
Abstract
The genetic diversity of HIV-1 strains circulating among HIV-1-infected South Africans was investigated in a cohort of 420 individuals enrolled as part of the AIDS Vaccine Integrated Project (AVIP) study. Representative samples (10%) were randomly selected from treatment-naive participants. Viral RNA was extracted for reverse transcriptase-initiated amplification and population-based sequencing of partial pol (encompassing protease and reverse transcriptase) and full-length integrase. Overall, HIV-1 sequences confirmed that 97.1% and 96.9% were HIV-1 subtype C in pol and integrase, respectively. Two participants were infected with unique A1/C and C/A1 recombinants in pol/integrase. Further pol sequence analysis identified mutation patterns associated with high level resistance to NNRTIs in two participants, whereas no primary mutations conferring resistance to integrase inhibitors were detected. The predominance of HIV-1 subtype C in South African populations is therefore confirmed in the AVIP cohort finalized for testing preventive or therapeutic vaccines against HIV-1 infection.
C
Although the constraints placed by ongoing HIV-1 genetic variation on vaccine efficacy remain unclear, it is highly unlikely that incorporation of proteins or DNA derived from a single natural viral isolate into a vaccine will confer protection against the same or other subtypes. Numerous strategies are being investigated to overcome this central problem, including the use of consensus/ancestral sequences, a combination of immunogens from different subtypes, and computationally optimized mosaic immunogens.
HIV-1 group M subtype C accounts for over 50% of existing HIV-1 infections and 47% of new annual HIV-1 infections worldwide.
3
Furthermore, it is the predominant circulating subtype among the heterosexual population in South Africa,
4
the country with the highest number of HIV-1-infected individuals (
Whole blood samples were collected between 2004 and 2007 from 420 HIV-1-positive antiretroviral (ARV) drug-naive individuals attending the Perinatal HIV Research Unit (PHRU) at Chris Hani Baragwaneth Hospital in Johannesburg, South Africa. Plasma and peripheral blood mononuclear cells (PBMCs) were isolated and stored at −80°C and liquid nitrogen, respectively, for each participant, using standard methodologies. Ethical clearance for the study was obtained for Research on Human Subjects (Medical) at the University of the Witwatersrand.
Ten percent of all plasma samples were randomly selected for viremia determination from treatment-naive participants with CD4 >400 cells/μl (n = 21) and CD4 <400 cells/μl (n = 21). Viral RNA was extracted from clarified plasma samples using the automated Roche MagNa Pure LC analyzer and the MagNA Pure LC Total Nucleic Acid Isolation Kit (Roche, Germany), according to the manufacturer's instructions. Extracted viral RNA was used to reverse transcriptase polymerase chain reaction (RT-PCR) amplify the partial pol (encompassing approximately 1.4 kb of the protease and reverse transcriptase) and full-length integrase, using in-house assays. 5,6 PCR amplicons were purified using the High Pure PCR Product Purification kit (Roche), as per the manufacturer's instructions. Primers spanning the partial pol and integrase were used for bidirectional sequencing of the amplicons on the ABI Prism 3100-Avant Genetic Analyzer (Applied Biosystems, Foster City, CA) at a population-based level. Sequencing results were edited using the Sequencing Analysis V3.3 program (Applied Biosystems), and the complete sequences were assembled and manually edited using Sequencher V4.7 (Genecodes, Ann Arbor, MI).
A multiple alignment of the regions of interest with references from HIV-1 subtypes A to K, CRF01_AE, and CRF02_AG (
Overall, HIV-1 sequences were obtained from 35 of the 42 participants (83% success rate). However, matched partial pol and integrase HIV-1 sequences were obtained from 32 participants, including 17 individuals with CD4 counts >400 cells/μl and 15 subjects with CD counts <400 cells/μl. Epidemiological, clinical, and subtyping data analysis of the 35 participants is summarized in Table 1. Seventy-five percent of the participants were female, with a median age of 33 years, and median viral loads of 21,700 RNA copies/ml (ranges 645 to >1,000,000 RNA copies/ml). Phylogenetic tree analysis of the partial pol (Fig. 1A) and integrase (Fig. 1B) sequences with viruses from the major subtypes showed that 30 of the 32 were HIV-1 subtype C.
