
Abstract
Subtype G has been estimated to represent the fourth most prevalent clade in the HIV-1 pandemic and subtype F is widely circulating in parts of South America (frequently within BF recombinant forms) and in Romania. However, functional envelope clones of these subtypes are lacking, which are needed for studies on antibody-mediated neutralization, coreceptor usage, and efficiency of viral entry inhibitor drugs. Here we report the construction, neutralization properties, and coreceptor usage of HIV-1 functional envelope clones of subtypes G (n = 15) and F (n = 7). These clones were obtained through RT-PCR amplification of HIV-1 gp160 from plasma RNA, and were used for pseudovirus production. All 15 subtype G-enveloped pseudoviruses were resistant to neutralization by gp120-targeted broadly neutralizing monoclonal antibodies (MAbs) b12 and 2G12, while a majority were neutralized by gp41-targeted MAbs 2F5 and 4E10. With regard to the subtype F envelopes, all seven pseudoviruses were resistant to 2F5 and b12, six were resistant to G12, and six were neutralized by 4E10. Coreceptor usage testing revealed that 21 of 22 envelopes were CCR5-tropic, including all 15 subtype G envelopes, seven of which were from patients with CD4+ T cell counts <200/ml. These results confirm the broadly neutralizing activity of 4E10 on envelope clones across all tested group M clades, including subtypes G and F, reveal the resistance of most subtype F-enveloped pseudoviruses to broadly neutralizing MAbs b12, 2G12, and 2F5, and suggest that, similarly to subtype C, CXCR4 tropism is uncommon in subtype G, even at advanced stages of infection.
Introduction
O
It has been recommended that panels of well-characterized reference strains of HIV-1 be developed for each major genetic subtype to aid in the evaluation of the breadth and magnitude of neutralizing antibody responses elicited by candidate vaccines. 17 Previously, panels of functional envelope clones of subtypes A, 14,20 B, 19,21 and C, 12,22,23 and CRF01_AE 24,25 have been developed. However, for subtypes G and F, only one and two functional envelope clones, respectively, have been reported in the literature, derived from in vitro cultured viruses (with low or borderline functionality in the case of the subtype F clones). 26 Subtype G has been estimated to represent the fourth most prevalent clade in the HIV-1 pandemic, corresponding to approximately 6% of global infections. 5 It circulates widely (38–54% prevalence) in Nigeria, 27,28 the most populous African country and the second in the world in number of HIV-1 infections (approximately 2.6 million), 29 and also in Portugal (24%) 30 and Republic of Congo (21%). 31 At lower prevalences, subtype G is also found in other Central and West African countries, Spain, 32,33 and Cuba (here frequently as BG recombinant forms). 34,35 Subtype F circulates in Romania (here as the predominant HIV-1 clade), 36 Central Africa, 37 and Brazil, 38,39 and, as part of BF recombinant forms, is highly prevalent in Argentina 40,41 and Uruguay, 42 and is found at a lower prevalence in Chile. 43,44
Here we report the cloning of 15 subtype G and seven subtype F functional envelope clones, amplified from plasma of HIV-1-infected individuals, which were analyzed for phenotypic properties regarding neutralization and coreceptor usage.
Materials and Methods
Samples
Most samples were collected in Spain from 2002 through 2009, except one in Chile in 2001 and one in Russia in 2004. RNA was extracted from 1 ml plasma using reagents from Nuclisens kit (Biomerieux, Marcy l'Etoile, France) following the manufacturer's instructions. Samples formed part of studies involving genotypic determination of antiretroviral drug resistance, and therefore RNA was initially amplified by reverse transcriptase polymerase chain reaction (RT-PCR) and sequenced in protease and a 0.9-kb fragment of reverse transcriptase. Those samples, which were of subtypes G, F, or recombinants containing segments of these subtypes, were selected for full-length envelope RT-PCR amplification and sequencing. Viruses of subtypes G or F in bulk sequencing of the external portion of the envelope were selected for single genome amplification (SGA) and cloning.
