
Abstract
Human endogenous retroviruses (HERVs) accounting for 9% of the human genome are considered as surrogate markers for genetic instability and as a driving force of genetic variation. Moreover, they modulate regular gene activities and give rise to expression of disease-associated peptides that may serve as diagnostic markers or even targets for T cell-based immune responses. To date, no data are available on the potential link between urothelial carcinogenesis, HERV activity, and tobacco smoking, the main risk for bladder cancer. Here, we report on potential alterations in HERV transcription induced by smoking in a newly established in vitro model and in human urothelium. Normal human dermal fibroblasts were cultivated with urine from never (n = 6) and current smokers (n = 6) and transcription levels for the HERV subfamilies HERV-E 4-1, HERV-T S71-TK1, and HERV-K HML-6 were measured by quantitative real-time PCR. Tendencies toward increased mean transcript levels were detected for cells treated with urine from current smokers. Equally, activity measured in human urothelium supported an increase of HERV transcription in current smokers (n = 9) compared to never smokers (n = 4).
Introduction
T
Even less is known about the influence of smoking on human endogenous retroviruses (HERVs) in urothelial carcinogenesis. HERVs constitute approximately 9% of the human genome, 7,8 and are considered remnants of ancestral infections with exogenous retroviruses that incorporate into germ-line DNA and are transmitted in Mendelian fashion. 9,10 Due to evolutionary selection, deletion, and mutation most of the at least 300,000 HERVs were thought to be inactive. Recent findings, however, demonstrate transcriptional and even translational HERV activity and indicate that HERVs function in the human genomic regulatory system: HERVs contain more than 500,000 promoter elements. At least half of them contribute to cellular transcription activity 11 and about 100 cellular genes are exclusively initiated by HERV-derived promoter elements. 12 Furthermore, they are capable of initiating antisense and interfering noncoding RNA as potential transcriptional regulators. 12,13 In many tumors altered HERV transcription was found. 14 –16
HERVs are also discussed as therapeutic and immunologic targets. In 2008, Takahashi et al. demonstrated regression of renal cell carcinoma induced by cytotoxic T cells targeting a tumor-associated HERV-E antigen. 17 Several in vitro cell culture models were established to better understand the regulation of HERVs and their function in the human genome. In cell culture studies HERV transcription was shown to be upregulated after infection with Toxoplasma gondii and with HHV-6. 18,19 Smoking was previously associated with the activation of transposable elements (TE), which include HERVs, by global hypomethylation. 20,21
The present study was conducted to investigate the influence of smoke metabolites in urine on HERV transcription activity in epithelial cells in vitro and in vivo. We established a new in vitro cell culture model with normal human dermal fibroblasts (NHDFs). NHDFs are a well-studied model for benign endothelial cells and are sensitive to external influences. 22 They were cultured with urine from never and current smokers (NS, CS) and transcriptional activity for the three HERV subfamilies HERV-E 4-1, HERV-T S71-TK1, and HERV-K HML-6 was measured by quantitative real-time polymerase chain reaction (QRT-PCR). In a parallel approach, we analyzed for the first time transcriptional activity in benign human urothelium of CS and NS. Concurring tendencies in epithelial cells from in vitro and in vivo material display for the first time a potential influence of smoking on HERV expression.
Materials and Methods
NHDF cultured with urine from CS and NS
A total of 5 × 105 NHDF cells (Promocell GmbH, Heidelberg, Germany) were cultured in 10 cm culture plates and grown in RPMI 1640 medium (Invitrogen, Karlsruhe, Germany) supplemented with glutamine, 10% fetal calf serum (Invitrogen), and 1% streptomycin/penicillin (100 U/ml; Invitrogen) under standard conditions at 37°C and 5% CO2. Urine was collected from healthy probands who reported a smoking history of either more than 15 pack years (CS) or as never smokers (NS). Urine was measured for pH [CS, 6.2 (5.4–6.6); NS, 6.2 (5.3–6.8)], filtered for sterilization, and stored at −20°C until usage.