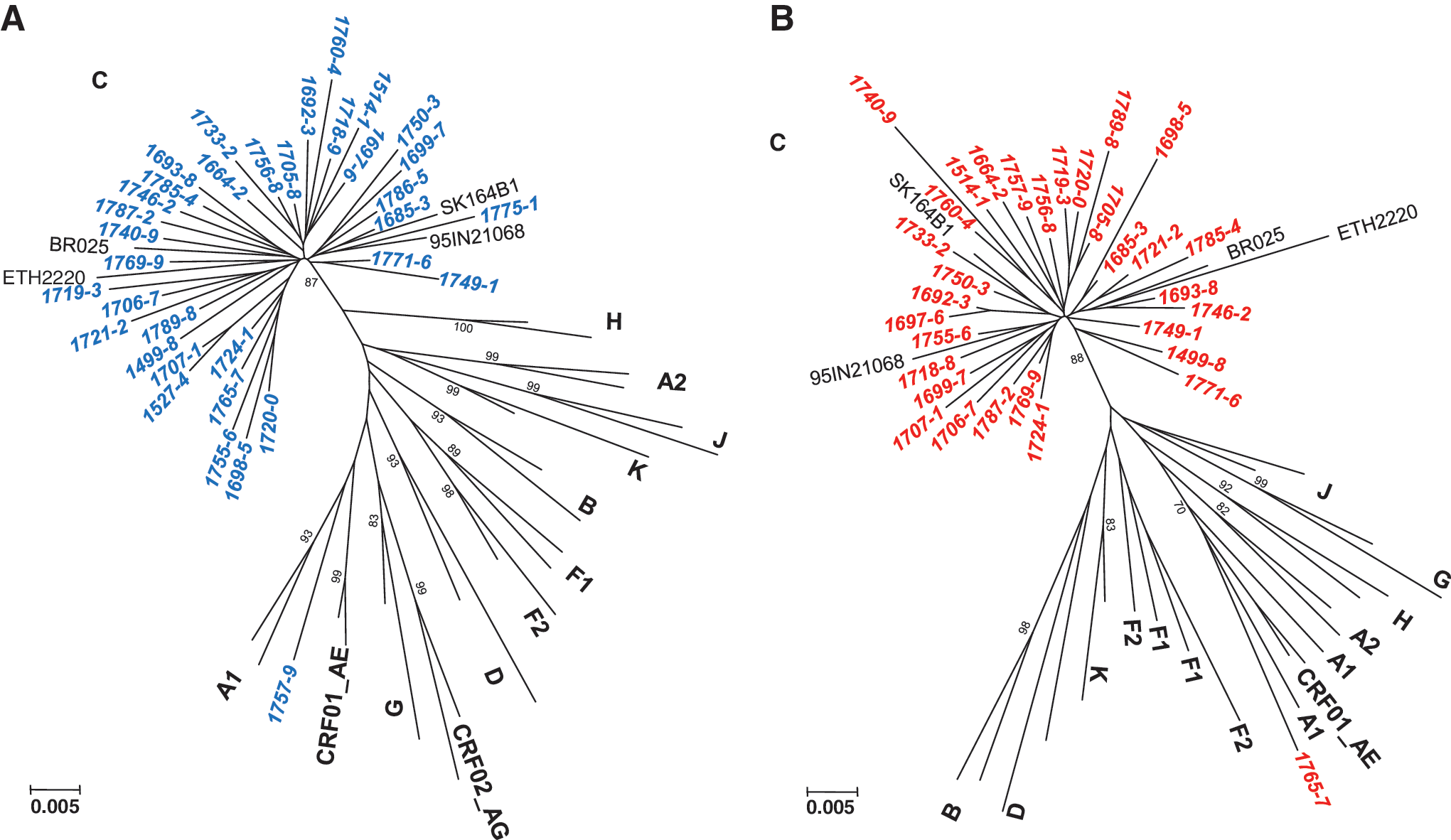
Phylogenetic relationships of the newly characterized partial pol (protease and reverse transcriptase) sequences (
M, male; F, female.
Partial pol/integrase, based on Rega HIV-1 subtyping tool (
For nucleoside reverse transcriptase inhibitors (NRTI), nonnucleoside reverse transcriptase inhibitors (NNRTI), and protease inhibitors (PI).
Integrase inhibitors (
Polymorphic mutations associated with raltegravir resistance.
Not available.
