
Abstract
There was an outbreak of HIV-1 transmission among 20 out of 122 Korean hemophiliacs from 1990. We assessed the genetic relationships among HIV-1 viruses found in three cash-paid plasma donors whose preseroconversion plasma was used to produce Korean-made clotting factor, 20 hemophiliacs infected with HIV-1 in Korea, three hemophiliacs infected with HIV-1 from clotting factor manufactured outside Korea, and 71 local control patients infected with the Korean subclade of HIV-1 subtype B (KSB). Full-length pol gene sequences (2841 bp) of viruses from frozen stored serum, samples obtained 1–3 years after diagnosis, were amplified by RT-PCR and sequenced by direct DNA sequencing. Phylogenetic and signature pattern analyses were used to investigate the relationships among the sequences. Donors O and P were associated with two clusters, of 8 and 12 hemophiliacs, respectively, which were demarcated from the 71 KSB-infected local control patients and donor R. These data confirm that HIV-1 transmission to 20 hemophiliacs occurred through infusion of Korean-made clotting factor.
Introduction
P
Two clusters of HIV-1 spread among hemophiliacs (HP) have been reported after 1988, one in Germany in 1989 6 and the other in Korea in late 1989 to early 1990. 14 Interestingly, both clusters consisted primarily of patients with hemophilia B. 6,14 We have previously investigated the Korean event through phylogenetic analysis of HIV-1 nef and pol gene sequences in peripheral blood mononuclear cells (PBMCs). 14 Importantly, all of these Korean HPs were infected with the Korean subclade of HIV-1 subtype B (KSB), 14 –18 a variant quite distinct from the worldwide consensus of HIV-1 subtype B. 17 This excluded the possibility that these patients were infected due to clotting factors imported into Korea before the production of Korean-made domestic clotting factor (DCF) began in late 1989. In addition, we observed genetic associations among the viruses of 20 HPs and those of HIV-1-infected plasma donors whose preseroconversion plasma was used to make DCF. Furthermore, the official report by the second investigation committee (established in 2002) on this accident revealed a statistically significant association with six lot numbers of DCF manufactured in 1990–1991 (Molecular and Epidemiological Report on HIV-1 Outbreak among HPs, issued in April 2004, pp. 42–43).
We have expanded our earlier analyses for several reasons. First, there was a long interval between HIV-1 diagnosis (November 1989, based on seroconversion) and sampling times from plasma donor O (February 1999) and the 20 HPs. Second, we had previously sequenced only 543 bp of the pol gene in HIV-1 from donor P (AY347694), and less than 957 bp in the other patients. Thus, the pol sequences from donor P had not been analyzed in our earlier phylogenetic analysis. 14
In this study, we used RT-PCR to amplify the near full-length pol gene (2841 bp) from sera obtained 1–3 years after diagnosis in most HPs, plasma donors O, P, and R, and 71 local controls infected with KSB (LCs). Moreover, we used a stored frozen serum sample from donor O that was obtained much earlier (January 1992) than the February 1999 PBMCs used in the previous report. We also assessed pol sequences in 36 new patients whose PBMCs had not been available earlier. In addition, signature pattern analyses in full-length pol gene were used to investigate the relationships among the sequences. There were significant differences at 19 nucleotide positions compared with 71 local controls. Among them, a specific nucleotide in the two and one positions was detected in clusters O and P only, respectively. Taken together, we found that plasma donors O and P were associated with two clusters, of 8 and 12 HPs, respectively, demonstrating that these 20 HPs had been infected with HIV-1 through DCF.
Materials and Methods
Plasma donors O and P: sources of HIV-1
In the late 1980s, plasma was collected for cash at domestic plasma centers run by Company X, which manufactures various blood products, including clotting factors. Four HIV-1-seropositive homosexual plasma donors, O, P, Q, and R, were detected during primary infection (Table 1). 14 Although no postseroconversion plasma from these donors was used in the production of DCF, preseroconversion plasma from these donors had already been used to produce DCF. These latter samples were not heat treated, although they were treated with a surfactant/detergent thought to be sufficient to disrupt enveloped viruses. A previous report on the nef gene found that viruses from donors Q and R, both of whom were diagnosed in 1992, were not associated with HIV-1 infection in HPs. 14
Distribution of HIV-1-infected hemophiliacs showed a significant difference in geographic distribution. All patients were from Seoul, Kyunggi-do, Daegu, Kyungnam, Ulsan, and Busan Metropolitan Cities axis with a local branch of manufacturer for the high population density (22%; 20/91). There was no such occurrence in Jeollanam/buk-do, Chungnam/buk-do, Gangwon-do, and so on without a local branch (0%; 0/39) (p < 0.001).