In a pilot study cell growth with urine concentrations in the medium of 0 (control), 5, 10, 15, 20, 30, 40, and 50% urine was tested. pH of the urine samples between 5.3 and 6.8 exhibited no influence on cell growth and pH adjustment was omitted.
For the final experiments NHDFs were treated for 2 weeks with urine from CS and NS. Twice a week culture medium was renewed and urine was added in the corresponding concentration. Subconfluent cells were split weekly. Duplicate experiments were repeated three times (CS, n = 6; NS, n = 6).
Human urothelial tissue samples
Benign urothelium was prospectively obtained from patients treated at the Department of Urology of the Mannheim Medical Center. A positive ethic vote for tissue collection and data acquisition for further investigations from patients being treated at our department was obtained by the Ethic Committee II, Faculty of Medicine, University of Heidelberg (number of request: 2007-030N-MA). Patients undergoing routine surgery for urothelial carcinoma (UC) were informed about the study. If they agreed to participate, tissues were collected from routine surgery and immediately placed in RNAlater (Qiagen, Hilden, Germany). Tissues were stored in RNAlater at 4°C overnight and were then snap frozen in liquid nitrogen and stored at −80°C until further processing. Risk factors, including smoking habits, were assessed by a standardized questionnaire for each patient included. A group of 5 female and 10 male white patients [mean age: 66 (51–88)] was divided into study groups of CS (>5 years of smoking, n = 9) and NS (n = 4).
RNA preparation
Frozen tissue fragments and cells harvested from tissue cultures were processed using Qiagen RNeasy MiniKit (Qiagen). Tissue fragments were mechanically disrupted in the TissueLyser (Qiagen) in RLT buffer. Cells from cell cultures were lysed in RLT buffer by passing six times through a 20-gauge needle using a 2-ml syringe. Further procedures followed the manufacturer's instructions. RNA processing was conducted as described previously. 23 In brief, RNA was treated with DNase, subsequently controlled for the absence of genomic DNA, and reverse transcribed into cDNA.
Quantification of HERV transcripts by QRT-PCR
For the amplification primers for subgroup-specific pol regions were used for HERV-T S71-TK1 (forward primer, 5′-gtaccccaggtaggaaactctggg-3, and reverse primer, 5′-cccctaccctttttgggg-3′) and HERV-E 4-1. 23 Since it was not possible to obtain subgroup-specific primers for the HERV-K HML-6 family, previously published degenerate primers specific for the HERV-K HML-6 family were employed. 24 For normalization, the housekeeping gene glycerinaldehyde-3-phosphate dehydrogenase (GAPDH) (forward primer, 5′-agtcaacggatttggtcgtattggg-3′, and reverse primer, 5′-acgtactcagcgccagcatcg-3′) was amplified. As reference sequences, standard plasmids containing cloned fragments of HERV-T S71-TK1 (accession no. U12970), HERV-E 4-1 (accession no. M10976), and HERV-K HML-6 (accession no. AF079797) as well as plasmids containing GAPDH (accession no. NM002046.3) gene fragments were used.
QRT-PCR measurements were preformed with the LightCycler 2.1 (Roche Diagnostics GmbH, Mannheim, Germany). Two μL of cDNA was added to 18 μl of reaction mix, comprising primers at 0.5 μM for the HERV targets and at 0.25 μM for GAPDH in LightCycler FastStart DNA MasterPLUS SYBR Green I ready-to-use hot-start PCR mix with Taq DNA polymerase (Roche Diagnostics GmbH) diluted with purified water according to the manufacturer's protocol. Amplification was performed using a 10 min denaturation step at 95°C, followed by 45 cycles of 1 s at 95°C, 10 s at 54°C (for HERV-targets), or 60°C (for GAPDH), and 12 s at 72°C. Melting curve analysis was performed and validated for each run to identify amplicons and exclude impurities (data not shown). Relative quantification was calculated using LightCycler software (version 3.5, Roche Molecular Biochemicals). Interexperimental validation was performed to ensure the reproducibility of the obtained data. Values were normalized for the housekeeping gene, thus expression levels were stated as the quotient of the targeted HERV divided by GAPDH. GAPDH proved to be stably expressed in benign urothelium of CS and NS as determined by microarray analysis in our laboratory (Gabriel et al., unpublished observations). Due to the low transcription levels of HERV targets, not all probes reached the lowest detection limit. Probes with expression levels lower than the standard curve were excluded from further calculations.
