
Abstract
The intestinal immune dysfunction due to loss of mucosal and peripheral CD4+ T cells in individuals with HIV/AIDS is presumably responsible for the establishment of persistent cryptosporidiosis. Simian immunodeficiency virus (SIV)-infected macaques were used to investigate the phase/timing in SIV infection, which permits a self-limiting Cryptosporidium parvum infection to become persistent in immunodeficient hosts because of significant mucosal immune defects. Two groups of SIV-infected macaques were challenged with C. parvum; one was challenged during the acute SIV infection phase (2 weeks post-SIV infection) and the second was challenged during the chronic SIV phase (CD4 counts 200–500 cells/μl of blood). Samples (fecal, blood, biopsy, and necropsy) were collected at different time points after infection to correlate the progression of disease with the immune status of the animals. All seven SIV-infected macaques challenged during the acute phase of SIV infection became persistently infected and excreted oocysts for 1–4 months. However, four of the six in the chronic SIV phase became infected with cryptosporidiosis, of which one survived 2 weeks and one became naturally infected. Sequential analysis of CD4+ in blood and intestines of coinfected macaques exhibited pronounced losses of CD4 T cells during the first 2 weeks after SIV infection, followed by transient rebound of CD4 T cells in the gut after C. parvum infection, and then a gradual loss over subsequent months. Persistent cryptosporidiosis was more consistently induced during the acute SIV phase indicating that profound viral damage to gut lymphoid tissue during the acute phase was more conducive, compared with the chronic phase, to establishing persistent cryptosporidiosis than low circulating CD4 T cells.
Introduction
C
Chronic cryptosporidiosis has also been recognized in immunodeficient rhesus macaques naturally infected with simian immunodeficiency virus (SIV). 7 The immunological alterations that result in the development of persistent cryptosporidiosis in individuals with HIV/AIDS are unclear at the present time. The inability of immunodeficient mice to eliminate C. parvum from the gut has indicated a role for CD4+ T cells and interferon (IFN)-γ. 8,9 Studies in humans have suggested an inverse relationship between the severity of cryptosporidiosis and peripheral CD4+ T cell count in individuals with HIV/AIDS. 10,11 A serious limitation of most studies of CD4 T cells during infection with HIV and SIV is analyzing lymphocytes derived from the gut mucosa or mesenteric lymph nodes. Analysis of peripheral blood mononuclear cells (PBMCs) does not accurately reflect the status in lymphoid tissues, particularly in the level of CD4 T cells in the gut mucosa.
Progress in HIV-1 and SIV pathogenesis has revealed that mucosal tissues including the gut are major sites for early host–pathogen interactions and CD4 T cell loss, 12,13 the preferred target for SIV/HIV infections. Studies in macaques have demonstrated that SIV selectively targets and destroys specific subsets of CD4+ T cells that are abundant in mucosal tissues but rare in peripheral lymphoid tissues. 14 Thus, the selective loss of intestinal CD4+ T cells from immunoeffector sites may likely explain the preponderance of opportunistic infections at mucosal sites.
The SIV macaque model of AIDS has provided an opportunity to examine CD4+ T cell activity simultaneously in various immunological compartments and tissues at different phases of SIV/C. parvum infection, something that cannot be performed in humans. Such observations made in the rhesus macaque model of SIV/AIDS, we predict, may shed light on the phase of SIV infection that makes immunodeficient animals more susceptible to contract opportunistic infection. To gain insight into the relationship of CD4+ T cell depletion during SIV infection and the establishment of persistent C. parvum infection, longitudinal biopsy samples of jejunum, ileum, and colon as well as peripheral blood samples were collected and analyzed during both acute and chronic stages of SIV infection, prior to and following C. parvum infection.
Materials and Methods
Animals
Of 16 C. parvum-negative macaques, as determined by Western blotting, 14 were injected intravenously with the simian immunodeficiency virus, SIVmac239 (50 ng of p27 viral-antigen equivalent), and divided into two groups of seven each. Group 1 was challenged at 2 weeks, during the acute SIV infection phase, with 107 oocysts of C. parvum. In group 2, one animal died before challenge with C. parvum, five were challenged during the chronic SIV infection phase, when the CD4 T cell counts dropped to 200–500 cells/μl of blood, and one acquired cryptosporidiosis naturally. Two SIV-naive C. parvum-seronegative macaques were also challenged with 107 oocysts of C. parvum as controls.