SGA of the envelope
Primers were designed to recognize sequences conserved within HIV-1 group M. Names, sequences, and HXB2 positions of primers are, for RT-PCR, SG3-up TACAGTGCAGGGGAAAGAATAATAGACATAATA (4809–4841) and SG3-lo AGACCCAGTACAGGCRARAAGC (9523–9544); and for nested PCR, Env-up GTTTCTTAGGCATCTCCTATGGCAGGAAGAAG (5957–5983) and Env-lo GTTTCTTCCAGTCCCCCCTTTTCTTTTAAAAAG (9063–9088) (GTTTCTT appending at 5′ ends of nested primers is intended to increase 3′ adenosine addition by Taq DNA polymerase for TA cloning 45 ). RT-PCR components included SuperScript III Reverse Transcriptase (Invitrogen, Carlsbad, CA) 4 U, BioTaq DNA Polymerase (BioLine, London, UK) 1 U, Pfu Turbo DNA Polymerase (Stratagene, La Jolla, CA) 0.1 U, RNasin (Promega, Madison, WI) 4 U, BioTaq 10 × buffer 2 μl, MgCl2 2 mM, dNTPs 0.2 pM each, primers 8 pmol each, and 2 μl RNA extract, in a 20 μl reaction volume. The RT-PCR thermal profile was 50°C 1 h; 94°C 2 min; 35 PCR cycles (94°C 15 s, 55°C 30 s, 72°C 5 min); 72°C 10 min. A 2-μl aliquot of RT-PCR product was used for nested PCR, containing the following reagents: BioTaq DNA polymerase 1 U, Pfu Turbo DNA Polymerase 0.1 U, BioTaq 10 × buffer 2.5 μl, MgCl2 2 mM, dNTPs 0.2 pM each, primers 10 pmol each, in a 25 μl reaction volume. The thermal profile was 94°C 2 min; 30 PCR cycles (94°C 15 s, 55°C 30 s, 72°C 3 min); 72°C 7 min. PCR products were visualized after 1% agarose gel electrophoresis under UV light with SYBR Safe (Invitrogen, Carlsbad, CA) staining. An SGA assay 46 was performed, with serial 2-fold RNA dilutions, assaying 8 to 16 replicates per dilution to obtain amplified products in less than 30% tubes, checking for the absence of mixed peaks in sequence electropherograms.
Envelope cloning
Bands were extracted from the gel with the MinElute Gel Extraction kit (Qiagen, Valencia, CA), minimizing UV light exposition, with 10 μl elution volume. Two microliters of DNA was used for ligation in pcDNA3.1/V5-His-TOPO TA expression vector (Invitrogen, Carlsbad, CA). MAX Efficiency Stbl2 competent cells (Invitrogen, Carlsbad, CA) were transformed with 4 μl of ligated product, seeded in LB-Amp plates, and incubated at 30°C overnight. Screening for colonies with the insert in the correct orientation was performed by colony PCR using primers recognizing the vector and the insert, respectively.
Pseudovirion production and functional testing
Envelope (300 ng) and pSG3Δenv backbone 47 (300 ng) plasmids were mixed with JetPEI (Polyplus Transfection, Illkirch, France) at a JetPEI:DNA (μl:μg) ratio of 4:1 in a volume of 100 μl. The mixture was added to 293T cells, seeded 1 day earlier at 3 × 104 cells/well in a 24-well plate. After transfection, medium was removed and replaced with 220 μl of fresh Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (DMEM + 10% FBS). Fifty microliters supernatant with pseudovirions was collected at 48 h after transfection and used to infect TZM-bl cells, 47 seeded 1 day earlier at 5 × 103 cells per well in a 96-well plate, which were assayed for luciferase activity 3 days after infection, using a Victor3 luminometer (Perkin Elmer, Waltham, MA). pSG3Δenv plasmid and TZM-bl cells were obtained through the NIH AIDS Research and Reference Reagent Program.
Sequencing and phylogenetic sequence analyses
Full-length gp160 sequences were obtained for all functional clones used in this study. The sequencing reactions were performed with an ABI Prism BigDye Terminator Kit (Applied Biosystems, Foster City, CA), with analysis in ABI PRISM 3700 automated sequencer (Applied Biosystems, Foster City, CA). Sequences were aligned with MAFFT v.6 48 and phylogenetically analyzed via maximum likelihood using the RAxML online program, 49 applying the GTR + CAT + I nucleotide substitution model. Recombination was analyzed by bootscanning using SimPlot v.3.5.1 50 . For this analysis, a window of 300 nucleotides was used, moving in 20 nucleotide increments, and trees were constructed using Kimura's two-parameter distances and the neighbor-joining algorithm.
