
Abstract
To elucidate the immunological role for the costimulatory molecule OX40 against the early stage of HIV-1 infection, fresh peripheral blood mononuclear cells (PBMCs) from normal donors were stimulated with immobilized anti-CD3 monoclonal antibody (mAb) together with soluble anti-CD28 mAb for 24 h, infected with CCR5-tropic (R5) HIV-1, and then cocultured in the presence or absence of OX40 ligand (OX40L). Results of these studied showed that OX40 stimulation led to a marked reduction in levels of p24, the frequency of intracellular p24+ cells, as well as HIV-1-mediated syncytium formation. The suppression was reversed by anti-OX40L mAb. The mechanism underlying the R5 HIV-1 suppression was shown to be mediated in part by the CCR5-binding β-chemokines RANTES, MIP-1α, and MIP-1β, since the effect of the OX40 stimulation was reversed by a neutralizing antibody mixture against these three β-chemokines. Thus, OX40 stimulation enhanced the production of these CCR5-binding β-chemokines by the activated PBMCs and subsequently down-modulated CCR5 expression on the activated CD4+ T cells. Taken together, the present data revealed a new role for OX40 in HIV-1 infection and documents the fact that OX40 stimulation suppresses the infection of primary activated PBMCs with R5 HIV-1 via enhanced production of R5 HIV-1 suppressing β-chemokines.
Introduction
O
It is not clear at present how OX40-mediated costimulation affects HIV-1 infection. Based on cell tropism, human immunodeficiency virus type 1 (HIV-1) isolates are classified into two main groups, CCR5-tropic (R5) and CXCR4-tropic (X4) strains. 26 The predominant HIV-1 strains isolated during acute infection are R5 tropic and often evolve to more pathogenic X4 strains as the disease progresses. 27 Despite these differences in tropism, both R5 and X4 HIV-1 strains are able to productively infect primary activated human CD4+ T cells. We have previously shown that OX40 stimulation enhances X4 HIV-1 production in primary activated CD4+ T cells and T cell lines via activation of the NF-κB pathway. 28 These observations prompted us to determine the role of OX40 costimulation on R5 HIV-1 infection. We report herein that in contrast to X4 HIV-1 infection, OX40 stimulation interferes with R5 HIV-1 infection by enhanced production of the R5 HIV-1-inhibiting β-chemokines.
Materials and Methods
Reagents
The medium used consisted of RPMI 1640 medium (Sigma-Aldrich. Inc., St. Louis, MO), supplemented with 10% fetal calf serum (FCS), 100 U/ml of penicillin, and 100 μg/ml of streptomycin (hereinafter called RPMI medium). Antihuman CD3 (clone OKT-3) and anti-CD28 monoclonal antibodies (mAbs) were purchased from the American Type Culture Collection (Rockville, MD) and Biolegend (San Diego, CA), respectively. Neutralizing mAbs against human RANTES, MIP-1α, and MIP-1β were purchased from R&D systems (Minneapolis, MN). The mAbs produced in our laboratory included mouse IgG1 mAbs anti-OX40L (clone 5A8), 29 antihuman OX40 (clone B-7B5), 28 anti-HIV-1 p24 (clones NP24 and 2C2) (Tanaka et al., unpublished), anti-CD25 (clone H-8), 30 and anti-HTLV-I tax (clone TAXY-8) 31 as well as rat IgG2b mAbs anti-OX40 (clone W4-54) 32 and isotype control anti-HCV (clone Mo-8). 33 The in-house mAbs were purified from BALB/c or CB.17-SCID mouse ascites fluids by a gel filtration method. Some of them were labeled using FITC, Cy5, or HRP using commercial labeling kits (Dojin or Amersham, Japan) according to the manufacture's instructions. Biotinylated recombinant soluble human OX40 (sOX40 in a form of murine IgG2a-Fc fusion protein) and OX40L (sOX40L in a form of murine CD8-fusion protein) were purchased from Ancell (Bayport, MN) and used with FITC-streptavidin (Biolegend) for staining. Unlabeled glycosylated recombinant human OX40L, which consists of OX40L and human CD33 signal peptide produced in NS1 cells, was purchased from R&D systems. The mock-transfected CEM cell line (CEM/mock) and CEM cell line transfected with human OX40L (CEM/gp34) and shown to express OX40L 25 were used for some of the studies described below.