Statistics
Statistical analysis was performed with SPSS 15.0. The approximate normal distribution of parametric results was confirmed by linearity in a Q-Q plot. Differences between the study groups were calculated by Student's t test. A p-value of <0.05 was considered significant.
Results
In vitro cell culture model with urine from CS and NS
In the present study, we established a new in vitro cell culture model to detect changes in HERV transcription induced by smoking. We therefore cultured NHDF cells with urine from CS and NS. In our pilot study, we verified cell growth in culture conditions of up to 50% urine in the culture medium. With increasing urine, supplement constrained cell growth was observed: cells cultured in urine from CS grew up to 30% urine supplement whereas cells cultured in urine from NS sustained viability up to 50%. Urine concentrations of 10% and 20% showed sufficient cell growth in all set-ups and were chosen for the final experiments. Cell growth was considered stable to varying pH of the urine samples because no differences in times with regard to subconfluence and splitting were observed. Throughout the 2 weeks of urine treatment, no morphologic changes occurred in the microscopic survey. The NHDF cell line was therefore considered stable and consistent growing in the urine-supplemented medium.
HERV transcription and smoking
Quantitative analysis of HERV activity suggested concurrent induction of HERV transcription by smoking in the new in vitro cell culture model and in benign urothelium.
HERV transcription in NHDF cells treated with urine from CS and NS
NHDF cells cultured with urine from CS revealed elevated mean relative transcription levels of HERV-E 4-1, HERV-K HML-6, and HERV-T S71-TK1 compared to cells cultured in urine from NS (Fig. 1A). HERV-K HML-6 (NS = 3, CS = 4) and HERV-T S71-TK1 (NS = 3, CS = 5) mean expression levels tended to be strongly increased by 626% and 376%, respectively. HERV-E 4-1 (NS = 4, CS = 6) mean activity suggests a minor elevation of 1.3% (Table 1).
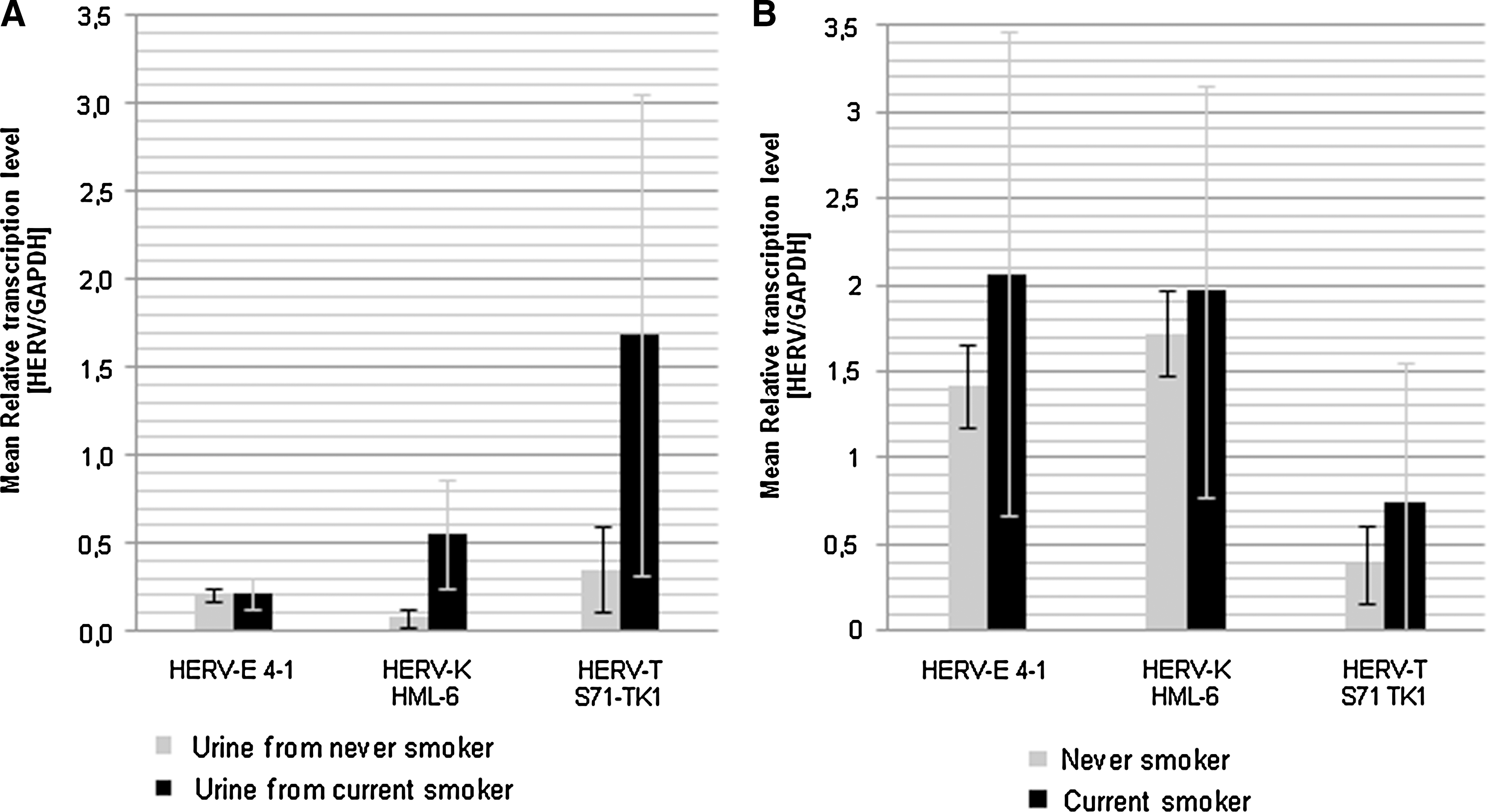