In addition, to study the role of preexisting antibodies, three additional SIV-naive C. parvum-seropositive macaques were challenged with C. parvum prior to SIV infection and again rechallenged with C. parvum 2 weeks after SIV infection. All animals were monitored for clinical symptoms of AIDS, diarrhea, and oocyst excretion in feces.
Animals were housed at the New England Regional Primate Research Center (NEPRC) and were maintained in a centralized biosecurity-level (BSL)-3 animal-containment facility, in accordance with the Guide for the Care and Use of Laboratory Animals. Clinical procedures and administration of appropriate anesthesia and analgesics were performed under the direction of a veterinarian. If the veterinary staff considered it to be necessary, rhesus macaques were euthanized in accordance with the recommendations of the American Veterinary Medical Association Panel on Euthanasia. All procedures and protocols were approved by the Institutional Animal Care and Use Committee at Tufts University and The Harvard Medical Area Standing Committee on Animals.
Collection of samples
Peripheral blood lymphocytes (PBL) were collected on a monthly basis from each animal to monitor the blood CD4+ cell count before and after SIV infection. Sera were collected weekly after C. parvum challenge, and feces were collected daily for 3 weeks, and weekly thereafter. Oocyst shedding was determined by microscopic examination of stained fecal smears. 15 Cryptosporidium-specific serum antibody responses were determined using an immunoblotting technique.
Endoscope-guided intestinal pinch biopsies of jejunum and colon were obtained prior to and at 2 weeks, 8 weeks, and monthly after C. parvum inoculation of SIV-infected macaques for histology and for flow cytometric analysis of intestinal lymphocytes. At necropsy, segments of jejunum, ileum, and colon were collected for histology and for the isolation of lymphocytes from the intestinal epithelium and lamina propria.
Intestinal lymphocytes and immunophenotypic analysis by flow cytometry
At necropsy, segments of 6–8 cm from the jejunum, ileum, and colon were collected from euthanized animals, and intraepithelial lymphocytes (IEL) and lamina propria lymphocytes (LPL) were isolated. 16 Briefly, IEL were isolated from intestinal segments by using EDTA and mechanical agitation, and LPL were isolated from the remaining intestinal pieces by using collagenase. Liberated cells were washed with RPMI-1640 and isolated through a 35–60% isotonic discontinuous Percoll (Sigma) density gradient. Lymphocytes were found to band at the interface between the 35% and 60% gradients. For endoscopic biopsy, samples, obtained from the proximal jejunum and distal colon at different time points of infection, were similarly processed using EDTA and collagenase, but cells (IEL and LPL) derived from these very small samples were pooled for analysis. In all cases, isolated lymphocytes (1×106) were stained as recommended. Peripheral blood (PB) was stained by a whole blood lysis technique for phenotypic analysis of PB samples, and incubated with monoclonal antibodies for 30 min at 4°C, followed by 7 min lyses with FACS lysing solution. Cells from blood and intestinal samples were then washed and resuspended in 2% paraformaldehyde for four-color flow cytometry analysis.
The isolated lymphocytes were stained with murine antihuman monoclonal antibodies to CD3, CD4, CD8, CD20, CD2, and CD69 conjugated to fluorescein isothiocyanate (FITC) or to phycoerythrin (PE) or to allophycocyanin (APC) and to peridinin chlorophyll protein (PerCP). Controls consisted of appropriate unstained and irrelevant isotype-stained samples as well as single-color-stained samples to verify the staining specificity of experimental antibodies. Data were acquired by using a Vantage flow cytometer and analyzed by CellQuest software.