HIV-1 preparation
HIV-1JR-FL and HIV-1NL4-3 viral stocks were produced by transfection of the 293T cells with the appropriate HIV-1 infectious plasmid DNAs utilizing the calcium phosphate method followed by in vitro culture of the transfected cells in RPMI medium for 2 and 3 days followed by collection of the supernatant fluid and virus isolation and purification as described previously. 34
Quatitation of HIV-1 antigen and chemokines
Production of HIV-1 was determined by the measurement of HIV-1 core p24 levels produced in the culture supernatants using a commercial ELISA kit (Retrotec, Buffalo, NY) or our in-house formulated and standardized kits using a pair of anti-p24 mAbs of mouse origin with a detection limit of p24 of 2 pg/ml (Tanaka, unpublished). The levels of the human β-chemokines RANTES, MIP-1α, and MIP-1β in the culture supernatants were determined using ELISA kits purchased from Biosource (Camarillo, CA).
Stimulation of PBMCs in vitro and infection with HIV-1
PBMCs from healthy donors were stimulated as described previously. 17 Briefly, PBMCs obtained by density gradient centrifugation on HistoPAQUE-1077 (Sigma-Aldrich) were suspended at 2 × 106 cells/ml in RPMI medium, dispensed into individual wells of 24-well plates (BD) (1 ml/well) precoated with 5 μg/ml OKT3 for 1 h, and cultured in the presence of soluble 0.1 μg/ml anti-CD28 mAb. After cultivation at 37°C in a 5% CO2 humidified atmosphere for 24 h, the activated PBMCs were washed once and infected with either R5 HIV-1JR-FL or X4 HIV-1NL4-3 at a multiplicity of infection (moi) of 0.005 for 2 h as described previously. 34 After washing three times, PBMCs were resuspended at 1 × 106 cells/ml in RPMI medium, dispensed into individual wells of 48-well plates (BD) (0.5 ml/well), and cultured in the presence or absence of OX40L stimulation by either coculture with 2 × 105 cells/ml CEM/gp34 or by the addition of 1 μg/ml recombinant OX40L (rec.gp34). Syncytium formation was observed using an inverted microscope at a magnification of 200 × as described previously. 35 In some experiments, 1-day-activated PBMCs were stimulated with 1 μg/ml recombinant OX40L for 24 h to examine the levels of chemokines in the culture supernatants.
Flow cytometry (FCM)
FCM analysis of live cells was carried out as described previously. 32 Briefly, cells to be analyzed were Fc-blocked with 2 mg/ml normal human pooled IgG on ice for 15 min. Aliquots of these cells were then subjected to staining using predetermined optimum concentrations of fluorescent dye-conjugated mAbs for 30 min on ice. The cells were then washed using FACS buffer (PBS containing 2% FCS and 0.1% sodium azide), fixed in 1% paraformaldehyde (PFA) containing FACS buffer, and analyzed using a FACS Calibur, and the data obtained were analyzed using the Cell Quest software (BD). To determine whether cell surface OX40 or OX40L is functional, aliquots of Fc-blocked cells were incubated with either biotinylated recombinant soluble OX40L or soluble OX40 at a concentration of 2.5 μg/ml for 30 min on ice, followed by staining with FITC-labeled streptavidin (Beckman Coulter) for 30 min on ice and then analyzed by FCM. For detection of HIV-1-infected cells, PBMCs were fixed with PBS containing 4% PFA followed by washing twice in FACS buffer containing 0.5% Triton X-100. These cells were Fc-blocked with 2 mg/ml normal human pooled IgG on ice for 15 min, and aliquots of these cells were stained with FITC- or Cy5-conjugated anti-HIV-1 p24 mAb (clone 2C2) for 30 min on ice. The cells were then washed using FACS buffer and absolute cell counts of p24+ cells were performed by FCM using a cell counting kit (BD) according to the manufacturer's protocol.
Statistical analysis
Data were tested for significance using the Student's t test by using Prism software (GraphPad Software).