Neutralization assay
Neutralizing antibody responses against HIV-1 Env pseudoviruses were measured using a luciferase-based assay in TZM-bl cells as previously described. 19,51 Briefly, seven 3-fold serial dilutions of monoclonal antibodies (MAbs), starting from a concentration of 50 μg/ml, were performed in duplicate (96-well flat bottom plate) in DMEM + 10% FBS (100 μl/well). Then 200 TCID50 (as determined in TZM-bl cells) of pseudovirus was added to each well in a volume of 50 μl and the plates were incubated for 1 h at 37°C. TZM-bl cells were then added (1 × 104/well in a 100 μl volume) in DMEM + 10% FBS containing DEAE-Dextran (Sigma, St. Louis, MO) at a final concentration of 11 μg/ml. Assay controls included replicate wells of TZM-bl cells alone (cell control) and TZM-bl cells with virus (virus control). Following a 48-h incubation at 37°C, 150 μl of assay medium was removed from each well and 100 μl of Bright-Glo luciferase reagent (Promega, Madison, WI) was added and luminescence was measured. The 50% inhibitory concentration (IC50) was calculated as the MAb dilution that caused a 50% reduction in relative luminescence units (RLU) compared to the virus control wells after subtraction of cell control RLUs. IC50 were calculated with five-parameter curve fitting using neutralizing antibody analysis software provided by the CAVD Vaccine Immune Statistical Center. Susceptibility of pseudoviruses to soluble CD4 (sCD4) and polyclonal HIV immunoglobulin (HIVIG) was also determined in a similar way, using 3-fold serial dilutions of these reagents, starting from concentrations of 50 μg/ml sCD4 and 2500 μg/ml HIVIG. Pseudovirus expressing the envelope of subtype B isolate 6535.3, known to be sensitive to all four MAbs used in the assay, was included as positive control. MAbs 2F5, 4E10, 2G12, and b12 were obtained from Polymun (Vienna, Austria), sCD4 protein was obtained from Progenics (Tarrytown, NY), and HIVIG was obtained from the NIH AIDS Research and Reference Reagent Program.
Coreceptor usage determination and prediction
Coreceptor usage was tested using GHOST cells, which express CD4 and either CCR5 or CXCR4, and carry HIV-2 LTR-driven GFP. Infection was done as described 52,53 with minor modifications. GHOST-parental, GHOST-CCR5, and GHOST-CXCR4 cells were seeded in contiguous alternating rows of a 96-well plate with 5 × 103 cells per well in 200 μl of DMEM supplemented with 7.5% fetal calf serum. The following day, medium was removed and replaced with 60 μl of fresh medium and a volume of pseudovirus-containing supernatant obtained from transfected 293T cells equivalent to 50 pg of p24 antigen. After overnight adsorption, virus was washed with phosphate-buffered saline and 200 μl of fresh medium was added. On day 3 after infection, GFP expression was observed by fluorescence microscopy, comparing contiguous wells of each GHOST cell line. Positive control viruses for each coreceptor were included in the assay.
Coreceptor usage prediction by genotypic methods was done by net V3 charge, 54 11/25 rule, 55 and by using Web PSSM 56 and Geno2Pheno 57 online programs.
Results
Functional assay in TZM-bl cells
One hundred and thirty-seven envelope clones derived from subtype G amplicons obtained from 21 samples and 139 envelope clones derived from subtype F amplicons obtained from 18 samples were tested for functionality in TZM-bl cells. Of the tested clones, 34 from 21 SGAs derived from 15 individuals harboring viruses with subtype G envelopes and 20 from 12 SGAs derived from seven individuals harboring viruses with subtype F envelopes were highly functional (clone/background luminescence ratio ≥50). Epidemiological and clinical data of these individuals are shown in Table 1. One clone per sample (that yielding the highest luminescence signal in the TZM-bl assay) was used for further phylogenetic analyses and phenotypic assays.
IDU, injecting drug use; HT, heterosexual contact; HO, homosexual contact; MTC, mother-to-child transmission; TFS, blood transfusion; n.a., not available; DRC, Democratic Republic of Congo.
