with the cell line. This suggests a connection between the elevated levels of TNF-α and PDGF-BB observed in the serum of animals inoculated with ascites cells and the elevated levels of NF-κB activity observed in these cells. In addition, we showed that the Akt pathway was activated in the tax-transgenic cell line used to inoculate animals. Tax 1 was shown in previous studies to activate Akt by interacting with and activating its upstream phosphatidylinositol-3-kinase (PI3K). 15 Several studies have shown that Akt is activated in HTLV-1–transformed cells and is linked to NF-κB activation, p53 inhibition, and cell survival. 16 Even though the mechanisms involved in promoting the elevated expression of TNF-α and PDGF-BB in inoculated animals in the present study have not been determined, it may be linked to the activated state of both the NF-κB and Akt pathways. Previous studies have shown that TNF-α is an NF-κB–inducible gene, 40 and PDGF-BB expression can be induced by TNF-α. 41 In addition, TNF-α and PDGF-BB have both been previously shown to activate the NF-κB pathway through the Akt pathway, 42,43 even though other studies have suggested that this may occur in a cell-specific manner. 44 Overall, it is possible that a positive-feedback mechanism exists in these cells, whereby NF-κB promotes the expression of TNF-α, which in turn upregulates PDGF expression, and both TNF-α and PDGF contribute to further activation of the NF-κB pathway. However, this hypothesis requires further investigation.
Our data show that the levels of both sICAM-1 and sVCAM-1 were significantly increased in inoculated mice compared with control animals. Even though the pathologic significance of soluble forms of ICAM-1 and VCAM-1 has not been fully elucidated, elevated levels are linked to the pathogenesis of many diseases, including ATL. Elevated levels of sICAM-1 and sVCAM-1 have been reported in HAM/TSP patients, and levels have been correlated with increased migration of HTLV-infected cells into cerebrospinal fluid. 45 Other studies showed that the levels of sICAM-1 correlated with the percentages of CD3+CD25+ cells present in the bronchoalveolar lavage (BAL) fluid of HTLV-1 carriers, suggesting that these molecules may contribute to the pulmonary complications observed in HTLV-1–infected individuals. 46 Based on these findings, it is very likely that these adhesion molecules may facilitate the high rates of migration of cells in inoculated SCID mice. Circulating isoforms of ICAM-1 and VCAM-1 can be released by uncharacterized mechanisms from the surface of cells. 33 Our analyses of the cell-surface expression of both of these adhesion molecules on the tax-transgenic cell line showed that, in contrast to VCAM-1, which was absent, ICAM-1 was highly expressed, raising the possibility that the high levels of soluble ICAM-1 detected in inoculated animals may have arisen because of the shedding of ICAM-1 from the surface of cells.
In conclusion, our study identified TNF-α, PDGF-BB, and the adhesion molecules ICAM-1 and VCAM-1 as factors that may contribute to the pathogenesis of disease in this tax-transgenic SCID model.
Footnotes
Acknowledgments
This project was supported by the National Virus Reference Laboratory, UCD, Ireland.
Author Disclosure Statement
No competing financial interests exist.