
Abstract
Moesin is a member of the ERM (ezrin, radixin, moesin) family of cytoskeleton/membrane structure organizing and signal transduction proteins. Previously, we found an increased expression of moesin during HIV-1 infection. Moesin was also reported to be incorporated into HIV-1 virions. To analyze whether moesin is a host factor affecting the replication cycle of human immunodeficiency virus type 1 (HIV-1), we used small interfering RNAs (siRNAs) to evaluate the effect of moesin knockdown on HIV-1 replication in P4-CCR5 cells. Moesin's knockdown did not affect the cell viability or cell phenotype. Interestingly, we observed a marked increase in viral replication, as demonstrated by enhanced HIV-1 RNA, p24 antigen, and ß-galactosidase reporter expression. Moesin-dependent enhancement of HIV-1 replication was confirmed in lymphocytic host cells (Jurkat). These results suggest an overall rather restrictive role of moesin for HIV-1 replication in host cells in vitro.
HIV-1
We previously identified moesin as being induced during HIV-1 infection by differential display screening. 1 Moesin (membrane-organizing extension spike protein) belongs to the ERM family (ezrin, radixin, moesin) of cytoskeleton organizing and signal transduction proteins. ERM proteins are membrane-cytoskeleton linkers that play an important role in the formation of microvilli, filopodia, membrane ruffles, and cell–cell junctions. These proteins also participate in signal transduction pathways that connect integrin receptor activation via Rho family GTPases with the cytoskeletal regulation. 10,11 It was shown that moesin is specifically incorporated into HIV-1 virions. 12 Proteolytic fragments of moesin derived from HIV-1 protease cleavage sites were detected in HIV-1 virions 12 and moesin was also found to bind to HIV-1 gp120. 13
To examine whether moesin is an important host factor impacting HIV-1 infection, the effect of moesin knockdown on HIV-1 replication was tested using small interfering RNAs (siRNA).
P4-CCR5 14 cells (HeLa CD4+ CCR5+ long terminal repeat-LacZ), H9 cells, and C8166 cells were cultured as previously described. 15 The HIV-1 strain LAI was taken from the supernatant of freshly infected H9 cells and titrated (TCID50 units/ml) on C8166 cells. 16
siRNAs with specific sequences for moesin (HP Validated siRNA, #SI00300888 and SI 02664130; Qiagen, Hilden, Germany), HIV-1 p24 17 as a positive control or nonsilencing siRNA (labeled with rhodamine) as a negative control were employed (Table 1). Transfections of P4-CCR5 cells (5×104 per well) were carried out with Lipofectamine 2000 transfection reagent (Invitrogen) and siRNA at 30 nM according to the manufacturer's recommendations. Flow cytometry was performed by FACScalibur and Cell Quest software (Becton Dickinson, Heidelberg, Germany). Cellular viability was tested by the WST-1 assay (Roche, Penzberg, Germany). P4-CCR5 cells were infected with HIV-1LAI in triplicate at a multiplicity of infection (MOI) of 0.01 24 h after siRNA transfection in the presence of 50 μg/ml DEAE-Dextran (Sigma, Taufkirchen, Germany) for 4 h.
This table lists the sense and antisense primers with 5′ FAM-labeled and 3′ TAMRA-labeled probes for quantitative TaqMan RT-PCR analyses of moesin, HIV-pol, and GAPDH (housekeeping gene) expression. The second part of the table presents the sense and antisense strands of synthetic siRNA oligonucleotides used for knockdown of p24 and of the rhodamine-labeled nonsilencing control siRNA. The proprietory siRNA sequences specific to moesin from Qiagen have not been published (see catalogue reference).
RNA was extracted using the RNeasy mini kit including treatment with RNase-free DNase I (Qiagen). Synthesis of cDNA was carried out using random hexamer primers and Superscript-II RNaseH-reverse transcriptase according to the manufacturer's specifications (Invitrogen).