Phylogenetic analyses
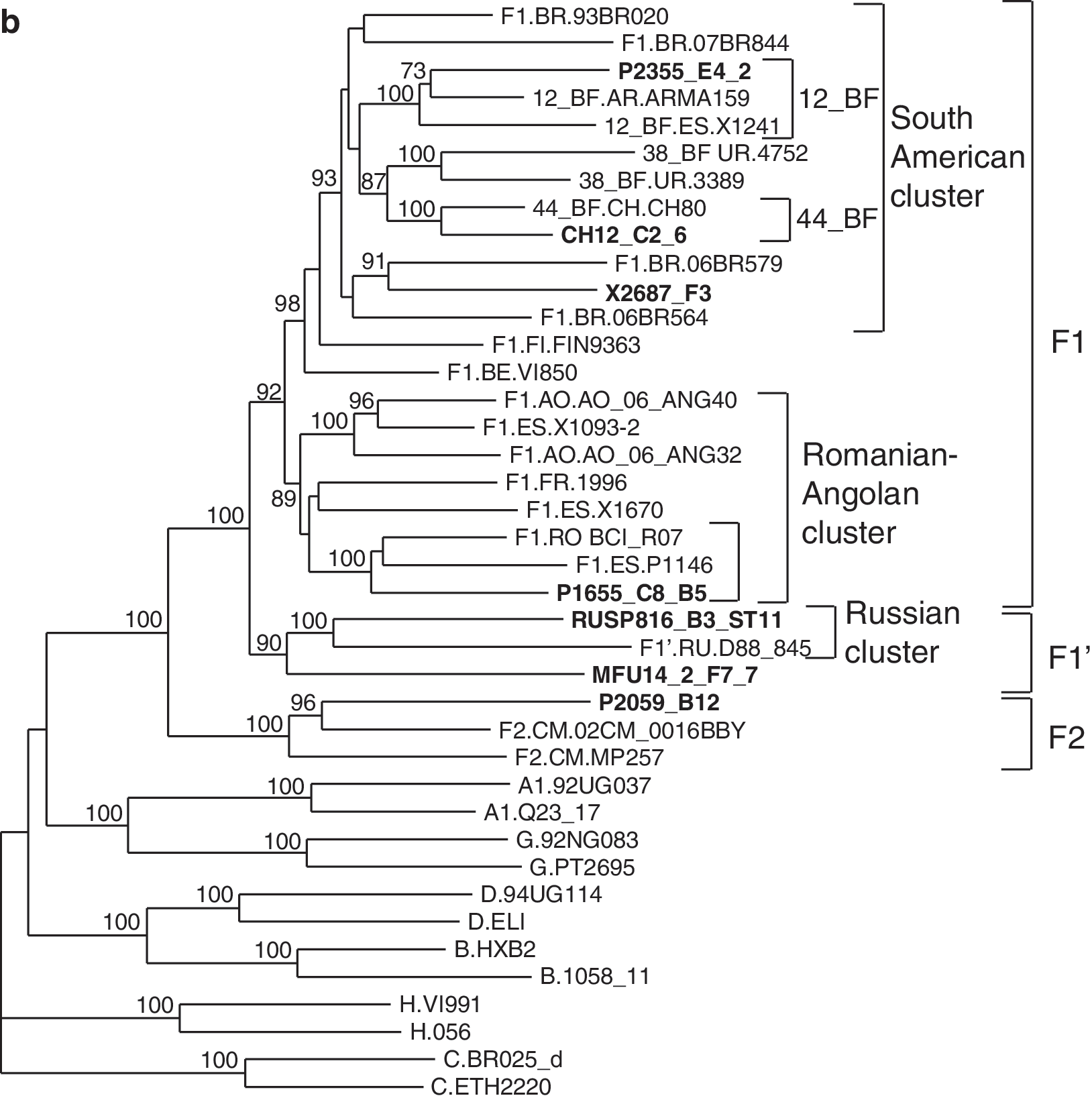
In bootscan analyses functional cloned envelopes were uniformly of subtypes G or F, except P2355, which was a BF recombinant, with a short subtype B segment (∼0.2 kb) in the intracytoplasmic domain of gp41 (results not shown). In a phylogenetic tree, 11 of 15 subtype G envelopes clustered with the Spanish–Portuguese variant, 32,33 one with the Cuban variant, 34,35 one with a predominantly Nigerian cluster, one with a Cameroonian cluster, and one in a cluster comprising one Kenyan and two Cameroonian viruses (Fig. 1a). Among subtype F envelope clones, one (P2059_B12) branched with subsubtype F2 references and the remaining six branched with subsubtype F1 references (Fig. 1b). Among these, two viruses, one from an individual from the Democratic Republic of Congo (DRC) (MFU14_2_F7_7) and one from Russia (RUSP816_B3_ST11), clustered with a reference virus from Russia forming a sister clade of the F1 subsubtype. This clade corresponds to the recently identified Russian subtype F variant, which was designated F1′, 58 to reflect both its distinctive phylogenetic position and its close relationship to proper F1 viruses; however, for the sake of simplicity, in this article F1′ viruses will be ascribed to the F1 clade. Among the proper F1 viruses, one branched with the Romanian variant and three with South American viruses, among which one (P2355_E4_2) clustered with the CRF12_BF references and another (CH12_C2_6) with CH80, a CRF44_BF virus from Chile. 44

Phylogenetic trees of full-length gp160 sequences of functional subtype G and F envelope clones. The trees were constructed via maximum likelihood using the online version of RAxML.