Mean relative transcriptional activity of the HERV subfamilies HERV-E 4-1, HERV-K HML-6, and HERV-T S71-TK1 was measured by QRT-PCR. Transcription values were normalized for GAPDH. Standard deviations are marked by error bars. (
NS, urine from never smokers; CS, urine from current smokers.
Numbers reflect the difference of CS to NS relative to NS.
HERV transcription in normal human urothelium
In the benign urothelium of CS and NS relative transcription levels of HERV-T S71-TK1 are approximately three-fold lower than that of HERV-E 4-1 and HERV-K HML-6. In all three HERV subgroups, CS revealed elevated mean relative transcription levels compared to NS (Fig. 1B). HERV-E 4-1 (NS = 4, CS = 8) and HERV-T S71-TK1 (NS = 3, CS = 8) showed major increases of 46% and 93%, respectively. HERV-K HML-6 (NS = 3, CS = 8) expression was slightly increased by 14% (Table 2).
NS, urothelium from never smokers; CS, urothelium from current smokers.
Numbers reflect the difference of CS to NS relative to NS.
Large differences observed, most notably in cell lines, very likely resulted from measuring close to the lower working range limit, where the measuring error is largest. Our preliminary results were not significant but might suggest a trend toward increased transcription by smoking.
Discussion
In the present study, we established a new in vitro cell culture model (NHDF cells) and results suggested upregulation of HERV transcription by urine from CS. In parallel, HERV transcriptional activity was measured for the first time in benign human urothelium of CS and NS and results concurred with the tendencies seen in the in vitro model of elevated transcription levels for CS.
NHDF cells are well established for the detection of genetic instability. 22,25,26 The primary endothelial cell line is also susceptible to changes in transcriptional activity by external influence, 27 and is likely to better respond to urine treatment than urinary epithelial cell lines since it is naturally not subject to urine contact. Despite changes in transcription NHDFs were considered stable and consistent growing up to 20% of the urine supplement. NHDF cells are therefore suitable as a model for transcriptional activity under certain urinary bladder conditions.
