
Abstract
HIV-1 budding requires short peptide motifs in p6Gag, known as late domains, that promote the release of infectious virions. The primary late domain of HIV-1 is a Pro-(Thr/Ser)-Ala-Pro (hereafter referred to as a PTAP) motif. This motif may be completely or partially duplicated. In this work we analyzed p6Gag sequences from 547 isolates from drug-naive patients and 213 isolates from patients failing HAART therapy. Complete duplications within PTAP were selected during HAART therapy in all HIV-1 subtypes analyzed: B (p = 0.0338), F1 (p = 0.0294), and C (p = 0.0001). Nevertheless, the patterns of these duplications were different; subtype C isolates accumulated longer duplications and displayed a higher frequency of duplications in both treated (54%) and drug-naive isolates (23%). Accumulation of PTAP duplications within subtypes B, F1, and C during therapy suggests a potential role of the duplications in antiretroviral drug resistance.
T
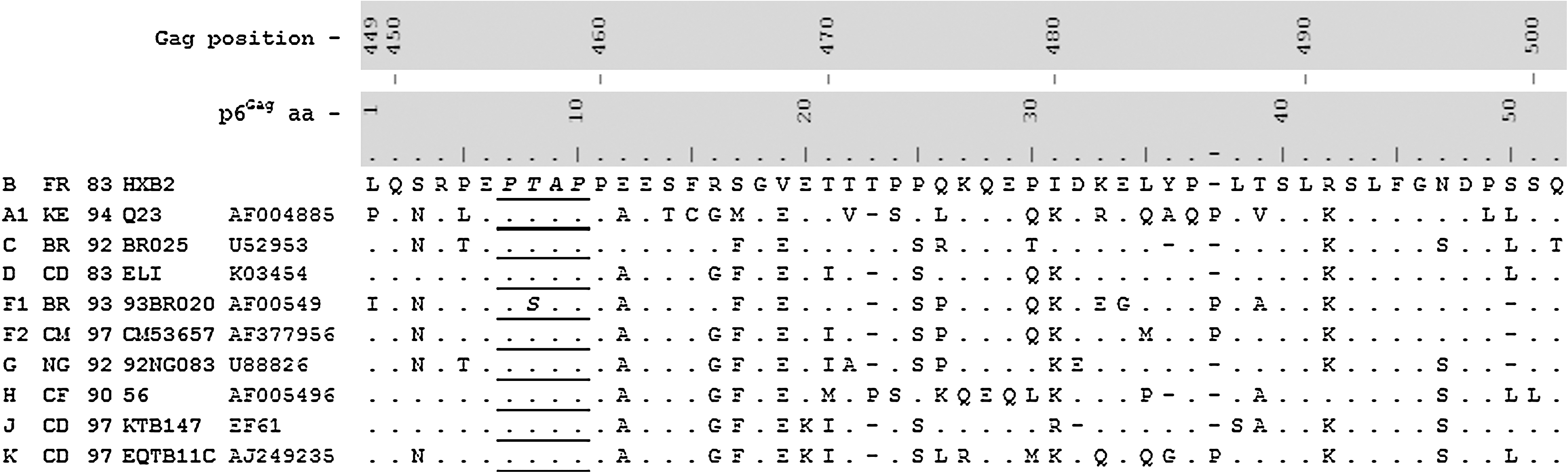
Amino acid sequences of p6Gag protein from reference sequences of different subtypes. Amino acid positions in the p6Gag and Pr55Gag precursor are shown in relation to the HXB2 reference sequence of subtype B.
Partial and complete duplications of the PTAP late domain are often seen in clinical isolates from drug-naive patients, and at an even higher frequency in isolates from patients on antiretroviral therapy. 6 The relationship between the evolution of these duplications and protease inhibitor resistance (PIR) mutations is not yet fully elucidated. Also poorly characterized is the profile of PTAP duplications among different HIV-1 subtypes. Recent studies have reported a higher prevalence rate for PTAP duplications than initially reported (10–20% vs. 5.4%) and some studies have also noted a slightly higher prevalence in ARV-experienced patients (15–36%), suggesting that ARV treatment may support the appearance/selection of such duplications. 6 –9 Recently, it was reported that the presence of PTAP duplications was linked to prior exposure to NRTIs and that partial duplication of this motif was associated with a 2-fold loss of phenotypic susceptibility to these drugs. 8 This PTAP duplication-associated resistance was also observed to be linked to the protease (PR) inhibitor amprenavir. 10 Moreover, insertions leading to the duplication of the PTAP motif in treatment-naive viral isolates were associated with high replication capacity. 11 In this work, we present strong evidence that duplications within this domain are selected by therapy and are influenced by the viral subtype.