Patients were being treated with antiretroviral drugs and revealed resistance mutations (Table 5).
Chungnam; regarding hemophilia, patient HP-18 did not visit any hospital in Chungnam province and had been admitted to a hospital and had visited a hemophilia clinic in Seoul. The relative risk for HIV-1 infection of hemophiliacs in Daegu/Gyeongbuk and Seoul provinces based on the local distribution of hemophiliac patients in 1992 were about 4.9 and 3.3 compared with other provinces, respectively.
This study for the first time reports on pol gene in donors-P and R.
We did not obtain the pol gene from donor-Q.
Twenty HIV-1-infected HPs and local controls
HPs 1–20 were diagnosed with HIV-1 infection between 1990 and 1994 (Table 1). 14 They had been treated primarily with imported clotting factors prior to the start of local DCF production. Three HPs (nos. 21–23) were infected with clotting factor purchased outside Korea before 1987 (Table 1). Except for a dead patient diagnosed in 1988 (case 4), 19 we included all homosexual patients diagnosed before 1990 (n = 15) and 17 of 18 homosexual patients diagnosed in 1990. Including these 32 patients, we determined sequences of the pol gene in 74 local control patients infected with KSB including three plasma donors. In addition, we determined the HIV-1 gene in 15 local patients infected with non-KSB or non-subtype B. Informed written consent was obtained from all living study participants.
RNA preparation and pol gene amplification
The earliest sera samples used in this study, from 15 HPs, were collected between April 1990 and April 1993 from patients newly diagnosed with HIV-1 infection at the division of AIDS of the Korean NIH. These blood samples had been collected at 6-month intervals for CD4 T cell measurements. Samples from the other five HPs were obtained at later dates (Table 1). RNA was extracted from 300-μl serum samples using a QIAamp Ultra sense Viral RNA kit (Qiagen, Hilden, Germany) and 2-μl aliquots of RNA were reverse transcribed by mixing with 1 μl oligo(dT), 1 μl dNTPs, and 6 μl DEPC-treated water, and incubating the mixture at 65°C for 5 min and then on ice for 1 min. To each sample was added 4 μl 5 × buffer, 2 μl 0.1 M DTT, 2 μl DEPC-treated water (Ambion Inc., Foster City, CA), 1 μl RNase-Out, and 1 μl Superscript3 enzyme; the samples were incubated for 60 min at 65°C, followed by inactivation of the enzyme for 15 min at 95°C (Invitrogen, Carlsbad, CA). The pol gene was amplified by nested PCR with TaKaRa R-Taq (TaKaRa Bio, Inc., Otsu, Shiga, Japan), with full sequences obtained by the overlap of three PCR products. Details are described in earlier reports. 18,20 –22 The subsequent amplicons were directly sequenced using Applied Biosystems 3730XL (Macrogen Inc., Seoul, Korea).
Sequences
The GenBank accession numbers for the pol sequences are HM210883–HM210884 and HQ026610-11 from donor O, HM210885–HM210886 from donor P; HM210887–HM210888 from donor R, HQ026569–HQ026609 and HQ026620–HQ026623 from 23 hemophiliacs, and HQ026498–HQ026568, HQ026612–HQ026619, and HQ026624–HQ026635 from local controls. The GenBank accession numbers for the vif sequences are DQ054367, DQ072731–DQ072850, DQ295192–96, DQ837381, AF224507, AF286239, AY581320–AY581423, and AY839827.
Phylogenetic analysis
Sequences were aligned to the HIV-1 subtype reference set from the HIV Sequence Database (
Viral signature pattern analysis
The VESPA (
Statistical analysis
Data are expressed as means ± standard deviations. Statistical significance was estimated using the Student's two-tailed t-test, the Chi-square test, and Fisher's exact test, using SPSS package version 12.0. Statistical significance was defined as p < 0.05.