49
Names of envelope clone sequences are in bold type. Reference subtype G and F isolate names are preceded by the two-letter code of the sampling country. Only bootstrap values ≥70% are shown. Intrasubtype clusters comprising clone sequences are bracketed on the right.
Neutralization phenotype
Neutralization phenotype of the cloned envelopes was characterized in a pseudovirus assay using TZM-bl cells, with four broadly neutralizing MAbs, b12, 2G12, 2F5, and 4E10 (all of them derived from subtype B-infected individuals 59,60 ), TriMab (an equimolar mixture of b12, 2G12, and 2F5), sCD4, and HIVIG (polyclonal immunoglobulin from HIV-1 subtype B-infected individuals) (Table 2).
Values in bold indicate neutralization of pseudovirions.
Subtype G
All 15 subtype G-enveloped pseudoviruses were resistant to b12 and 2G12 at the highest concentration tested (50 μg/ml) and a majority (87% and 67%, respectively) were susceptible to 2F5 and 4E10 (Table 1). However, the activity of these two MAbs was relatively low for the susceptible pseudoviruses, with a mean IC50 of 14.7 μg/ml and 12.1 μg/ml, respectively, and only one pseudovirus being neutralized by 4E10 at a concentration <1.0 μg/ml. TriMab neutralized only four (27%) viruses, and this activity correlated with that of the 2F5 component. sCD4, which blocks gp120 binding to the CD4 receptor, neutralized most (87%) pseudovirions, implying that the CD4 binding site is usually accessible on the pseudotyped subtype G envelope glycoproteins. HIVIG neutralized only four (27%) subtype G-enveloped pseudovirions.
Subtype F
All seven subtype F-enveloped pseudoviruses were resistant to b12 and 2F5 at the highest concentration tested (50 μg/ml); only one (CH12_C2_6) was susceptible to neutralization by 2G12, with an IC50 of 2.88 μg/ml; and six were neutralized by 4E10, with an IC50 of 0.29–14.17 μg/ml (mean 4.68 μg/ml) (Table 1). Only one pseudovirus (CH12_C2_6) was neutralized by TriMab, which correlated with susceptibility to 2G12. sCD4 and HIVIG each neutralized six subtype F pseudoviruses.
Correlation of neutralizing activities of MAbs with epitope sequences
We examined the correlation of the neutralizing activity of each MAb with the sequences of the corresponding epitopes on the envelope clones (Table 3).
All positions correspond to the HXB2 envelope protein.
Amino acid positions correspond to sites for which mutations have been shown to result in resistance to b12 (see main text for details). Dots indicate identity with the residue(s) on top, which correspond to the subtype B consensus sequence.
Amino acid positions correspond to potential N-linked glycosylation (PNLG) sites, with sequence NX(S/T). Those whose carbohydrate chains have been shown to be the most important for 2G12 binding are in bold type. Plus and minus signs indicate the presence or absence, respectively, of a PNLG site at the corresponding position.
Residues correspond to positions 662 through 667. Residues shown to be critical for binding and neutralization are in bold type. Dashes indicate identity with the residue on top.
Residues correspond to positions 671 through 680. Residues most important for binding or neutralization are in bold type. Residues dispensable for binding and neutralization are in lower case. Dashes indicate identity with the residue on top. T676S substitution does not interfere with neutralization by 4E10. 11,14
b12
This MAb targets a discontinuous epitope on gp120 overlapping the CD4 binding site. 61,62 By site-directed mutagenesis, gp120 amino acid substitutions D185N, N276A, S364H, S365A, P369L (but not P369A), and T373R combined with N386 have been shown to result in resistance or reduced susceptibility to b12. 63 –65 Among the subtype G envelopes, all had substitutions at residue P369 (12 to L, two to V, and one to S), 13 at residue D185 (nine to N, two to T, and one each to K, G, and S), and five each at residues S364 (three to P and one to H) and S365 (four to A and one to V), with all clones but one containing at least one of the mutations reported to impact b12 activity (P369L and/or D185N and/or S364H). Among the subtype F envelopes, six had the substitution P369L, six had substitutions at residue D185 (three to N and one each to P, G, and S), and one each had the S364H and S365P substitutions, with all clones but one containing at least one of the mutations reported to affect b12 activity. (However, it should be pointed out that P369L has recently been shown to be tolerated in the context of subtype C envelopes without a major impact on b12 binding or neutralizing activity, 66 which makes uncertain the role of this mutation in b12 resistance of subtype G and F envelopes.)