p6Gag sequences from 213 HIV-1 isolates were obtained by direct sequencing using the ViroSeq kit assay in an ABI3100 Sequencer machine. In Brazil, the ViroSeq kit was the selected genotyping method for analyzing HIV-1 ARV drug resistance in isolates from patients failing highly active antiretroviral therapy (HAART), in the official Brazilian Ministry of Health HIV-1 drug resistance network (RENAGENO). Isolates represent different states and city locations. The DNA fragment generated by the ViroSeq kit after polymerase chain reaction (PCR) encompasses the SP1-NC-SP2-p6 region located upstream of the PR coding region. The sequencing reaction uses a reverse primer specific for the 5′ region of the reverse transcriptase (RT) coding region, named primer F, which also sequences the p6Gag region.
To recover p6Gag sequence information and evaluate the frequency of PTAP duplications, all primer F sequences were manually edited in SeqMan software (DNASTAR Package). Restricted criteria to exclude sequences without good chromatogram quality were adopted to avoid any errors. Duplications within the PTAP motif of p6Gag were annotated and analyzed by subtype, and their frequencies compared to HIV-1 p6Gag sequences from treatment-naive individuals compiled in the Los Alamos National Laboratory HIV sequence databank (
We were able to recover 213 sequences from single subtype genetic profiles in the p6Gag from patients failing HAART (Fig. 2). From this total, 118 samples were subtype B (55.4%), 61 samples were subtype C (28.6%), and 34 were subtype F1 (16%). Complete and partial (APP or PTA) PTAP duplications were found in 9.3% and 10.1% of subtype B samples, respectively, whereas in subtype C their frequencies were 54% and 0%, respectively. For subtype F1, complete and partial duplications of the late domain occurred with frequencies of 17.6% and 0%, respectively.

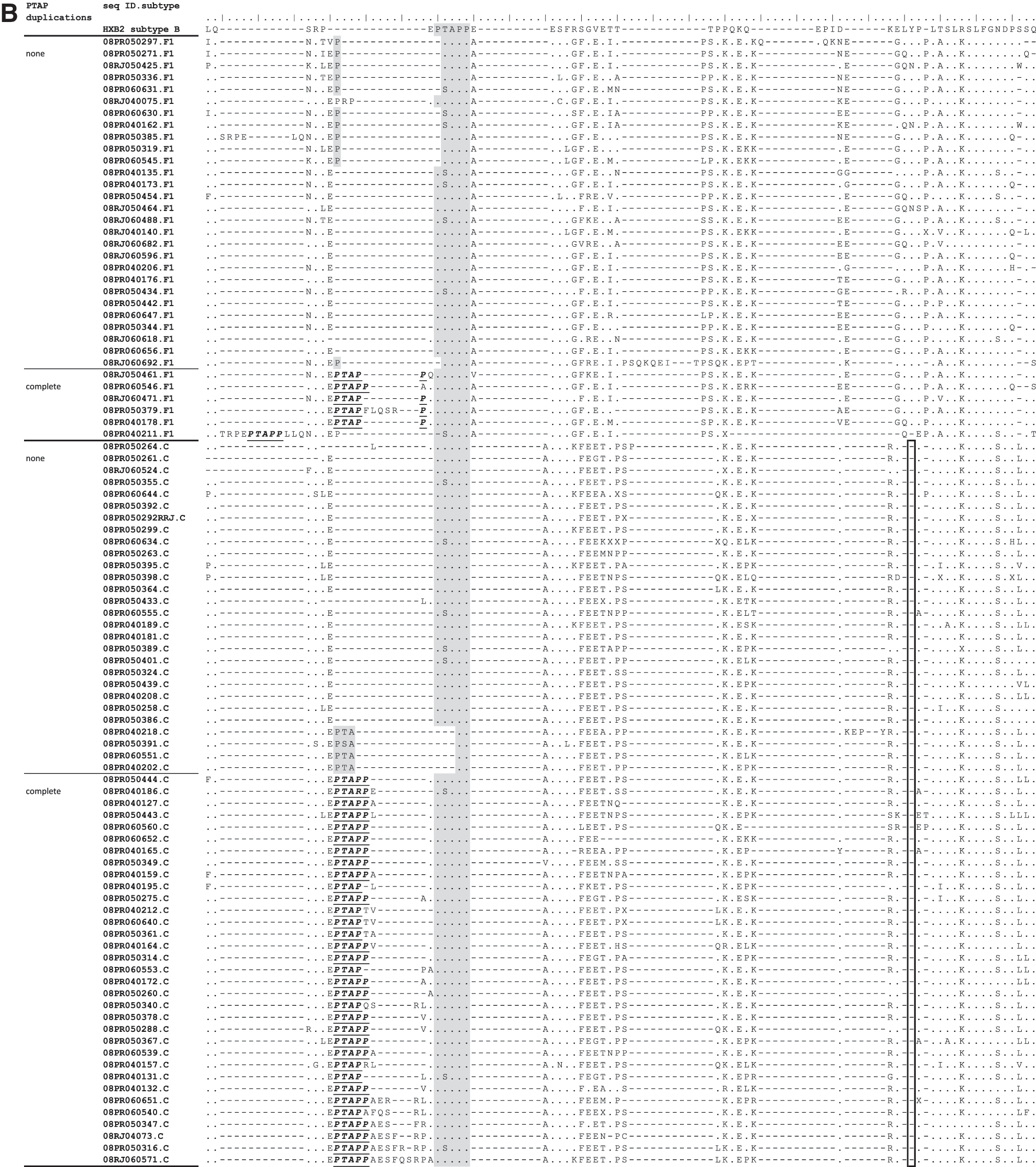
Sequence alignment of p6Gag of ARV drug-resistant isolates from (
Complete and partial duplications were found in isolates from treatment-naive individuals with frequencies of 3.7% and 8.8%, respectively, for subtype B, 23% and 1.3% for clade C, and 0% for subtype F1 (Fig. 3). These results provide evidence that the type and the length of PTAP duplications are dramatically different between subtypes B and C. Subtype C virus