Real-time polymerase chain reaction (PCR) was performed in duplicate reactions employing ABI PRISM 7700 (Applied Biosystems, Darmstadt, Germany) with standard conditions (50°C for 2 min, 95°C for 10 min, and 40 cycles at 95°C for 15 s and 60°C for 1 min) as previously described 18 with specific primer and probe sets (Table 1). HIV-1 p24 ELISA was performed using a commercially available kit (Beckmann Coulter, Krefeld, Germany). The β-galactosidase assay and β-Gal staining of cells have been described before. 15
We previously showed that moesin transcription increased early during HIV-1 infection of CEM T cells. 1 To analyze whether moesin has an impact on HIV-1 infection, we evaluated the effect of moesin knockdown on HIV-1 replication using P4-CCR5 cells transfected with synthetic siRNA duplexes specific to moesin, which were subsequently infected with HIV-1LAI. P4-CCR5 HeLa cells were employed because they permitted a higher transfection efficiency compared to lymphocytic cell lines. Quantitative real-time PCR revealed an approximately 3-fold up-regulation of moesin expression associated with HIV-1 infection versus mock-infected cells at 2 days postinfection (Fig. 1B). HIV-1 viral mRNA transcript levels normalized by cell number (housekeeping gene GAPDH) 19 showed that HIV-1 replication was still in the early logarithmic phase at day 2 (Fig. 1A), suggesting that moesin expression was induced by HIV-1 only early on day 2.

HIV-1 infection kinetics and moesin expression.
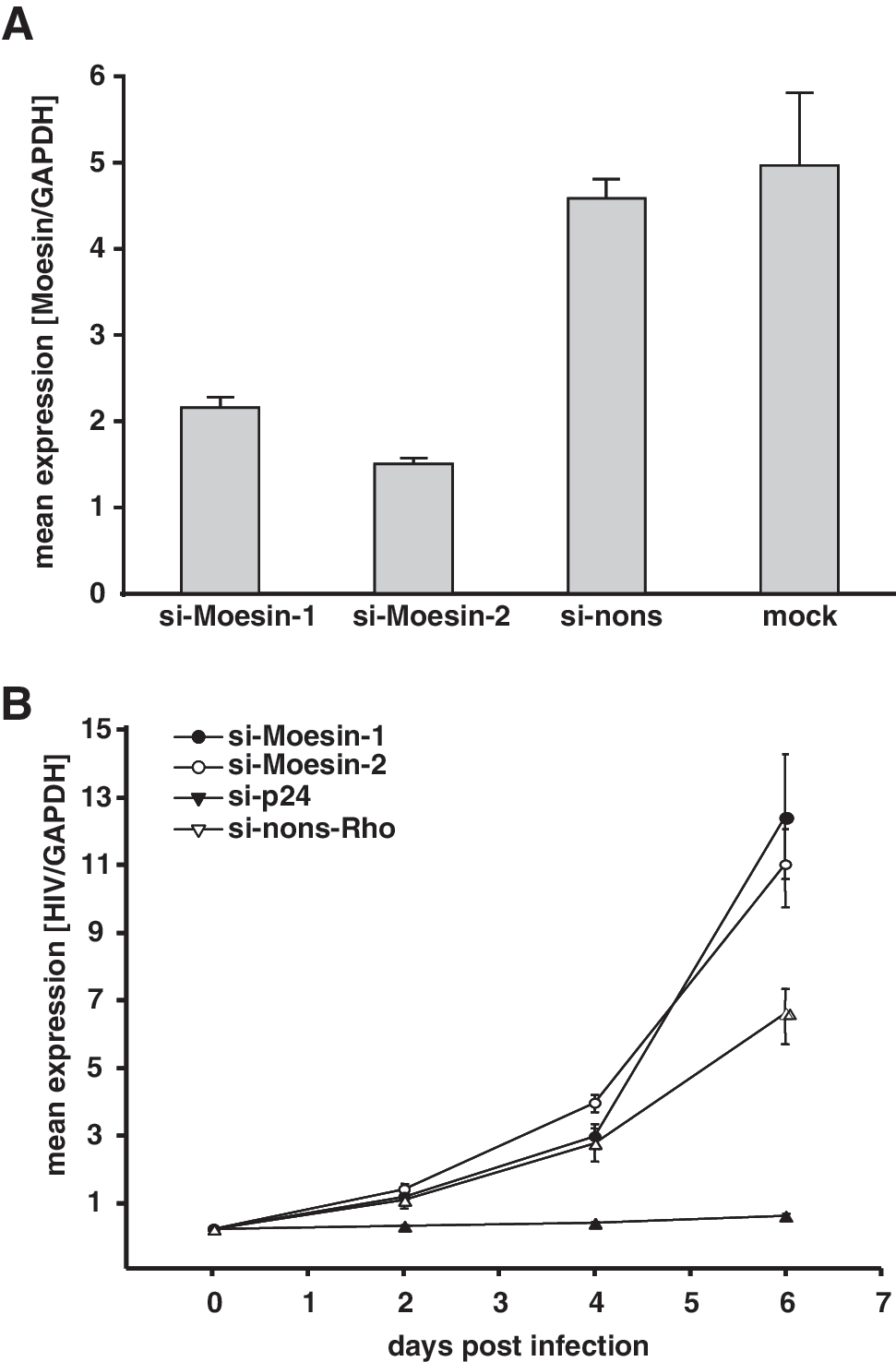
P4-CCR5 cells were transfected with siRNAs directed against moesin and a nonsilencing siRNA, which was 5-prime labeled with rhodamine. Flow cytometry analysis of nonsilencing siRNA-transfected cells at 24 h posttransfection revealed transfection efficiencies of about 90%. The relative levels of moesin mRNA transcripts were significantly decreased by more than 90% after 2 or 4 days, as shown by quantitative PCR (Fig. 2A). The viability and proliferation rate of P4-CCR5 cells transfected with siRNAs against moesin or with the nonsilencing siRNA remained unchanged as determined by the WST-1 cell proliferation assay (Fig. 2B). Thus, moesin-directed siRNAs efficiently reduced the expression of moesin without a general effect on cell viability.