2G12
This MAb recognizes a mannose-dependent epitope on gp120, with N-linked glycans attached to residues 295, 332, and 392 being the most important for binding. 67,68 Thirteen subtype G and four subtype F envelope clones lacked one or more potential N-linked glycosylation (PNLG) sites (NXS/T) at these positions, which may explain their resistance to 2G12. However, one subtype G and two subtype F 2G12-resistant clones had all three mentioned PNLG sites, with the F clones also containing two additional PNGL sites, at 339 and 397 positions, known to contribute to 2G12 binding. 68
2F5
This MAb recognizes the core epitope ELDKWA at the membrane-proximal external region (MPER) of gp41, 69 with DKW residues (at positions 664–666) being critical for 2F5 binding and neutralization activity. 70,71 Among the subtype G clones, there was a perfect correlation between mutations at the DKW motif and resistance to neutralization, with both resistant clones containing mutations at K665. Among the subtype F clones, only three had mutations at the DKW motif, and one maintained all residues of the previously defined core epitope (ELDKWA), although all were resistant to 2F5.
4E10
This MAb binds to a linear epitope at the MPER of gp41 adjacent to the 2F5 epitope, with the core sequence NWF(D/N)IT (residues 671–676). 71 Site-directed mutagenesis using an alanine scanning strategy has shown that mutations at residues W672, F673, at the core epitope, and W680, seven residues C-terminal of these two, result in resistance to neutralization by 4E10. 71 Among the four subtype G envelope clones resistant to 4E10, two had mutations at one or more of the mentioned residues. However, the three other subtype G and the only subtype F 4E10-resistant clones lacked mutations at these residues or other mutations known to decrease 4E10 binding. 72
Coreceptor usage
In vitro assays of pseudoviruses using GHOST-CCR5 and GHOST-CXCR4 cells revealed that all tested subtype G clones were CCR5-tropic, including seven from patients with CD4+ T cell counts <200 cells/ml. The results were generally consistent with four genotypic prediction methods, although for two envelopes the Geno2Pheno program predicted CXCR4 usage when using false-positive rate cut-offs ≥2.5% and ≥10%, respectively (Table 4). Regarding the subtype F envelopes, six clones were CCR5-tropic and one was dual CCR5- and CXCR4-tropic. Four genotypic methods were concordant in predicting the R5 phenotype for all seven subtype F envelopes (Table 4).
R5: CCR5. R5X4: CCR5 and CXCR4.
Calculated by subtracting negatively charged residues from positively charged ones in the V3 loop of Env: (R + K)−(D + E). V3 net charge ≥+5 is associated with CXCR4 usage. 54
Presence of basic amino acids residues at the 11 or 25 position in the V3 loop (corresponding to positions 306 and 322 in HXB2 Env) is associated with CXCR4 usage or syncytium-inducing phenotype. 55
The subtype B X4R5 matrix was used for the analysis.
R5/X4: CXCR4 tropism is predicted when using a false-positive rate cutoff ≥ 2.5% (X2486) or ≥ 10% (X2131).
Discussion
Here we report the construction of 15 subtype G and seven subtype F functional envelope clones, which were analyzed for coreceptor usage and neutralization properties. Previously, only one subtype G and two subtype F functional envelope clones, derived from in vitro cultured viruses, had been reported in the literature. 26 The newly derived clones were obtained from plasma by a single genome amplification assay, cloned into an expression vector, and tested for functionality in a pseudovirus assay using TZM-bl reporter cells. 19 All clones selected for high functionality, each from a different individual, yielded a luminescence signal at least 50 times above background levels, which renders them usable in neutralization assays.
The methods employed to obtain the functional Env clones were similar to those that have previously been described. 19,46 However, several modifications were introduced to increase the efficiency of amplification and cloning and to reduce costs. First, PCR primers were designed to recognize sequences highly conserved in group M. Second, amplification was done by one-step RT-PCR using a mixture of Taq and Pfu DNA polymerases and a small amount (1/20 that recommended by the manufacturer) of SuperScript III reverse transcriptase (this protocol was modified from the one used for amplification of hepatitis C virus RNA 73 ); this resulted in much reduced costs and a greater efficiency of amplification, compared to a published two-step RT-PCR protocol 46 (results not shown). And third, a seven nucleotide extension, GTTTCTT, was appended to 5′ ends of nested primers to increase adenosine addition to the 3′ ends of PCR products 45 ; this modification proved to be essential for high efficiency cloning.