Results
Origin of the Korean subclade of subtype B
A total of 327 HIV-1 infected patients (1 in 1985, 4 in 1986, 9 in 1987, 22 in 1988, 37 in 1989, 54 in 1990, and 200 in 1991–1993) were diagnosed in Korea up to 1993 (Table 2). To date, all overseas sailors whose HIV-1 sequences was determined (n = 19) revealed non-KSB of HIV-1 (non-B in 18 patients) and their wives (n = 7) also revealed subtypes corresponding of their husbands. Nine of the 14 patients diagnosed in 1985–1987 were commercial sex worker who had sexual contact with U.S. soldiers at U.S. military camps in Korea, two were hemophiliac patients, two were infected in Africa, and one was a man who had sex with a man (homosexual). We determined sequences in seven commercial sex worker. Six revealed western subtype B (non-KSB) and one revealed CRF02-AG, indicating that KSB did not begin to circulate in 1987. Thus, all patients who lived outside Korea, the overseas sailor/his wife, and prostitutes revealed non-KSB (Table 2).
Two hemophiliacs infected with HIV-1 in the United States and two males living in Africa.
Based on epidemiological data, 172 patients, including 20 HPs, were regarded as being infected locally in Korea (Table 2). We sequenced nef and/or vif genes earlier than the pol gene to determine whether the subtype in patients without known HIV-1 sequences is KSB. In case of KSB, we next amplified the pol gene. Almost all of these 172 patients were found to share an apparent recent common ancestral virus, although a few patients with insufficient epidemiological data showed discrepancies between their HIV-1 subtypes and epidemiological data. The local circulating virus has been classified as KSB. We amplified and sequenced full-length pol gene sequences in 109 of these patients (Table 3).
Among the 22 and 42 patients, 18 and 38 were diagnosed after 1993, respectively.
In addition to 74 full-length pol genes, we determined partial pol genes in nine patients (number in parenthesis).
Considering the seroconversion time of donors O and P (late 1989–early 1990) and the first diagnosis of HIV-1 infection in HP-1 in 1990, we hypothesized that viruses from patients diagnosed before 1991 were more likely to be sources of transmission of HIV-1 to the 20 HPs than those from patients diagnosed after 1990. We therefore attempted to include as many patients as possible who had been diagnosed before 1991. Because all four plasma donors with seroconversion identified at the plasma center were male homosexuals, we amplified the vif, or nef and/or pol genes in 31 of all 32 males infected with HIV-1 through homosexual contact before 1991; one patient, diagnosed in 1988, who died prior to obtaining samples remains unknown (Tables 1 and 2).
We amplified the full-length pol gene in 20 of 32 KSB-infected patients who had been infected with HIV-1 through homosexual contact and diagnosed up to December 1990 (Table 3). In addition, the pol gene was partially sequenced, for one-third or two-thirds of its full length in five patients (including one indirectly by amplification of a blood recipient) up to 1990. This study, including 78% (25/32) of homosexually infected patients diagnosed before 1991, is therefore a comprehensive, nationwide investigation. Thus, we found that KSB had already begun to be transmitted in 1988.
Molecular epidemiologic data from the pol gene
We determined the full-length pol gene in 109 patients: 71 LCs, 3 plasma donors, and 20 HPs (nos. 1–20) infected with KSB, 3 HPs (nos. 21–23), and 10 non-KSB (1 subtypes D, 1 CRF02_AG, and 8 subtype B). For phylogenetic analysis, we analyzed 2841 bp, from nucleotides 2253 to 5093, with 92 KSBs (Fig. 1). These 92 KSBs included two clusters, consisting of 20 HPs and donors O and P; donors O and P were associated with clusters O and P, which included 8 and 12 HPs, respectively (Table 3). In contrast, donor R, who was diagnosed in 1992 (after the first investigation committee in 1992), did not cluster with any HP (Fig. 1).