Cigarette smoke contains to date 81 carcinogens as defined by the IARC. 28 In most cases, they undergo metabolism and activation and accumulate in the urine as reactive molecules. 29,30 Carcinogen levels in urine can vary due to individual polymorphisms and cigarette consumption. 3,31,32 Nevertheless, urine was chosen to best imitate urinary bladder conditions. It takes into account the wide diversity of smoke carcinogens and the metabolic process. More precise set-ups with regard to quantification are needed, as the in vitro composition of known major carcinogenic urinary metabolites neglects the wide range of minor carcinogens. In addition, induction of cigarette smoke extract to the medium as previously described 33,34 would lack the metabolic activation of the smoke carcinogens and is therefore inappropriate.
In cell models, HERV transcription has previously been shown to be amplifiable by chemicals or infections. 18,19,35 Here the application of urine from CS showed a trend to induced transcriptional activity of HERV-E 4-1, HERV-T S71-TK1, and HERV-K HML-6 in the NHDF cells compared to the application of urine from NS. In a parallel approach, we measured for the first time the transcriptional activity of the three HERV subgroups in human urothelium obtained from CS and NS. Equally elevated transcription levels suggested that dermal epithelial cells in urine culture are a suitable model for HERV transcription in human urothelial cells.
To date, the exact role of HERVs in human gene expression is broadly discussed but remains uncertain. Furthermore, no direct function can be assigned to the three selected targets. HERV-E and HERV-K HML-6 were previously associated with carcinogenesis. 16,17,19 In cell culture models, HERV-E also showed transcriptional upregulation after infection with HHV-6 and Toxoplasma gondii. 18,19 HERV-K HML-6 is expressed in various normal tissues and showed significantly lower transcription in human breast cancer. 16,23 HERV-T has been identified as active almost exclusively in benign genitourinary tissues, such as uterus, cervix, testis, and placenta. 23 In fact, the three HERV subgroups can be regarded as exemplary and investigations in other HERV families will have to follow.
The results indicate that smoking and its metabolites may regulate transcriptional activity of HERVs. HERVs are postulated to affect the entire gene expression by several regulatory mechanisms including their ability to enhance gene transcription, provide alternative promoters, disrupt gene exon-intron structure, cause premature transcriptional termination, and generate antisense and interfering noncoding RNA. 12,13,36,37 Potential increases in HERV activity by smoking may therefore be associated with aberrant cellular transcriptional activity.
Because smoking is the main risk factor for developing urothelial carcinoma, we conclude that the suggested upregulation of HERV transcription by urinary smoke metabolites may induce or reflect the overall cellular transcriptional changes that lead to or are part of precancerous cellular activity. If the tendencies shown in this study can be confirmed in consecutive studies, HERVs might serve as clinical markers in the future. For urothelial carcinoma, clinical markers are urgently needed, since high rates of recurrence and progressiveness make it a life-threatening disease. 38
Our study suggested for the first time that expression of HERVs is affected by smoking. The strengths lie in the novel approach and the concordant tendencies found in vitro and in vivo. Limitations were the pilot character of the study with a small sample size and difficulties in quantification due to low transcription of HERVs in the samples, and the lack of significant results. The results, however, have encouraged us to repeat experiments with larger study groups and additional HERV targets. It will be of major interest to better understand the functions of HERVs and determine their underlying mechanisms. Then upregulation of HERV transcription might serve in clinical practice as new markers for carcinogenesis, as an estimation of the risk for smokers of developing urothelial carcinoma, and even as therapeutic targets.
Gene Name and Accession Numbers
Glycerinaldehyde-3-phosphate dehydrogenase (GAPDH), NM 002046.3; HERV-E 4-1, M10976; HERV-K HML-6, AF079797; HERV-T S71-TK1, U12970.
Footnotes
Acknowledgments
We are grateful to all colleagues and nursing staff from the Department of Urology for their great support and cooperation. We also greatly appreciate the help and technical assistance of our colleagues at the laboratory of the Medical Clinic III.
Author Disclosure Statement
No competing financial interests exist.