Results
C. parvum infection of SIV-naive macaques
The Western blot technique was optimized to identify C. parvum-seronegative animals used for these experiments. The course of C. parvum infection was established in two SIV-naive rhesus macaques challenged orally with C. parvum. Animals became asymptomatically infected and oocysts were detected 4–5 days later with excretion lasting 10–12 days as determined by modified acid-fast (MAF)-stained fecal smears, which was considered as an acute but asymptomatic C. parvum infection. These two animals also provided the basis for the absolute CD4+ T cell counts in the blood (900–2500 cells/μl of blood), consistent with published values. 17,18
C. parvum infection of macaques during acute SIV infection
All seven macaques, orally inoculated with C. parvum 2 weeks after SIV challenge, became persistently infected and continued to excrete oocysts for several weeks (Table 1). SIV-infected macaques that excreted oocysts over 14 days are considered to be chronically infected. As previously demonstrated, SIV caused profound and selective depletion of CD4+ T in the gut-associated lymphoid tissue (GALT) within the first 2 weeks of viral inoculation, much earlier than a decline could be detected in the peripheral blood, 19 indicating that persistent cryptosporidiosis in SIV-infected macaques can be effectively established within 2 weeks following challenge, during the acute viral phase. However, only two of the seven animals showed signs of severe diarrhea, which undoubtedly contributed to the rapid deterioration of health and development of AIDS requiring euthanasia. The peripheral blood absolute CD4 counts of these animals at the time of C. parvum challenge varied from ∼380 to 700 cells per μl of blood (Table 1). Oocyst excretion in the acutely SIV-infected macaques did not correlate with the peripheral blood CD4 counts.
SIV-infected macaques were challenged with C. parvum (with 107 oocysts) 2 weeks following SIV infection.
C. parvum infection of macaques during chronic SIV infection
Of the seven SIV-infected macaques in the chronic group, one died before the C. parvum challenge due to AIDS (data not shown), five were challenged with C. parvum when their circulating CD4+ cell count dropped to 200–500 cells/μl of blood, and one acquired cryptosporidiosis naturally. Of the five macaques, three of which were challenged when their CD4+ counts dropped below 300 cells/μl of blood, all developed cryptosporidiosis (animals 10, 11, and 12 in Table 2). Two of them continued to excrete oocysts for 16–28 months, and the third did for only 2 weeks before succumbing to AIDS (Table 2). All three developed severe diarrhea and lost weight before euthanasia. Two remaining macaques (8 and 9) challenged with C. parvum when their CD4+ counts were close to ∼500 cells/μl of blood excreted no oocysts and had no diarrhea despite repeated parasite challenges. Animal #13 acquired natural cryptosporidiosis at the terminal stages of its SIV infection, when the CD4 count was ∼300 cells per μl, excreting oocysts for about 10 days before euthanasia. Curiously, this animal was excreting C. hominis, the human species (data not shown); this is the first reported natural case of a macaque infected with the human species.
SIV-infected macaques were challenged with C. parvum when their CD4 T cells dropped <211–500 cells/μl of blood.
SIV-infected macaque acquired natural cryptosporidium infection (Type 1 isolate).
n/a, not applicable, as macaques acquired natural C. parvum infection.
Challenge of C. parvum-seropositive SIV-infected macaques
Three SIV-naive macaques that had preexisting C. parvum antibody showed no evidence of infection after challenge with the parasite. The same three C. parvum-seropositive animals when challenged with C. parvum 2 weeks following SIV infection became infected and excreted oocysts for 5–18 days but with no evidence of diarrhea, indicating that while previous exposure to C. parvum is protective, this protection is markedly reduced during the acute phase of SIV.
C. parvum-specific antibodies
SIV-naive macaques challenged with C. parvum developed serum IgM, IgG, and IgA against major immunodominant parasite antigens (p900, p23, and p15) within 2 weeks, with the IgG isotype against p900>p23>p15 being predominant. The antibody responses disappeared at 8 weeks postinoculation (Fig. 1A). There was only a limited antibody response of the IgG isotype to the p23 parasite antigen in SIV-infected macaques challenged with C. parvum (Fig. 1B). This limited response of C. parvum-specific antibody in SIV-infected macaques could be due to severe depletion of CD4+CD8− single-positive and CD4+CD8+ double-positive T cells.