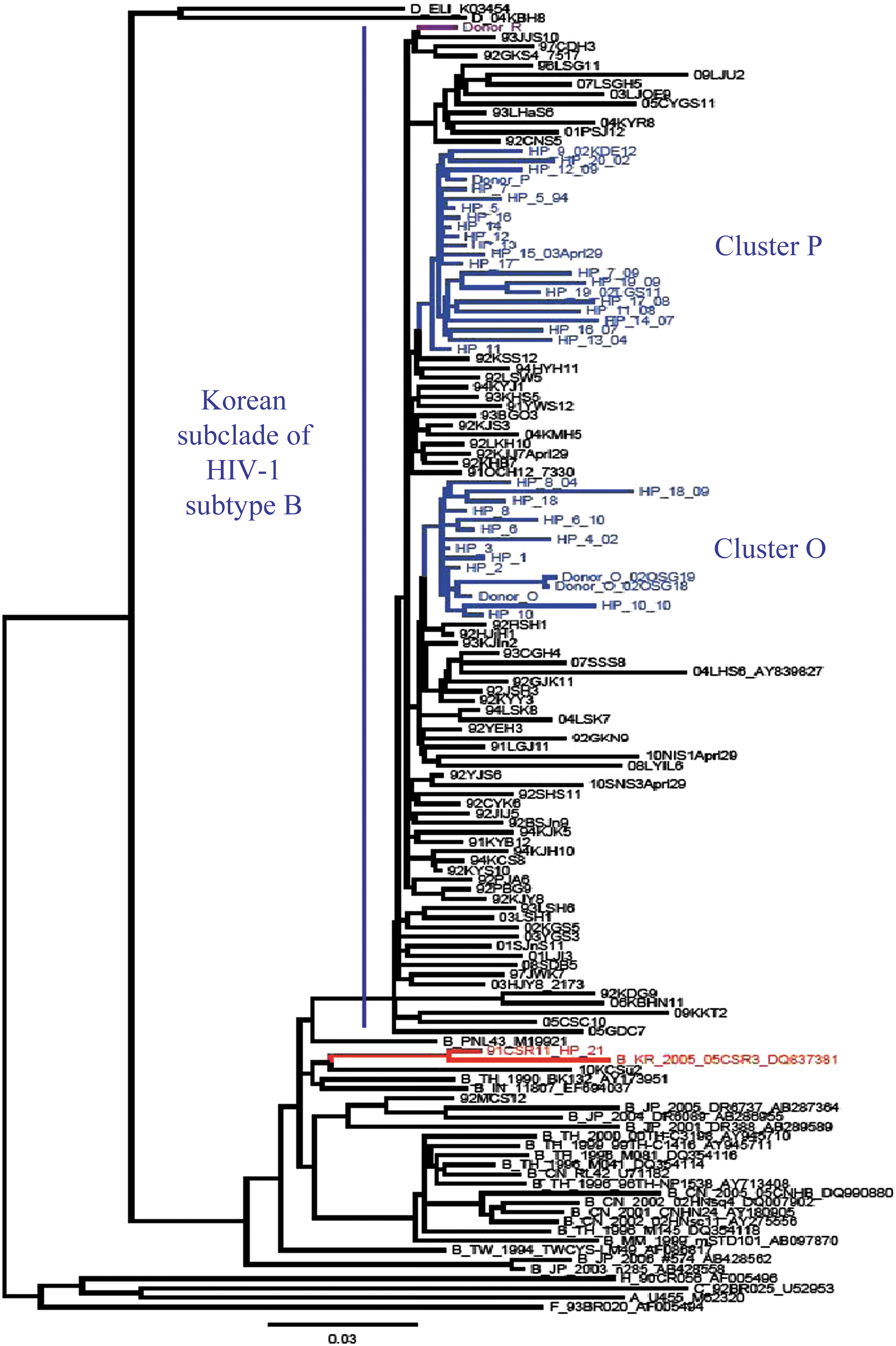
Phylogenetic tree of full-length pol gene sequences (2841 bp) of 20 hemophiliacs (HPs), plasma donors O, P, R, and 68 local control patients infected with Korean subclade of HIV-1 subtype B (KSB) and non-KSB including HP-21. Sequences were aligned with the HIV-1 subtype reference set from the HIV Sequence Database (
We also determined full-length pol sequences from samples taken at different time points from 12 HPs. We found that each sample belonged to the same cluster as the earliest sample from that individual, without cross-exchange between the two clusters (Fig. 1).
Nucleotide/amino acid signature patterns
When we assessed the signature pattern nucleotides in clusters O and P, we found that 11 positions in cluster O and 8 in cluster P showed statistically significant relationships with nucleotide sequences in the 71 LCs and donor R (p < 0.01), along with a significant difference between the two clusters (p < 0.01) (Table 4). Changes in nucleotides at 9 positions were nonsynonymous, whereas those at 10 positions were synonymous, compared with the consensus nucleotide sequences of the 71 LCs and donor R. Interestingly, none of the 71 LCs and donor R had G, G, and A at positions 2321, 4235, and 5070, respectively (Table 4). Nucleotide A at position 5070 was found only in cluster O, resulting in a nonsynonymous change at codon 281 in integrase (IN) (V281M) (Fig. 2). V281M was not observed in the remaining 146 patients, including the 71 LCs.