from patients with treatment failure presented no partial duplications, and the largest duplications were found in this subtype: 11.4% of the duplications had more than nine amino acids in contrast with subtype B and F1 with 1.6% and 5.8%, respectively, bearing large (more than nine amino acid) duplications. The largest observed duplication was 14 amino acids in length. The accumulation of complete PTAP duplications was increased 2.3- to 2.5-fold in subtype B and C when comparing drug-naive with treatment-failure isolates. Although there were few subtype F1 isolates in our analysis, we could observe the accumulation of complete PTAP duplications from 0% of isolates from drug-naive to 17.6% from treatment failure (p = 0.0338) patients. These results revealed a tendency for HAART treatment to select viral isolates carrying these duplications.

Frequency of complete duplications of PTAP late domain found among subtypes F1, B and C isolates from ARV drug-naive patients versus isolates from patients failing HAART therapy.
Duplications of the highly conserved domains KQE and SPRE were found in 0.8% (1/118) and 5% (6/118), respectively, of subtype B treatment-failure isolates; 0.6% (2/294) and 2.3% (7/294) in subtype B drug-naive isolates; 0% (1/228) and 1.75% (4/228) in subtype C drug-naïve isolates; and finally in subtype F1 2.9% (1/34) and 2.9% (1/34). None of these duplications was found in subtype C isolates from patients in treatment failure. Subtype C isolates do not carry the complete KELY or LYP domain, which lacks a tyrosine residue at position 36 (del36Y; Fig. 3B). In this study we also identify an unusual duplication pattern involving the central portion of p6Gag, present in four samples from subtype B and one from subtype F1 (3.3% and 2.9% of the treatment failure group). These duplications share a core of amino acids, TPSQ. The four different patterns found in subtype B were TPSQ, TITPSQ, KQGTTTPSQ, and TITPSQTITPSQ, and duplications TTPSQKQE and ITPSQKQE in subtype F1. There is no information to date concerning the relevance of these duplications, particularly in subtype F1 where the TPSQ and KQE duplications occurred in combination. In this study there was no significant difference in the number of PIR mutations between isolates from either clade (B and C) whether or not they carried PTAP duplications.