Results
Functional OX40 expression on CD4+ T cells in primary activated PBMCs
To confirm that primary activated CD4+ T cells express functional OX40 on their cell surface, fresh PBMCs from healthy donors were stimulated in vitro with immobilized anti-CD3 mAb (OKT-3) along with soluble anti-CD28 mAb for 24 h and aliquots were stained with biotinylated recombinant OX40L (gp34) or OX40 followed by FITC-streptavidin or directly conjugated mAbs against OX40 and OX40L. As shown in Fig. 1A, two-thirds of CD4+ T cells were positively stained with recombinant OX40L (gp34), indicating that these CD4+ T cells expressed functional OX40. Similar percentages of CD4+ T cells were also stained with anti-OX40 mAb. In the same culture, there were some CD4− cells that were stained with biotinylated OX40L (6.2% of total PBMCs) and anti-OX40 (9.7% of total PBMCs). These cells mainly consisted of CD8+ T cells (data not shown). In the same activated PBMC culture, OX40L+ cells as demonstrated by OX40 binding and anti-OX40L (gp34) staining were present at a low frequency, 1.1% and 0.6%, respectively. These OX40L+ cells mainly consisted of activated CD4+ and CD8+ T cells, as reported previously, 32 suggesting that OX40-mediated costimulation of T cells may occur via T–T cell interaction initiated during an early period after PBMC activation.

Flow cytomeric analysis of OX40 and OX40L expression by anti-CD3/CD28-activated primary cultures of human PBMCs. (
A kinetic study of OX40 expression by primary activated PBMCs using anti-OX40 mAb (Fig. 1B) shows that the percent positive OX40+CD4+ T cells peaked on day 2, declined on day 3, and was reinduced by secondary stimulation and then gradually declined. After primary stimulation, the mean fluorescence intensity (MFI) of OX40 expression by CD4+ T cells was highest on day 1 (MFI of 174) and declined on day 2 (MFI of 113) and day 3 (MFI of 64). The MFI following secondary stimulation peaked on day 1 and thereafter decreased to a stable level. The mean density of OX40 expression by CD4+ T cells was two to three times higher than that of the non-CD4+ (presumably CD8+ T cells) at any stage of in vitro culture. In contrast, the expression level of CD25 (another activation marker) by the CD4+ and the non-CD4+ T cells was essentially similar throughout the culture period. These results showed that in the present experimental conditions, high levels of functional OX40 can be induced mainly on CD4+ T cells in primary cultures of human PBMCs following activation, and its increased expression was transient as compared with CD25.
Effects of OX40 stimulation on R5 HIV-1 infection
To examine the effect of OX40 stimulation on infection of CCR5-tropic HIV-1, PBMCs activated for 24 h were infected with R5 HIV-1JR-FL at a low moi, and then were cocultured in the presence or absence of OX40L+CEM cells (CEM/gp34) or recombinant soluble OX40L (rec.gp34). The reason for choosing 1-day activated PBMCs as the target cells was because we wanted to study the response of CD4+ T cells that were relatively naive to OX40L stimulation. In addition, we have previously shown that prolonged cultivation of activated PBMCs reduces the sensitivity of these cells to exogenous OX40L stimulation due to presensitization that may occur by endogenous OX40L expressed by T cells via T–T cell interactions. 32 As shown in Fig. 2A (top left), the production of HIV-1JR-FL in the culture supernatants as shown by the levels of p24 was significantly reduced by coculture of activated PBMCs with CEM/gp34 (p < 0.001 on days 4 and 5) but not OX40L−CEM (not shown). This inhibition was OX40 specific since the addition of the OX40L neutralizing mAb (clone 5A8) at coculture initiation reversed the inhibitory effect by OX40L stimulation (p < 0.001 on days 4 and 5). Furthermore, the addition of soluble recombinant OX40L at a concentration of 1 μg/ml also reduced the level of HIV-1 p24 production (p < 0.01 on days 4 and 5). The dose-response of the soluble OX40L is shown in Fig. 2A (top right), showing that the soluble OX40L at a concentration of more than 1 μg/ml suppressed R5 HIV-1 proliferation. It is worth noting that HIV-1JR-FL often induces multinuclear giant cells (syncytia) in primary activated PBMCs, and that such syncytium formation was markedly inhibited by OX40 stimulation (Fig. 2B), suggesting that OX40 stimulation could interrupt the spread of R5 HIV-1 infection by inhibiting cell-to-cell interaction.