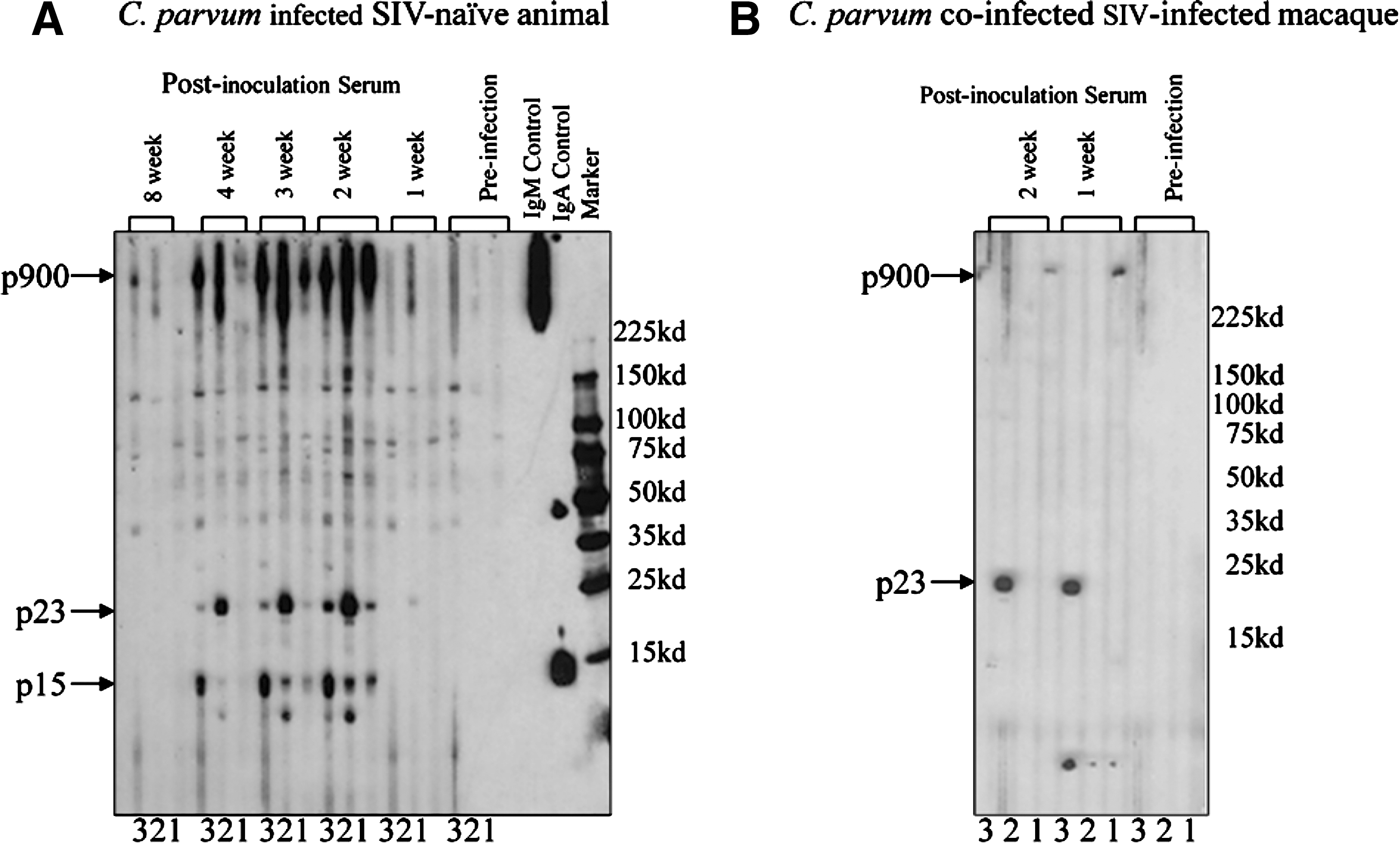
Immunoblot profiles of serum samples obtained from (
Histological analysis at necropsy
The histological findings of SIV-infected macaques with persistent cryptosporidiosis are as described in humans with AIDS, with parasite forms present in the gallbladder, bile, pancreatic duct, liver, and trachea with severe inflammation and mucosal hyperplasia. Moderate diffuse lymphoplasmacytic and eosinophilic inflammation with intraluminal parasite forms was observed in the jejunum, ileum, distal colon (Fig. 2), and rectum in euthanized macaques. The common bile duct and gallbladder showed cystic hyperplasia and the liver showed multifocal suppurative cholangiohepatitis. There was blunting and fusion of the villi and tortuous crypts.