Alignment of pol gene nucleotides in three regions showing 100% in clusters O or P versus 0% in 72 local controls. The first three nucleotides (2319–CTA-2321), the second three nucleotides (4233–TTA-4235), and the remaining third regions (5041–5093) correspond to the protease, integrase, and integrase region, respectively. The third region also corresponds to the initial 53 nucleotides of the vif gene (based on NL4-3). Sequences were obtained from 20 hemophiliacs (HP), three plasma donors (O, P, and R), and 71 local control patients infected with the Korean subclade of HIV-1 subtype B (KSB), three HPs infected with non-KSB and reference strain NL43, and four patients infected with four different non-subtype B. All 11 sequences and three sequences obtained from donors O and P revealed nucleotide G at positions 2321 and 4235, respectively. Nucleotide G at positions 2321 and 4235 were found in clusters O and P only, respectively, resulting in a synonymous change. Nucleotide A at position 5070 was found in cluster O only, resulting in a nonsynonymous change at codon 281 in integrase (V281M). Among these three positions, only position 5070 was extremely conserved irrespective of subtype. In addition to the 92 patients shown, the remaining 54 patients also carried nucleotide “G” only. The dots indicate sequence identity compared with consensus.
Nine nucleotide positions with nonsynonymous changes are shown in bold. There were significant differences in eight positions between cluster P and background (71 LCs plus donor R), and in 11 positions between cluster O and background (71 LCs plus donor R) (p < 0.01 for all 19 positions). G and A at positions 2321 and 5070, respectively, were detected only in cluster O, whereas G at position 4235 was detected only in cluster P. Thus, V281M in integrase was detected only in cluster O, but not in the remaining 146 patients (p < 0.0001).
Presence of integrase V281M in all patients comprising cluster O
We also sequenced the vif gene in 155 Korean patients with HIV-1 (Table 3), including of 125, 20, and 10 patients infected with KSB, subtype B and non-B HIV-1, respectively. We determined 1290 vif sequences, consisting of 1170, 88, and 32 of these respective subtypes. We found that codons 280 to 288, encoding the N-terminal part of IN, could not be amplified by our primer set OBP3/4. The vif gene starts at IN codon 271, indicating that this region was borrowed from vif amplicons. A nonsynonymous change at nucleotide 5070 resulted in a methionine at codon 281 of IN. Surprisingly, IN V281M was observed only in the nine patients constituting cluster O.
We obtained 23 vif amplicons from donor O, five in October 1991 (V281M3 and V281L2), two in January 1992 (V281L2), seven in February 1993 (V281L3, V281V3, and V281M1), and nine after February 1999 (V281V9). We found that codon 281 in donor O, which was M4 or L7 in 11 of 14 samples before 1995 (11/14), changed to V9 after 1995 in all nine samples. However, in five of the eight HPs comprising cluster O, V281V appeared later, together with V281M. The frequency of V281M, which was 94.4% (17/18) in 1990–1992, was 88.9% (16/18) in 1993–1995. In other words, it is presumed that V281M dominated in the beginning.
Except for the 180 vif amplicons from cluster O, all 1110 vif amplicons in the remaining 146 patients carried V281 only. This finding also strongly supports the epidemiological linkage for the transmission of HIV-1 from two plasma donors to 20 HPs.
Presence of resistance mutations to PI and RTI; absence of Q148HKR and N155H
Major resistance mutations in the earliest samples were seen in three HPs (nos. 18–20) who had been treated with antiretroviral agents (Table 5). All of these patients were naive to integrase inhibitor (INI). An assay for a resistance mutation to INI (i.e., Q148RHK and N155H) 24 showed that there was no major mutations (Q148R was present in two patients). Single accessory mutations have been reported, including T97A, V151I, G163R, I203M, and S230N 25 ; however, we found G163E (7/74), I203M (1/74), and S230N (4/74). We found that none of these samples had G140S, which is associated with an in vitro 5- to 10-fold decrease in susceptibility to antiretroviral agents. 26
WT, wild type.
Minor mutation.
Sequence identity of donors O and P
We sequenced full-length pol genes from two samples obtained from each of donors O and P. In donor O, the intrapersonal identities between two amplicons obtained in January 1992 (GenBank accession nos. HM210883–84) and two amplicons obtained in 2002 were 99.6%, and 99.7%, respectively. It was 98.6% between 1992 (HM210883) and 2002 (HQ026610). The intrapersonal identity between two amplicons obtained from donor P in February 1993 (HM210885) and October 1993 (HM210886) was 99.6%.