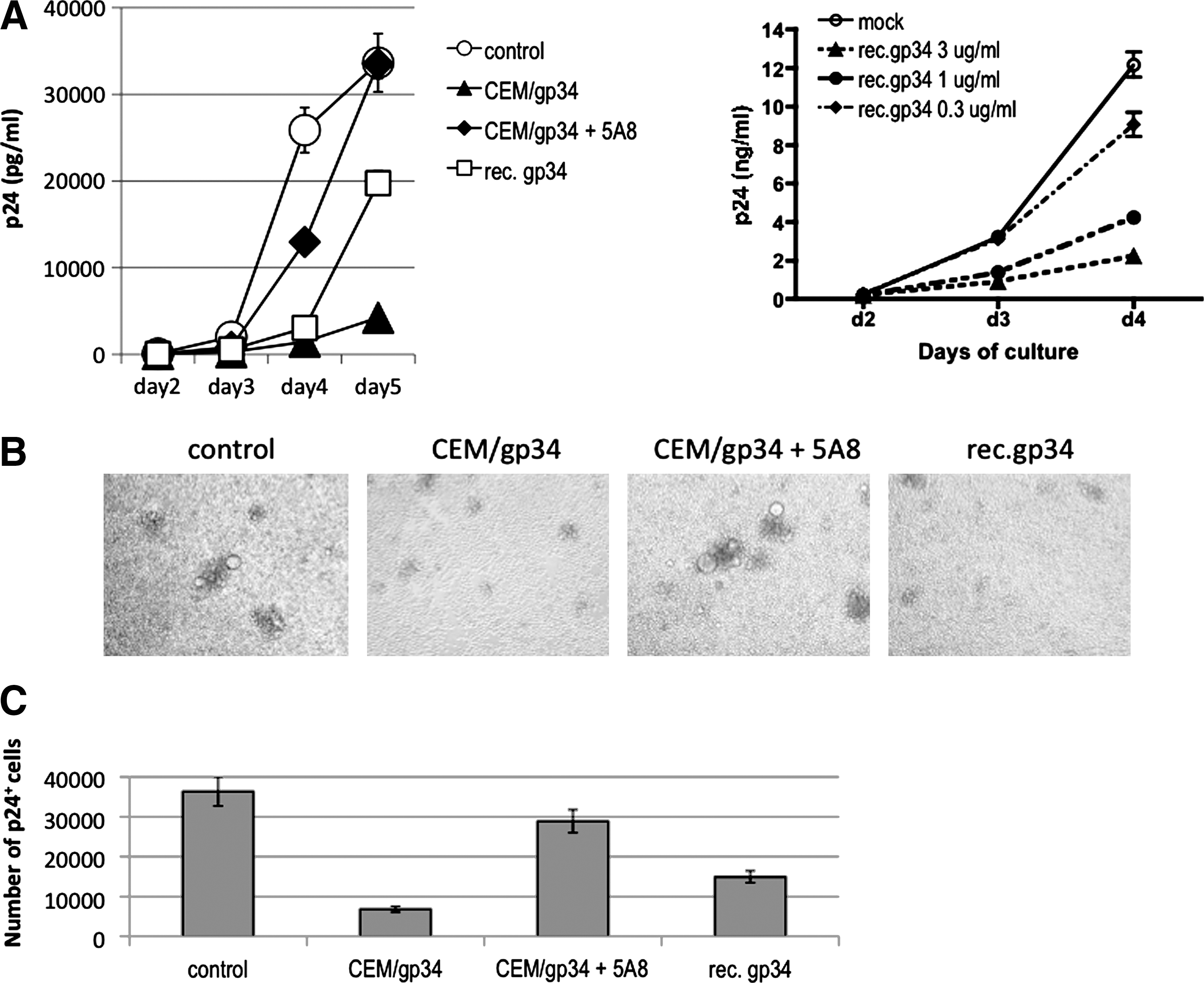
Inhibition of R5 HIV-1 replication by OX40 stimulation in vitro. (
To determine whether the inhibition noted by OX40 stimulation was indeed due to blocking of the spread of HIV-1 to uninfected cells or secondary to inhibition of de novo production of HIV-1 from the infected cells, we examined the cell cultures for the frequency of intracellular HIV-1 core p24 antigen-positive cells in each culture. Figure 2C shows that the HIV-1 p24 Ag+ cell number per culture was significantly lower (p < 0.001) in the OX40-stimulated cultures than the control cultures. Again, the reduction was specifically reversed by the addition of anti-OX40L (gp34) mAb (p < 0.001). Together, these results clearly indicate that OX40 stimulation suppresses R5 HIV-1 infection in primary activated PBMCs
OX40 stimulation does not inhibit infection of X4 tropic HIV-1 in activated PBMCs
Our laboratory previously reported that X4 HIV-1 infection of primary CD4+ T cells activated twice (day 0 and 3) was markedly enhanced by OX40 stimulation. 28 We therefore tested whether OX40 stimulation in the present culture conditions could modify infection of the day-1-activated PBMCs with X4 HIV-1. Thus, 1-day activated PBMCs were infected with X4 HIV-1NL4-3 and then cultured in the presence or absence of OX40 costimulation. As shown in Fig. 3, X4 HIV-1 infection was not blocked (p < 0.001 on days 3–5) or slightly enhanced by stimulation by coculture with OX40L-expressing CD4+ T cells. These data suggest that the effect of OX40 stimulation on HIV-1 infection is distinct depending on the coreceptor tropism of HIV-1 strains.