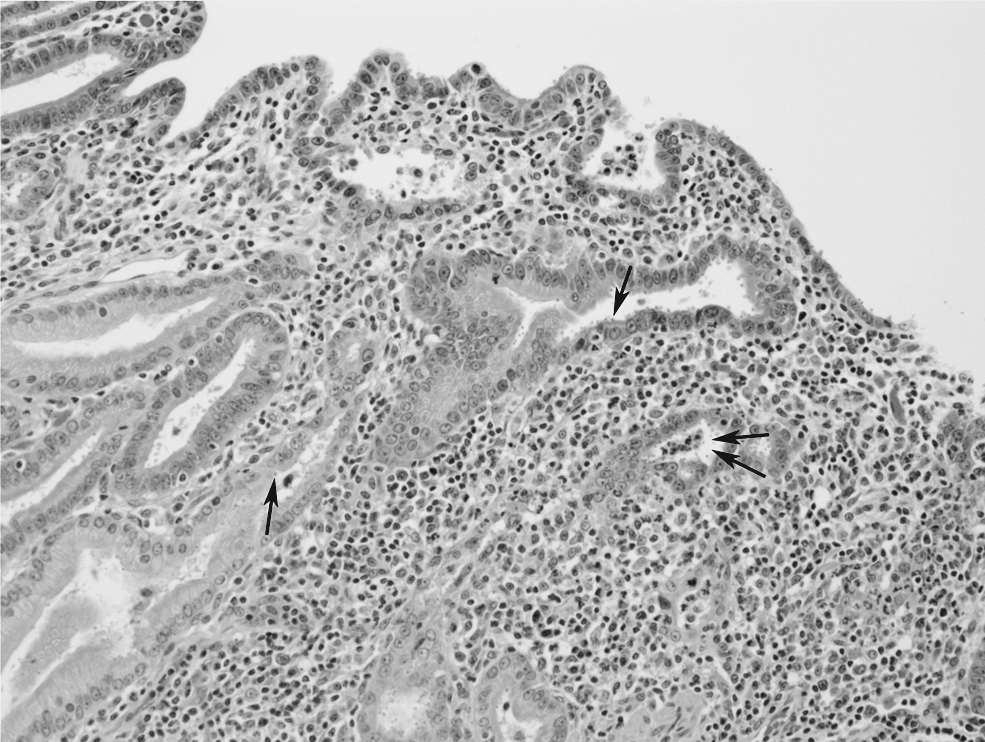
Micrograph of a colonic section from SIV-infected macaque (#10; Tables 2, 4, and 5) that was euthanized due to severe AIDS. It developed chronic cryptosporidiosis after experimental challenge. Note extensive inflammatory cell infiltration in the lamina propria of an area obliterated by crypts. Parasite forms can be seen attached to flat cell surfaces and within crypts (arrows).
Immunophenotypic analysis of PB and gut lymphocytes of SIV-naive or SIV-infected macaques with cryptosporidiosis
The prevalence of T cell subsets was analyzed in blood and intestinal tissue prior to and after SIV infection and then following C. parvum challenge at both acute and chronic SIV infection phases by using four-color flow cytometry:
C. parvum challenge during acute SIV infection
SIV challenge in this group led to a drastic drop in CD4 T cells in blood, jejunum, and colon biopsy samples, with depletion more pronounced in the gut (jejunum and colon) than in PB within 2 weeks postinfection (Table 3). After exposure to C. parvum, restoration of both CD4+ single or CD4+CD8+ T cells in PB and in jejunum and colon, especially at 2 weeks after C. parvum challenge, was observed, followed by a gradual decline until euthanasia. There was a corresponding increase in CD8 cells, as the CD4 cell population continued to decline. No further significant changes in CD8 cells was observed after exposure to C. parvum. Depletion of CD4+ T cells in the gut during the acute SIV phase presumably contributed to the establishment of the persistent cryptosporidiosis observed in the SIV-infected macaques.