The difference in overall frequency of complete PTAP duplications among subtype F1 (17.6%; 6/34), subtype C (54%; 33/61), and subtype B (9.3%; 11/118) isolates from patients failing therapy may reveal a preferential use of this genetic polymorphism as an alternative mechanism for conferring ARV drug resistance in subtype C viruses. These results are strongly significant by statistical analysis; p = 0.0001 and p = 0.0294 for subtypes C and B, respectively. Subtype C isolates also tend to acquire PTAP duplications with a smaller number of accumulated PIR mutations.
Although PTAP duplications can occur in isolates from both treatment-naive and treatment-experienced patients, and are present in different subtypes of HIV-1, the frequency of these duplications is dramatically increased in isolates under HAART selective pressure. This is the first study to demonstrate an association between PTAP duplications and treatment failure in HIV-1 subtypes B and C. These results raise a variety of questions concerning the mechanism by which the duplications are generated. Duplications can be inserted into the HIV-1 genome by RT; whether RT from different subtypes behaves differently in this respect remains to be investigated. During treatment failure, subtype C isolates typically do not accumulate PIR mutations in PR as do subtype B isolates. It has thus been speculated that resistance to PR inhibitors in subtype C strains could arise via mutations in the PR substrate (Gag and Gag-Pol), both at PR cleavage sites or in conserved regions outside cleavage sites. Some studies demonstrated that susceptibility values obtained from PR inhibitors vary when compared to chimera virus carrying only protease or Gag + protease from the same isolate. 13 Since we find a high number of PTAP duplications in subtype C, and this domain is responsible for an important secondary structure conferred by prolines, and its position is close to a cleavage site, we suggest that these duplications may play a role in subtype C virus susceptibility to PR inhibitors. Moreover, subtype C isolates lack the Alix-binding YPXnL motif, which has been shown to play a role in the virus budding process. 5 As the PTAP domain also participates in virus release through interaction with ESCRT machinery, PTAP duplications could be a mechanism to compensate for the natural deletion of the Tyr in position 36 of p6Gag in the YPXnL motif. The lack of this critical Tyr could also contribute to the selection of frequent PTAP duplications in subtype C vs. subtype B and F1 isolates. However, the efficiency of subtype C virus binding to Alix remains to be tested. Although those domains are important to the viral budding process, they are not essential, as the interaction of PTAP and Tsg101 has been shown to be the dominant late domain interaction for promoting HIV-1 release. 2 –4 Duplications within the PTAP motif in p6Gag lead to the insertion of a Ser-Pro-Thr sequence in the overlapping pol open reading frame. This Ser-Pro-Thr sequence was described by Cao et al. as the first cytotoxic T-lymphocyte (CTL) escape insertion mutation. 14 Escape from the host CTL response could thus provide the driving force for PTAP duplications without necessarily involving any specific HAART selective pressure, but in this case the frequency of PTAP duplications would increase over time, which is not observed. Also, the results presented herein clearly show an increase in the PTAP duplication frequencies in isolates from individuals failing ARV drug therapy when compared to isolates from ARV drug-naive individuals, for all clades analyzed. This supports a possible HAART selective pressure acting in favor of these duplications.
The role of PTAP duplications in isolates carrying PIR mutations still has to be investigated. Whether these duplications confer viral advantages in different aspects of HIV-1 replication is of extreme importance. HIV-1 can accumulate these PTAP duplications to increase viral fitness. The high frequency of these insertions in a clinical isolate cohort may indicate the necessity of PR/Gag coevolution for the establishment of a viable productive viral population carrying PIR mutations. The importance of these insertions to virus assembly and viral loads remains to be addressed.
GenBank Accession Numbers
GenBank accession numbers for the p6Gag sequences obtained in this study are HQ315902–HQ316114.
Footnotes
Acknowledgments
We thank Eric Freed for a critical review of the manuscript. This work was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ). A.N.M. is supported by a FAPERJ nota 10 scholarship.
Author Disclosure Statement
No competing financial interests exist.