OX40 stimulation does not inhibit X4 HIV-1 infection of activated PBMCs. Fresh primary anti-CD3/CD28-activated PBMCs were infected with X4 HIV-1NL4-3, as described in Fig. 2, and production of HIV-1 was monitored by p24 ELISA. Data shown are representative of five independent experiments using three different donors.
CCR5-binding β-chemokines are involved in the R5 HIV-1 interference by OX40 stimulation
The present observations that OX40 stimulation interfered with R5 but not X4 HIV-1 infection of activated PBMCs prompted us to examine the factors that could potentially be involved and reasoned that the CCR5 binding β-chemokines were likely candidates. Therefore, studies were carried out using a mixture of neutralizing mAbs against RANTES, MIP-1α, and MIP-1β in efforts to determine if their addition could reverse the R5 HIV-1 inhibiting effect by OX40 stimulation. As shown in Fig. 4A, the OX40-mediated R5 HIV-1 suppression was significantly reversed by the addition of the mixture of antibodies (p < 0.001 on days 4 and 5). Interestingly, use of mAbs against the three individual β-chemokines in similar cultures performed in parallel did not show any detectable reversal of the effect of OX40 stimulation (data not shown), suggesting that all the three chemokines contributed in the suppression of R5 HIV-1 infection. To confirm that these β-chemokines were actually produced in the OX40-stimulated PBMCs, we quantitated the concentrations of the chemokines in the culture supernatants. Figure 4B shows that OX40 stimulation enhanced the production of these β-chemokines in the activated PBMCs (p < 0.001). Furthermore, after OX40 stimulation, the level of CCR5 expression on the cell surface of CD4+ T cells in the activated PBMCs was significantly reduced (Fig. 4C). In contrast, CXCR4 expression by CD4+ T cells was slightly up-regulated in the same OX40-stimulated PBMC culture. Therefore, these results indicate that OX40 stimulation can suppress R5 HIV-1 infection via enhanced production of CCR5-binding β-chemokines. The fact that the inhibition of R5 HIV-1 infection was reversed by the addition of antibodies against the β-chemokines suggests that the inhibition is not secondary to down-regulation of CCR5 but likely was due to the blocking of the CCR5 by the β-chemokines.