Gene silencing efficiency for moesin and cell viability. P4-CCR5 cells were transfected with two different siRNAs (final concentration of 30 nM) targeting moesin (si-Moesin-1 or si-Moesin-2) or with nonsilencing control siRNA (si-nons-Rho) or no siRNA (mock).
To examine the effect of moesin knockdown on HIV-1 replication, P4-CCR5 cells were transfected with moesin-specific siRNAs and subsequently (24 h later) infected with HIV-1LAI. The nonsilencing siRNA (si-nons-Rho) was utilized as a negative control while the siRNA targeting HIV-1 p24 was used as a potent positive control, as previously demonstrated. 17 Sequential cell-associated HIV-1 viral mRNA levels were determined by real-time quantitative PCR during 6 days after infection. As expected, the positive control siRNA (si-p24) exhibited a marked inhibition in viral mRNA transcription. In contrast, the siRNA-mediated reduction of moesin expression was followed by significantly increased viral mRNA transcript levels at day 4 and day 6 compared to HIV-1-infected controls, which had been transfected with the nonsilencing siRNA (si-nons-Rho) (Fig. 3A).

Moesin knockdown and HIV-1 replication. P4-CCR5 cells were transfected with siRNAs against moesin (si-Moesin-1 or si-Moesin-2), HIV-1 p24 (si-p24), nonsilencing siRNA (si-nons-Rho), or without siRNA (mock). Cells were infected with HIV-1LAI (MOI 0.01) 24 h posttransfection. Cells and supernatant samples were collected at 0, 2, 4, and 6 days after infection for quantitative PCR analysis of HIV-1 transcript levels
The effect of moesin suppression on HIV-1 replication was confirmed by p24 ELISA, showing a significantly increased p24 expression at day 4 postinfection compared to controls with nonsilencing siRNA (Fig. 3B).
Furthermore, the effect on HIV-1 replication was examined by the integrated HIV-1-LTR-driven reporter vector expressing ß-galactosidase in P4-CCR5 cells. HIV-1 Tat-mediated transactivation of the LTR leads to expression of measurable ß-galactosidase activity, allowing measurements of enhancing effects on HIV-1 replication as increases in ß-galactosidase activity. ß-Galactosidase activity on day 4 after infection was markedly increased by siRNA suppressing moesin compared to the nonsilencing control siRNA (Fig. 4A). The induction of LTR-driven transcription was confirmed by microscopy of these cell cultures after X-Gal staining on day 6 postinfection (Fig. 4B).