The remaining two participants were infected with unique intersubtype recombinant strains: virus 07ZA17579 was subsubtype A1 and C in the pol and integrase regions, respectively, whereas virus 07ZA17657 was subtype C and subsubtype A1 in the partial pol and integrase regions, respectively (Fig. 1A and B). These recombinant strains combine segments of subsubtype A1 that are typically found in East and Central Africa with subtype C, which is more common in Southern Africa. 3,6 Thus, they could have originated either in East or Central Africa, or they could represent recombination events that occurred between East or Central African and South African strains. The remainder three samples (07ZA17751, 07ZA17865, and 06ZA15274) could not be amplified in the integrase region and were designated HIV-1 subtype C by analysis of their partial pol regions. Since participants 17751, 17865, and 15274 had viral loads of 1140, 1300, and 1190 RNA copies/ml, respectively, it is conceivable that the in-house integrase assay has a different lower limit of detection than the in-house partial pol assay. Alternatively, primer mismatches could account for the lack of amplification. Furthermore, six of the seven samples that could not be amplified in pol and integrase were from participants with low/undetectable viral loads (viral load ranges <400 to 1450 RNA copies/ml).
Analysis of antiretroviral drug resistant mutations among the therapy-naive study participants showed that most of them had viruses susceptible to all known reverse transcriptase and protease inhibitors (n = 33/35, Table 1). However, mutation patterns associated with high level resistance to nonnucleoside reverse transcriptase inhibitors (NNRTIs) were identified in two (5.7%) participants (17657 and 16923). It is possible that participants 17657 and 16923 may have taken antiretroviral drugs previously, and not disclosed this to the study coordinator. Alternatively, they could have been infected with antiretroviral drug resistant virus, or participant 16923, who is a female of childbearing age, may have taken nevirapine as part of the national PMTCT (prevention of mother-to-child transmission) program in South Africa. Participant 17193 contained K103R, a mutation known to occur in 1–2% of untreated individuals. Because K103R was not found in combination with V179D (which reduces nevirapine, delavirdine, and efavirenz susceptibility by about 15-fold), this mutation has no effect on NNRTI susceptibility. 7 In addition, the presence of V118I, a subtype C-specific polymorphism associated with NRTIs, was detected in participants 17503 and 17898. V118I is found in approximately 2% of therapy-naive individuals. It causes low-level resistance to lamivudine (3TC) and possibly to other NRTIs when present with E44A/D and/or one or more thymidine analogue mutations. 8 However, these associated mutations were not found in these participants. Furthermore, participant 17503 had the E138A mutation, a polymorphism recently associated with decreased responses to the NNRTI inhibitor etravirine. 9
The presence of subtype C-specific polymorphisms and minor resistance mutations to protease inhibitors was detected among 11 other participants. Seven participants (17332, 17462, 17699, 17716, 17872, 16938, and 16997) had the T74S mutation that has been described to confer potential low level resistance to nelfinavir. 10 However, this is a naturally occurring subtype C-specific polymorphism, and was thus not listed in Table 1 (as per WHO recommendations). Of the remaining four participants, 17491, 17071, 17212, and 16923 had the Q58E, L10V and V11I, N88K, and A71T, respectively (Table 1). Q58E is a major tipranavir resistance mutation associated with decreased susceptibility to tipranavir/ritonavir and possibly other protease inhibitors. 11 L10V is a minor tipranavir/ritonavir resistance mutation, resulting in resistance to most protease inhibitors when present with other mutations. 11 V11I is a protease inhibitor-selected mutation associated with decreased response to darunavir/ritonavir. 12 The presence of N88D/S/T/G has diverse effects on several protease inhibitors. 13 Finally, the A71T/V polymorphisms are associated with protease inhibitor-based treatment. 14
As expected, none of the 32 participants had primary mutations associated with resistance to the integrase inhibitors raltegravir and elvitegravir (Table 1). Samples 17579 and 16938 contained the G163R and V151I polymorphic mutations, respectively, that have been described with raltegravir use. 15 Sequencing results from integrase obtained in this study confirms previous findings from our laboratory that no baseline polymorphisms affecting the susceptibility to raltegravir could be detected in therapy-naive individuals, and therefore, it can be considered for use in the HIV-1-infected South African population. 5
The overall predominance of HIV-1 subtype C in the infected South African population is confirmed in our AIVP cohort. However, the presence of unique recombinant forms, together with a small, but significant proportion (5.7%) of participants harboring antiretroviral drug-resistant virus with primary mutations, highlights the need for ongoing surveillance of circulating HIV-1 isolates within South Africa,. This is critical to ensure that any vaccine efficacy is not adversely affected by the continuous dynamic evolution of HIV-1.
Sequence Data
All sequences were submitted to GenBank using Sequin V9.2 (
Footnotes
Acknowledgments
The AIDS Vaccine Integrated Project is funded by the FP6 project of the European Commission.
Author Disclosure Statement
No competing financial interests exist.