The percentage values represent the proportion of gated lymphocytes. The percentage population of lymphocytes represents the mean of four (animal #4–7) animals (± SEM).
C. parvum challenge during the chronic SIV phase
SIV-infected animals were challenged at the chronic phase of infection, when their CD4+ T cells reached 200–500 cell/μl of blood (normal values are ∼900–2500 cells/μl of blood). The progression of SIV infection to achieve target CD4 levels prior to C. parvum challenge (from 8–47 weeks in two animals to 117–172 weeks in three animals) and the survival frequency after challenge were highly variable among this group of animals, which makes it difficult to study immunological parameters in a controlled manner. Of the five SIV-infected macaques challenged with C. parvum, animals #10 and 12 developed a typical chronic C. parvum infection and excreted oocysts for 28 and 16 months, respectively. While the trend in terms of the T cell flow data analysis was similar, timing was very different. We have consequently selected #10 (Table 4), which survived 28 months before euthanasia, to demonstrate a likely trend in a chronically infected host.
The data, represented in percentage population, were generated by gating through lymphocytes and then through different subsets (CD4 and CD8) of lymphocytes at different time points prior to and after C. parvum coinfection of SIV-infected macaques.
Prior to SIV infection, the percentile of CD4 T cells in PBMCs was slightly lower than the normal range (45–60%) observed in other macaques. 17,18 Similarly, mucosal CD4 lymphocytes had relatively lower CD4+ T cells (27%) and higher CD8+ T cells (58%) as compared to PB lymphocytes (Table 4). Prior to C. parvum infection, the CD4+ T cell percentage was depleted for CD4+ T cells in PB (from 38% to 11%) and for intestinal mucosa (from 27% to 5% in jejunum and 12% to 3% in colon biopsy samples). This was true for double-positive (CD4+CD8+) T cells in intestinal mucosa (Table 4). The persistence of CD4+ T cell depletion in intestinal mucosa during chronic SIV infection was adequately reflected in the PB compartment as well. There was no obvious change in the proportion of peripheral CD8+ T cells and B cells (data not shown) in PB after chronic SIV infection but a slight reduction was noticed in the gut mucosa (Table 4).
Two weeks after challenge with C. parvum, the percentage of the CD4+ T cell subset in the mucosa of the jejunum was increased from 5% to 11%, followed by a further increase to 45% after 8 weeks postinoculation. However, at 8 weeks postchallenge, the percentage of CD8+ T cells dropped from 68% to 35%, which coincided with an increase in the percentage of CD4+ from 11% to 45% in the chronic SIV macaque model. Although a slight increase in the CD4 percentage was observed in the colon, no such increase was observed in PB after C. parvum coinfection. This increase in CD4+ populations was followed by a gradual decline in the percentage of CD4 in the circulation and in the gut (jejunum and colon) until euthanasia. There was a gradual decline in the level of CD8+ T cells in the circulation and in the gut from 3 months until euthanasia (Table 4). No restoration of either CD4+CD8− single-positive or CD4+CD8+ double-positive T cells in the gut was observed at later time points following C. parvum infection.
CD4 T cells in the gut (IEL and LPL) and in the circulation at necropsy
CD4+ T cells in the blood, lymph nodes, and IEL and LPL of duodenum, jejunum, and colon were severely depleted, varying from 0.1% to 3.0% in IEL and 2% to 5% in LPL from the jejunum, ileum, and colon (Table 5), which was significantly lower than the normal macaques (15–20% in IEL and 40–45% in LPL). 20 Similarly, CD4+ T cells in the circulation and lymph nodes were less than ∼30–40% that of SIV-naive macaques (50–60%). 20 Severe depletion of CD4+ cells was apparent in the circulation and in the gut during the acute and the chronic phases of SIV infection, especially at the terminal stages.