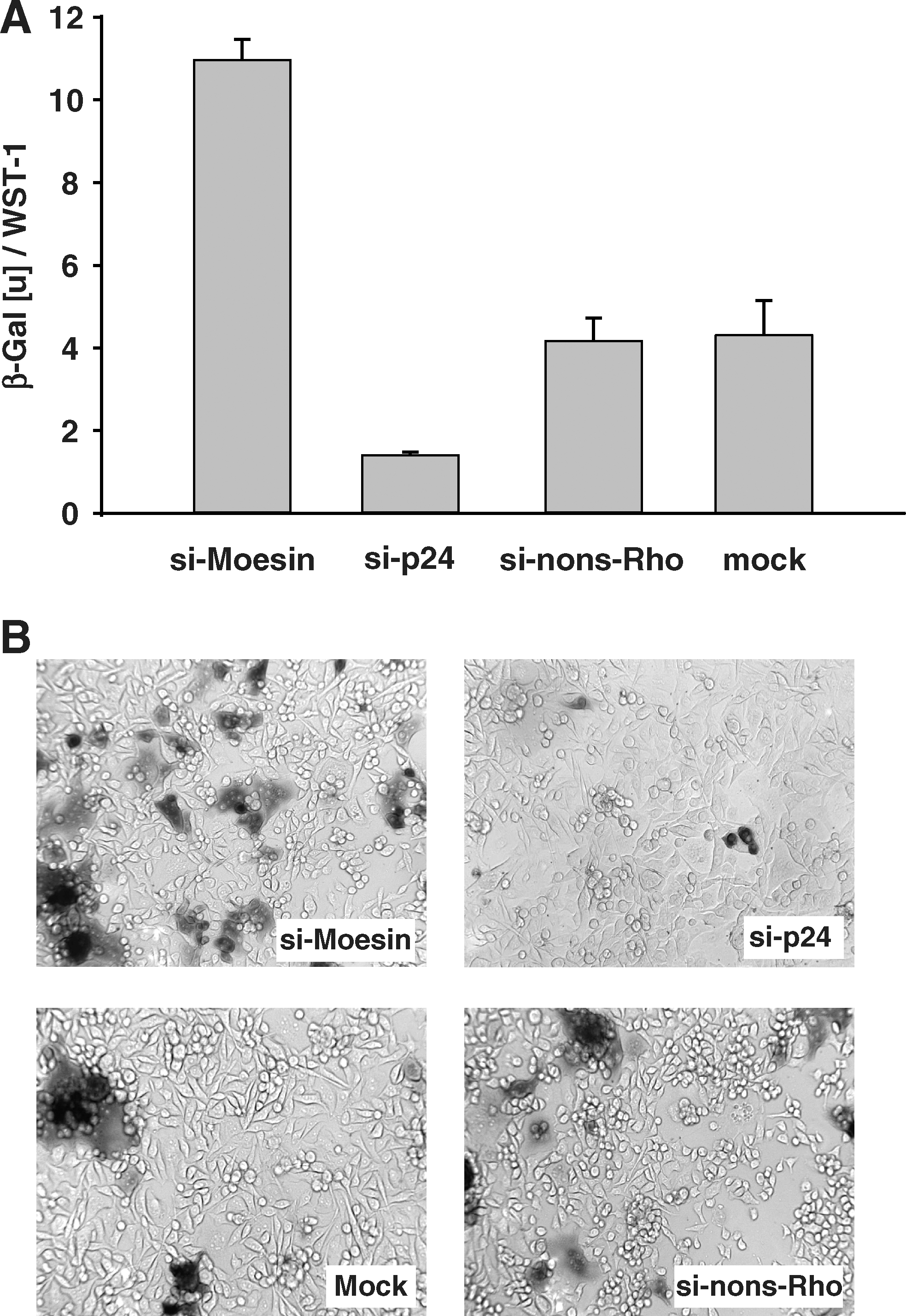
Silencing of moesin and LTR transactivation in HIV-1-infected cells. P4-CCR5 cells contain one integrated copy of the HIV-1 LTR linked to the ß-galactosidase (ß-Gal) reporter gene. HIV-1 Tat-mediated transactivation of the LTR leads to expression of quantifiable ß-Gal.
These results demonstrate that siRNA-mediated suppression of moesin increased HIV-1 replication and indicate a possible function of moesin in restricting HIV-1 replication.
To examine whether the results observed in HeLa cells (P4-CCR5) can be confirmed in lymphocytic cells, the typical host cell of HIV-1, we tested the effect of knockdown of moesin expression on HIV-1 replication in Jurkat cells. Jurkat cells were transfected with two different moesin-specific siRNAs, respectively, which suppressed the ratio of moesin/GAPDH mRNA copy numbers to 47% (si-Moesin-1) and 33% (si-Moesin-2) compared to nonsilencing control siRNA (si-nons-Rho) (Fig. 5A). HIV-1 replication normalized by GAPDH for cell numbers was quantified by real-time PCR in serial samples in infected Jurkat cells, which had been transfected with siRNAs specific to moesin or an unspecific control before. The Jurkat cultures transfected with si-RNAs against moesin showed a significant increase in HIV-1 replication compared to the nonsilencing siRNA control on day 6 postinfection (Fig. 5B), while the positive control si-p24 specific to HIV-1 directly inhibited HIV-1 replication. The weaker suppression of moesin expression in Jurkat cells compared to P4-CCR4 cells may be attributed to the lower transfection efficiency in lymphocytic cell lines observed in general. Thus, it may be explained that the Jurkat system exhibits slower kinetics (only marked on day 6) for the impact of moesin knockdown on HIV-1 replication than the HeLa system.