Analysis of neutralization by broadly neutralizing MAbs using a pseudovirion assay revealed that subtype G envelopes were uniformly resistant to b12 and 2G12, targeted to gp120, and a majority was susceptible to 2F5 and 4E10, targeted to the MPER of gp41. However, the activities of these MAbs were relatively low, with pseudoviruses derived from only one envelope clone being neutralized by 4E10 at concentrations <1.0 μg/ml. These results are similar to those reported using 15 plasmids expressing subtype G Env libraries, 74 which showed low frequency (13% and 20%, respectively) susceptibility to b12 and 2G12 and high frequency (80% and 100%, respectively) neutralization by 2F5 and 4E10, although, in contrast to the previous study, susceptibility to 4E10 was not uniform among the clones tested by us. The pattern of susceptibility to broadly neutralizing MAbs observed for subtype G envelope clones resembles that previously described for subtype A 20 and CRF01_AE 24,25 clones. With regard to the subtype F clones, only 4E10 neutralized most pseudoviruses (six of seven), with 2F5 and b12 being uniformly ineffective and 2G12 being effective only against pseudoviruses derived from one clone. These results point to subtype F as the group M clade showing the widest resistance to the four broadly neutralizing MAbs raised against subtype B used in this study, since other clades are frequently neutralized by at least two of these MAbs, as shown in previous reports and in this study, with only subtype B being frequently neutralized by all four MAbs. 11,19,21 The results also show that 4E10 is the only broadly neutralizing MAb used in this study capable of neutralizing viruses across all major group M clades, including subtypes G and F, although its potency is considerably lower than that of the recently reported broadly neutralizing MAbs PG9, PG16, VRC01, and VRC02. 74,75 Previous studies have shown that most subtype C viruses are resistant to 2F5 76,11,12 and here we show that subtype F viruses are also resistant to this MAb. The MAbs targeted to gp120 show even more restricted activities, with 2G12 being frequently effective only against subtype B viruses and b12 only against viruses of subtypes B, D, and C.
Analysis of the sequences of epitopes targeted by the MAbs showed that clones containing mutations at residues known to be critical for MAb binding and neutralization were resistant to neutralization by the corresponding MAbs. However, some clones resistant to 2G12, 2F5, and 4E10 lacked known resistance-associated mutations at the epitopes (Table 3), a phenomenon previously reported by other authors. 11,12,14,20,72 This was most frequently seen among subtype F envelope clones, all of which were resistant to 2F5, although four of them lacked mutations at the DKW motif critical for 2F5 binding. 70,71 This suggests that conformational changes determined by extraepitopic substitutions may influence resistance to MAbs in these clones.
HIVIG, consisting of polyclonal immunoglobulin from HIV-1 subtype B-infected individuals, neutralized only four (27%) of 15 subtype G-enveloped pseudoviruses but six (86%) of seven subtype F-enveloped pseudoviruses. This suggests a limited B–G cross-clade neutralizing activity of sera from HIV-1-infected individuals (similar to what has been reported for other serum–pseudovirus clade pairs 12,14 ) but a relatively high degree of B–F cross-clade neutralizing activity, which could reflect the closer phylogenetic relatedness of subtype B to subtype F relative to other subtypes (except D). 77
While the subtype G and F Env pseudoviruses described here do not appear to be unusually sensitive to antibody-mediated neutralization (as compared to sensitive tissue culture laboratory adapted viruses), it will be important to more thoroughly characterize the neutralization profile of these viruses using genetically diverse HIV-1+ serum samples to assess whether they exhibit a Tier 1- or Tier 2-like phenotype. 16
The envelope clones were also tested for the usage of coreceptors for cell entry in GHOST cells. Entry of HIV-1 into target cell is mediated through the interaction of the envelope with CD4 and a coreceptor, usually the β-chemokine receptors CCR5 and CXCR4. Initial infection is almost invariably established by CCR5-tropic (R5) variants, which predominate until late stages of the infection, when CXCR4-tropic (X4) or dual-tropic (R5X4) viruses frequently emerge, concomitantly with a faster CD4+ T cell decline. It has been reported that CXCR4 coreceptor usage is common in advanced HIV-1 infection in most major HIV-1 clades, except in subtype C. 78 –80 However, data on coreceptor usage for subtype G viruses are insufficient, and were not reviewed in a recent article, 81 probably due to the relatively low number of isolates analyzed, particularly at late stages of infection. We found that all 15 subtype G envelope clones exhibited an R5 phenotype, including those from seven patients with CD4+ cell counts below 200/ml. This result is concordant with previous studies performed with subtype G isolates suggesting an overrepresentation of the R5 phenotype, which was observed in 22 of 24 viruses, including eight from Cameroon, 82,83 six from Ghana (all from AIDS patients), 84 and eight from Spain. 85 –87 One envelope clone from Uganda 26 and one isolate from Russia 88 exhibited a non-syncytium-inducing phenotype, usually associated with CCR5-tropism. Only two subtype G viruses have been reported to use CXCR4. 78,82 Thus, out of 41 analyzed subtype G viruses or envelope clones, all but two were of the R5 phenotype, including 13 from 15 patients with AIDS or CD4+ T cell counts <200/ml. The results reported here, together with those reported previously using primary viral isolates, suggest that, similarly to subtype C, 78 –80 CXCR4 coreceptor usage is uncommon in subtype G, even at late stages of the infection.