Samples were collected from SIV/C. parvum coinfected macaques immediately after euthanasia. Portions of lymph nodes and sections of jejunum, ileum, and colon were collected fresh in complete RPMI media for lymphocyte isolation.
PBMC, peripheral blood mononuclear cells; MLN, mesenteric lymph node; PLN, peripheral lymph node; IEL, intraepithelial lymphocytes; LPL, lamina propia lymphocytes.
Discussion
We investigated the nature and time course of the progressive immune dysfunction observed in the gut mucosa of macaques infected with SIV with a view to determine the parameters at which cryptosporidiosis, an acute enteric infection in the immunocompetent host, become persistent and life-threatening in the immunodeficient host.
The SIV/AIDS macaque is an established model that exhibits many of the characteristics seen in humans with HIV/AIDS. 21 Naturally occurring cryptosporidiosis has also been described in macaques, with and without causing diarrhea. In SIV-infected or naive animals, as in humans, infections are attributed to either the human isolate C. hominis or the mammalian C. parvum. As expected, the outcome of experimental infections with C. parvum in our investigation was variable, and reflects the lack of accurate history, uniform genetic background, and potential earlier exposures despite rigorous serological evaluation. This, combined with using a relatively limited number of animals of different ages and origins, provides at best a trend, if not quite conclusive results. The individual variations observed in terms of clinical response, nature, duration, and outcome of illness precluded any attempt to combine the data for a meaningful statistical analysis.
However, the data strongly indicate that macaques challenged with C. parvum at the acute SIV phase (2 weeks post-SIV infection) are more likely to develop persistent cryptosporidiosis than animals at the chronic SIV phase, when circulating CD4 T cell are lower. While all seven animals became infected with C parvum during the acute phase (Table 1), only three of the five animals having circulating CD4 T cells ≤300 developed chronic cryptosporidiosis; two animals with CD4 T cells ∼500 showed no evidence of infection despite repeated challenge with C. parvum (Table 2). As with individuals with HIV, a higher CD4 count in SIV-infected macaques was not associated with clinical disease, while a low CD4 count (below 300 cells/μl of blood) was more likely to manifest chronic symptomatic C. parvum infection.
The rapid and early depletion of T cells in the gut described perviously 20 was presumably the key factor permitting an early establishment of persistent C. parvum, the nadir of which was reached 14–21 days after SIV infection. 22 Our results confirmed that in SIV-infected macaques major immunological perturbations occur in the gut mucosa, long before significant abnormalities are detected in the circulation. It was shown that the profound and rapid CD4+ T cell loss is selective for specific subsets of activated memory CD4+ T cells that coexpress CCR5. 20,23,24 The majority of them reside in the intestinal mucosal surfaces of humans and macaques 20,23,25 but not in the PB, lymph nodes, and the spleen and, therefore, are not susceptible to infection with most strains of HIV/SIV. 20
Our studies demonstrate that SIV-infected macaques, when challenged either during the acute (2 weeks post-SIV infection) or during the chronic SIV infection phase (<211–500 CD4/μl of blood) developed chronic cryptosporidiosis (oocysts excreted over 14 days after challenge), whereas in SIV-naive seronegative macaques, only acute cryptosporidiosis was observed, indicating that the CD4 lymphocyte status in the gut mucosa is as, if not more, critical than the peripheral CD4+ level. Such a profound and relatively rapid depletion of intestinal CD4+ T cells has also been demonstrated in HIV-infected human. 26 –29
The qualitative increase in CD4/CD8 T cell ratio at 2 and 8 weeks following C. parvum infection of SIV-infected animals indicated that CD4+ T cells may be an important effector cell type combining to resolve acute cryptosporidiosis. The markedly increased susceptibility of macaques to cryptosporidiosis during the acute phase of the SIV infection, while the innate and the adoptive arms of the immune response are still intact, reflects the significance of the T cells in protecting the gut against opportunistic infections. 