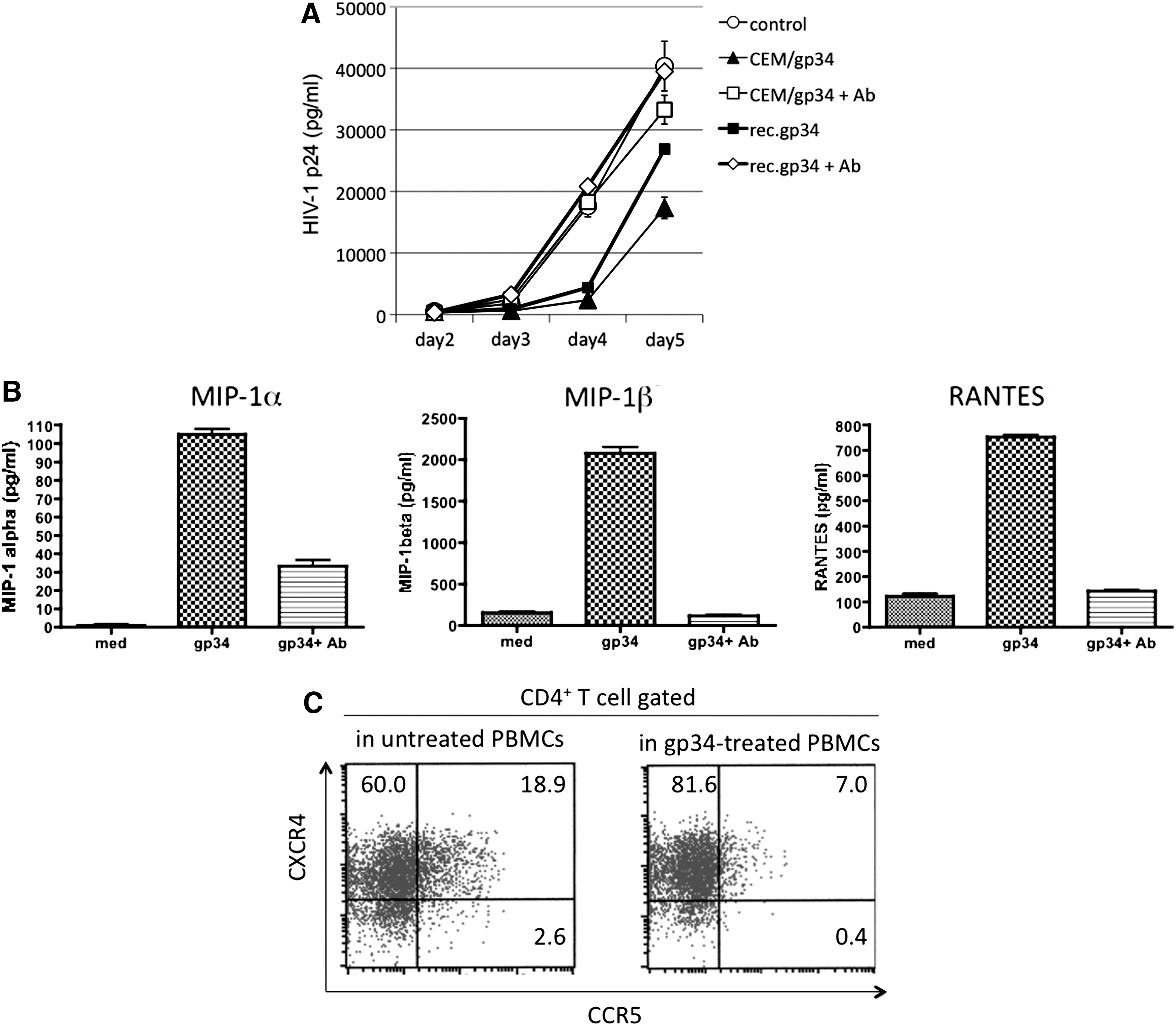
Enhanced production of CCR5-binding β-chemokines is involved in the R5 HIV-1 inhibition by OX40 stimulation. (
Discussion
In the present study, we showed for the first time that costimulation of primary activated PBMCs by OX40L reduced the productive infection with R5 HIV-1. This effect is distinct from the effect of OX40L-induced OX40 stimulation on infection by X4 HIV-1 in which OX40 enhances the infection. 28 The underlying mechanism for R5 HIV-1-restiriced suppression was shown to be enhanced production of the CCR5-binding β-chemokines that led to either blocking and/or down-modulation of CCR5 expression on activated CD4+ T cells.
It has been reported that OX40 stimulation of activated T cells induces enhanced production of a variety of cytokines such as IL-2, IL-4, IL-5, IL-6, IL-8, IL-13, and TNF-α. 20 –23 As far as we know there have been no reports on enhanced CCR5-binding β-chemokine production by activated PBMCs and CCR5 down-modulation in activated CD4+ T cells by OX40 stimulation. The enhancement of each chemokine production increased in the order RANTES (sixfold), MIP-1β (∼15-fold), and MIP-1α (∼50-fold) (Fig. 4B). It is possible that the levels of chemokines quantitated in the culture supernatants were even higher since some of them might have been absorbed by CCR5-expressing cells. All three β-chemokines might be involved in R5 HIV-1 suppression since our preliminary data show that the mixture of all the three mAbs against the β-chemokines, but not single mAb against each chemokine, was required for complete reversal of the OX40-mediated R5 HIV-1 suppression in the present culture conditions (data not shown). Further studies need to be performed to determine whether it is indeed the direct result of CD4+ T cell activation or due to synthesis by the other cell lineages present in the culture conditions employed. Data from our preliminary experiments show that the enhanced production of the three β-chemokines by OX40 stimulation was significantly reduced (>80%) after removal of both CD4+ and CD8+ T cells, and only partially after removal of CD14+ monocytes, but not after removal of CD19+ B cells (data not shown). Thus, it is possible that both CD4+ and CD8+ T cells and to a lesser extent monocytes are the major producers of these β-chemokines. Further studies including analysis of cells stained for cell surface markers coupled with intracellular staining for the chemokines in question are currently in progress.