When we changed resistance mutations into wild-type sequences in three HPs (nos. 18–20), we found that the sequence obtained in February 1993 from donor P was 99.18 ± 0.35% identical to that of cluster P (nos. 5, 7, 9, 11–17, 19, and 20), and that the sequence obtained in January 1992 from donor O was 99.07 ± 0.47% identical to that of cluster O (nos. 1–4, 6, 8, 10, and 18). It significantly increased compared with 98.11 ± 0.89% before this study (p < 0.05). Except for four of five sera obtained from five HPs (nos. 4, 9, 15, 19, and 20) after 2000 and the serum from HP-18, all showed high sequence identity >99.0% compared to their respective donor.
Excluding the cluster O sequences obtained after 2000 (HP-4), we found that sequence identity of the seven HPs, when compared to donor O, increased to 99.22 ± 0.23%. When we compared the donor O sequence to those of six local controls with the highest identity, we found a significant association (98.85 ± 0.15%, p < 0.01). Similarly, the sequence identity of donor P compared to HPs increased to 99.35 ± 0.08%.
Determination of full-length pol sequences at two time points in each of 12 HPs showed an intrapersonal identity of 98.47 ± 0.43% over 125 ± 61 months (range, 16–203 months).
Discussion
We amplified the pol, nef, and vif genes from sera obtained earlier than the PBMC samples used in our previous study, enabling us to extend the follow-up period for genetic variations of KSB in each patient. We sequenced the nef, vif, and pol genes from samples obtained over 18 years in many patients and traced their variations. In addition, our evidence indicates that the origin of KSB 15 in patients diagnosed with HIV-1 could be extended back to 1988.
These detailed genetic sequencing analyses further substantiate the previously described genetic similarities between viruses obtained from HPs in Korea and viruses obtained from the two plasma donors. The most striking feature of our pol gene sequence data was the highly homologous nature of the sequences in plasma donors O and P in 1992–1993 and those in the HIV-1-infected HPs in Korea in 1991–1993. The very few “signature sites,” where sequences from donors O and P share bases with the hemophiliacs but not with the sequences from the other “Korean subclade” of subtype B, are not highly significant (Table 4). The major point is that the hemophiliacs were infected with the Korean subclade of subtype B, which proves that they were not infected with foreign-made clotting factor.
Similar results were also observed by analyzing nef (126 patients infected with KSB including 20 HPs) and vif (in 106 patients infected with KSB) (data not shown). This new evidence adds substantially to the already overwhelming evidence that links the domestically made clotting factor to HIV infections among HPs in Korea.
Transmission clusters are distinguished by high sequence identity, based on the established criteria of high bootstrap values and short branch length. 10 Although our previous study, based on a phylogenetic tree of pol sequences, revealed two clusters of HPs, HP-14 (AF273196 obtained in September 1999) and HP-11 (AY219020 obtained in October 2002), did not belong to either. In contrast, we have shown here that 8 and 12 HPs belonged to clusters O and P, respectively (Fig. 1). These differences may have been due to the use of samples obtained earlier, which showed fewer changes after 90 months (in HP-14) and 123 months (in HP-11) and the sequencing of 3-fold longer sequences.
Remarkably, all nine patients of cluster O carried the integrase mutation V281M (Fig. 2), whereas all other patients carried valine only. The V281M was not described in HIV-1 subtype B, although six sequences were described in non-subtype B (
In Korea, 250 HIV-1-infected patients were diagnosed in 1985–1992. Among the 250 patients, presumably 134 and 116 were infected with non-KSB and KSB, respectively. The combination of our pol, nef, and vif gene data obtained by RT-PCR to our previous data pool revealed the sequence nature of at least one of the three genes in 85 (73%) of the presumed 116 KSB infected patients.
In the present study, sera sampled from different people at different time points relative to one another before and after therapy were used. Despite the different time point in sampling, the signature pattern over 2841 bp was well conserved. Thus, epidemiologically linked sequences (clusters O and P) clustered among 71 KSB in the phylogenetic tree analysis.
In conclusion, the comprehensive and specific findings shown here demonstrate a clear link between DCF and the HIV-1 infections among HPs in Korea.
Footnotes
Acknowledgments
This work was supported by a grant from the Korean Society of Ginseng funded by the Korea Ginseng Corporation (2009–2010). We thank former Dean, professor Jae-Dam Lee, for his encouragement and support.
Disclosure Statement
No competing financial interests exist.