22 The subsequent increases in T cell subsets after the acute SIV phase were still incapable of clearing an established chronic cryptosporidiosis. As SIV/AIDS progresses, T cell depletion in the gut and in the circulation intensifies, and animals become more susceptible once again to cryptosporidiosis and to other opportunistic infections. In addition, the challenge experiments in macaques with C. parvum indicated the secondary role played by preexisting specific serum antibodies in protecting the host against cryptosporidiosis, as seropositive animals did not excrete any oocysts and showed only limited infection when rechallenged after acute SIV infection. This could be due to functional defects or depletion of memory CD4+ T cells after infection, which might lead to a significant decrease in the antibody responses to C. parvum in SIV-infected macaques as observed in this study. Several functional deficiencies, such as memory function, were described in CD4+ T cells in the early stages of infection with HIV, long before the onset of CD4 T cell depletion was observed. 30
Intestinal IELs and LPLs also showed major changes in terms of CD4 T cell depletion, exclusively in LP of the intestinal tract (jejunum, ileum, and colon). The results showed that the CD4+ T cell depletion was accompanied by an increase in absolute numbers of CD8+ T cells. There were minor changes in the peripheral blood from these animals at the same time points. Because different lymphoid trafficking patterns exist for peripheral and mucosal lymphoid compartments, the regional immunological changes would not be detected in the peripheral circulation. This could be the reason why certain opportunistic infections that occur in AIDS patients (e.g., M. Avium, cryptosporidiosis, candidiasis, microsporidiosis, pneumocystosis) are primarily diseases of mucosal surfaces. These diseases induce further cellular activation, as observed in the rapid increase in CD4 T cells after C. parvum challenge, specifically in mucosal tissues, which could lead to increased susceptibility to infection with SIV/HIV and might accelerate the pathogenesis of AIDS and associated opportunistic infections.
Furthermore, several structural and functional alterations occur in the intestine during HIV infection. 31 –37 Similar changes were also observed in SIV macaques coinfected with C. parvum. They probably are a consequence of immunological alterations in the mucosal environment. It is well established that the immunological perturbations in the intestinal mucosa result in marked changes in intestinal architecture and function. 38 –41 Such alterations could lead to diminished immune responsiveness and the establishment of the persistence of opportunistic infections, as was shown in this study.
In brief, C. parvum infection during the acute phase of SIV infection resulted in a 100% infection rate and persistent cryptosporidiosis in all the challenged animals. Since macaques were infected shortly after SIV infection, there was less probability of acquiring natural C. parvum infection over the 2-week period. Challenging macaques 2 weeks after SIV infection had the benefit of enabling us to investigate different parameters in the SIV model well before the onset of AIDS, often initiated by other opportunistic infections. This was reflected in a clustering of the onset of oocyst excretion in the feces. Furthermore, the trend observed at the T cell level in the gut was similar to that found in those challenged during the chronic SIV phase. In contrast, C. parvum infection during the chronic SIV phase resulted in 20% infection with a variable infection rate. It took many months for SIV-infected macaques to achieve a target circulating CD4 T cell count below 500 cells/μl of blood during which some animals died due to SIV/AIDS-related complications prior to C. parvum infection. Since it was difficult to establish persistent C. parvum infection in SIV-infected animals until their CD4 counts reached>300 cells/μl of blood, animals in the chronic phase were close to their terminal stage of SIV/AIDS, which did not provide enough time to study different immunological parameters in a timely and controlled manner. This longer period prior to C. parvum infection also makes SIV-infected macaques more susceptible to acquiring other natural, often inapparent, opportunistic infections, which complicate such studies.
We believe the acute SIV macaque provides a model to closely investigate gastrointestinal infections associated with AIDS including C. parvum with a view to determine and characterize the primary effector function of mucosal CD4+ T cell immunity, which would facilitate the evaluation of immunotherapeutics to control gut infections in immunodeficient hosts.
Footnotes
Acknowledgments
This work was supported by NIH award—PO1DK55510, and RO1RR13537. We would like to thank the animal attendants who provided care for these animals for prolong periods of time, and to the pathology team who performed the necropsies and provided the histological and microbiological results.
Author Disclosure Statement
No competing financial interests exist.