Several lines of evidence suggest that targeting CCR5 by small chemical compounds, modified RANTES, or MIP-β is a promising therapy for HIV-1 infection not only for HARRT-naive patients but also for treatment-experienced patients infected with multidrug-resistant, CCR5-tropic HIV-1. 36 –39 In addition, since R5 but not X4 HIV-1 strains are the major transmitters of primary HIV-1 infection, 26,27 the therapeutic targeting of CCR5 as a prophylaxis for HIV-1 prevention appears reasonable and appropriate. Thus, on the basis of the results of the present study it can be hypothesized that OX40 stimulation may have a therapeutic potential for the prevention of R5 HIV-1 infection. The choice of methods to activate CD4+ T cells via the OX40 pathway will obviously be critical for optimal therapeutic benefit with the knowledge that such activation is a two-edged sword and thus strategies that include methods to finely regulate the extent and duration of OX40-mediated activation need to be identified. In this regard, the use of soluble recombinant human OX40L may be one approach since the concentration of the recombinant OX40L required for HIV-1 suppression was around 1 μg/ml (Fig. 2). Another strategy that has been utilized to increase the efficiency and bioactivity of OX40L involves the generation of multimeric forms of OX40L trimer molecules, which have been shown to be markedly more potent in IL-8 production using an OX40-exressing cell line. 23 Other strategies that may be worthwhile to investigate include the generation of viral vectors that express OX40L or OX40L mRNA-transfected dendritic cells. 40,41 We submit that the use of T cells expressing OX40L that are induced by ex vivo activation under DNA-damaging conditions for autologous transfusion-based therapy may provide yet another therapeutic option. 32 Further studies exploring the direct in vivo effects of OX40 stimulation on HIV-1 production in the SIV-infected nonhuman primate model and in the hu-PBL-SCID mouse model are in progress.
Similar to our findings on OX40 costimulation, it is well known that a strong CD28 costimulation together with CD3 stimulation of purified CD4+ T cells from HIV-1-negative or HIV-positive humans results in inhibition of R5 HIV-1 infection due to the production of the three CCR5-binding β-chemokines. 42,43 Data from our preliminary experiments show that activated PBMCs stimulated with immobilized anti-CD3 mAb without additional CD28 costimulation were permissive to R5 HIV-1 proliferation in vitro irrespective of OX40L stimulation (data not shown), suggesting that the CD28 costimulation is essential for OX40-mediated R5 HIV-1 suppression. As it has been demonstrated that CD28 costimulation augments and sustains OX40 expression on T cells in the murine system, 24,44 we also confirmed it in our human system (data not shown). It might be possible that CD28 ligation is required to enhance OX40 function. On the other hand, it will be interesting to reveal whether an OX40–OX40L interaction is involved in CD3/CD28 costimulation-mediated R5 HIV-1 inhibition.
Finally, since OX40-stimulation has been shown to include adjuvant effects, 1,2,9 it is reasoned that OX40 activation in addition to its anti-R5 HIV-1 effect would also promote CD4+ and CD8+ T cell immune responses against HIV-1 and enhance OX40+ NKT cell and neutrophil activities. 8,9 Indeed, it has been shown that OX40 ligation of human CD4+ T cells enhanced HIV-1-specific CTL responses in vitro. 45 These OX40 activities in concert may have great therapeutic potential in HIV-1-infected individuals.
Footnotes
Acknowledgments
This work was supported by grants from a Grant-in-Aid from the Ministry of Education, Culture, Sports, Science, and Technology of Japan and Research on HIV/AIDS and Health Sciences focusing on Drug Innovation from the Ministry of Health, Labor and Welfare of Japan.
Author Disclosure Statement
No competing financial interests exist.