Moesin knockdown and HIV-1 replication in Jurkat cells. Jurkat cells were transfected with siRNAs against moesin (si-Moesin-1 or si-Moesin-2), HIV-1 p24 (si-p24), or nonsilencing siRNA (si-nons-Rho) and analyzed for moesin expression by qRT-PCR
The present study examined moesin's role in HIV-1 replication, since moesin had been found to be up-regulated during HIV-1 infection of CEM T cells. 1 Based on its diverse roles in membrane structures 20 and membrane trafficking 21,22 and its incorporation into HIV-1 particles, 12 it was hypothesized that moesin may exert a function in viral entry, assembly, or budding.
Moesin-directed siRNAs efficiently reduced the expression of moesin without an effect on growth rates of HeLa and P4-CCR5 cells, indicating that moesin was not required for cell viability. Furthermore, no phenotypic changes in cell shape, size, or cell adhesion were observed. This finding is in agreement with the observation that moesin suppression did not cause phenotypic changes in mouse cell lines. Only when moesin was knocked down in conjunction with ezrin and radixin, a marked reduction of membrane protrusions was observed. 23 Furthermore, moesin-null mice were viable and was displayed no obvious distinct phenotype. 24 Therefore, it has been proposed that other ERM family proteins may be able to substitute for moesin.
While our study was ongoing, several interesting papers have been published regarding moesin's involvement in the HIV-1 entry mechanism. Naghavi et al. found that overexpression of moesin blocked infection of rat cell lines by HIV-1 pseudotyped virus and knockdown of moesin enhanced infection. 25 They showed that moesin regulates stable microtubule formation and limits retroviral infection, suggesting a role in early postentry events. Kubo et al. found that HIV-1 vectors pseudotyped with VSV-G and R5-tropic HIV-1-Env (strain JRFL or NH2) exhibited an increased transduction of TE671/CD4/R5 cells upon moesin knockdown. 26 Our results are basically in line with these findings. However, deviating from their results with X4-tropic (NDK or HXB2) pseudotyped HIV-1 that comprised a decreased transduction rate following moesin knockdown, we observed an increased replication rate in P4-CCR5 HeLa cells with the genetically unmodified x4-tropic strain HIV-1LAI. While the data of the other groups are based on different cellular systems with nonreplicative pseudotyped virus recombinants focusing on early events from viral entry until reverse transcription, our data reflect a system of several replication rounds with an unmodified HIV-1 strain.
Barrero-Villar et al. reported an inhibition of HIV-1NL4.3 infection in moesin-silenced CEM T cells. 27 The difference from our data may be attributed to the usage of distinct host cells, isolates, and MOIs. Furthermore, our data pertain to a longer observation period of 2–6 days postinfection, whereas the other group evaluated HIV-1 infection within 3 days. A review by Liu et al. summarizes the complicated interaction between CD4/CXCR4 receptors, ERM proteins, filamin, cofilin, and actin. 28 Apparently actin polymerization mediated by moesin promotes initial virus binding and fusion, but inhibits subsequent early postentry events, thus possibly reconciling the different observations.
Our results indicate a net restrictive effect of moesin over several rounds of replication. Therefore, the observed transcriptional up-regulation of moesin during HIV-1 infection may lead to the hypothesis that the forced overexpression of moesin may be associated with a reduction of HIV-1 replication, which may be examined in future studies.
An immunological role of moesin has been supported by other observations. Moesin expression was shown to be increased after lipopolysaccharide (LPS) stimulation. 29 LPS-induced tumor necrosis factor (TNF)-α secretion could be blocked by an antibody 30 or antisense oligonucleotide specific to moesin. 29 Similar to bacterial infections, moesin may be involved in signaling for cytokine expression in viral infection. It has been well established that interferon-alpha and interferon-beta (IFN-α/β) are induced in virus-infected cells. 31 IFN-α/β trigger a program for antiviral response. Interferon-stimulated gene 15 (ISG15) was found to be induced as a response to viral infection 32 –35 and to IFN-α/β treatment. 36,37 Moesin has been shown to be a target of ISG15 and modified upon IFN-ß treatment of HeLa cells. 38
It remains an open question as to whether moesin is involved in the process of viral assembly, budding, or maturation in addition to regulating viral entry. Furthermore, moesin's role in T cell migration, T cell receptor signaling, cytokine production, and CD95-induced cell death 39 may imply pleiotropic effects in complex biological systems such as animal models of HIV-1 or HIV-1-infected patients.
Footnotes
Acknowledgments
We are grateful to the H. W. & J. Hector Stiftung (foundation), Mannheim, Germany, which provided a grant to support this research.
Author Disclosure Statement
No competing financial interests exist.