With regard to coreceptor utilization by subtype F envelope clones, we found that six exhibited an R5 phenotype and one an R5X4 phenotype. The dual-tropic envelope corresponds to a patient with AIDS, while three other AIDS patients had R5 envelopes. A literature review reveals that of a total of 55 viruses of subtype F or with a subtype F envelope whose phenotype was analyzed, 47 were of R5 or non-syncytium-inducing phenotype and eight were of X4, R5X4, or syncytium-inducing phenotype. 78,82,83,85,87,89 –92 These studies included four viruses from adult patients reported to have AIDS 82,85 and 34 vertically infected children with “serious infections,” 89,90 eight of which (three adults and five children) were of R5X4 or X4 phenotypes. Taken together, our results with envelope clones and those reported by other authors with viral isolates suggest that CXCR4 usage by subtype F viruses may be relatively common in late stages of the infection in adult individuals, although analysis of more viruses from patients with AIDS or late-stage infection would be necessary to estimate the frequency of CXCR4 utilization by this subtype in advanced disease. Four genotypic methods for coreceptor usage prediction were correct in predicting the R5 phenotype in six subtype F envelopes, but CXCR4 usage by P1655_C8_B5_1 failed to be predicted by all methods. In this regard, it should be pointed out that genotypic methods have been shown to perform poorly for predicting CXCR4 usage in non-subtype B clades 93 and that the 11/25 rule 54 failed to predict CXCR4 usage in all five Romanian subtype F X4 viruses when DNA from peripheral blood mononuclear cell cultures was analyzed and in three of five of these viruses when virion RNA was analyzed. 89,90 Our own analyses of V3 sequences from previously reported subtype F X4 or R5X4 viruses using Geno2Pheno and Web PSSM programs further confirm the low sensitivity of genotypic methods for predicting CXCR4 usage by subtype F viruses (results not shown).
In conclusion, 15 subtype G and seven subtype F functional envelope clones derived from plasma of HIV-1-infected individuals have been obtained, which could be used as references in studies on antibody-mediated neutralization, coreceptor usage, or susceptibility to viral entry inhibitor drugs. The envelopes could also be employed to construct chimeric simian–human immunodeficiency viruses for use in the macaque model and as immunogens in HIV-1 vaccines. In a pseudovirion assay all subtype G envelopes were resistant to broadly neutralizing MAbs b12 and 2G12, but were frequently neutralized by MAbs 2F5 and 4E10, a pattern resembling that previously reported for subtype A and CRF01_AE envelope clones, and all but one of the subtype F envelopes were resistant to at least three of the four MAbs, of which only 4E10 was active against most subtype F-enveloped pseudoviruses. Subtype G envelopes were uniformly of the R5 phenotype, including those from seven patients with CD4+ T cell counts below 200/ml, which, together with reported results using viral isolates, suggests that, similar to subtype C, switch to or acquisition of CXCR4 coreceptor usage along the course of infection is infrequent in subtype G, implying that CCR5 antagonist drugs are expected to be effective against most HIV-1 subtype G viruses.
Sequence Data
Sequences have been deposited in GenBank under accessions EU885759, EU885761–EU885765, FJ817368–FJ817371, GQ222684, GQ222685, GQ222687, GQ862780, GQ862781, GU332502–GU332507, GU332509, GU332513, HM068551, and HM068553.
Footnotes
Acknowledgments
We thank the personnel at the Genomic Unit, Centro Nacional de Microbiología, Instituto de Salud Carlos III, Spain, for technical assistance in sequencing; Milagros Pinilla and Ana Sánchez for assistance in sample and data processing; and Rafael Delgado and Lorena Martínez Prats for help in setting up the envelope cloning method. This work was funded through Collaboration for AIDS Vaccine Discovery/Comprehensive Antibody-Viral Immune Monitoring Consortium (Bill and Melinda Gates Foundation, award #38619). Ana Revilla and Elena Delgado contributed equally to this work.
Author Disclosure Statement
No competing financial interests exist.