
Abstract
Highly active antiretroviral therapy (HAART) is associated with fat redistribution and metabolic disorders. The present study was undertaken to evaluate the association between peroxisome proliferator activated receptor (PPAR)α and PPARγ polymorphisms, two genes involved in lipid metabolism and adipocyte differentiation, and elements of the metabolic syndrome, lipodystrophy, or carbohydrate metabolism abnormalities in patients receiving HAART. The frequency distribution of rare alleles for PPARα (L162V) and PPARγ (P12A and H449H) was compared using the chi square test in 363 HIV-1-infected patients classified according to the presence or absence of the metabolic syndrome after 48 months of follow-up on their first PI-containing regimen. The P12A rare g allele was present in 12% patients with normal glucose metabolism, 11% patients with impaired glucose tolerance or impaired fasting glucose, and 35% patients with diabetes (p=0.014). The rare g allele for L162V was present in 14% of patients free of hypertriglyceridemia and in 7% patients with hypertriglyceridemia (p=0.04). The rare g allele for L162V was found in 15% of patients free of any sign of lipodystrophy and 8% with at least one sign of lipodystrophy (p=0.04) and the rare t allele for H449H was found in 14% of patients free of any sign of lipodystrophy and 23% of patients with at least one sign of lipodystrophy (p=0.05). There was no convincing association between any polymorphism of PPARα and PPARγ and each individual component of the metabolic syndrome, except for the relationship of the P12A polymorphism with diabetes. Confirmatory studies on a larger number of individuals are needed.
Introduction
H
The genes regulated by peroxisome proliferator activated receptor (PPARα) express key proteins involved in extracellular lipid metabolism, fatty acid oxidation, homeostasis, and inflammation. 11 Several polymorphisms of the human PPARA gene have been described. In the Framingham offspring study, the less common g allele of the L162V polymorphism (rs1800206) was associated with significantly higher serum concentrations of total cholesterol, LDL cholesterol, apoB, and apoC-III as compared to the c allele. 11 PPARγ plays an important role in the differentiation of adipocytes and in the regulation of insulin sensitivity. Variation in the PPARG gene may be a risk factor for the development of diabetes and the metabolic syndrome. 12 –14 Four PPARγ isoforms with various distribution in tissues have been identified. 15 –17 Two main variants were identified in the PPARG gene, a silent (C/T) substitution at nucleotide 1431 in the sixth exon (H449H, rs3856806) and a Pro12Ala missense mutation due to a C/G transversion (P12A, rs1801282). Studies on the association between the P12A mutation and diabetes or insulin resistance have led to conflicting results. 13,18 –22 A meta-analysis reported a significant increase in diabetes risk (1.25-fold) associated with the most common P12A allele (allele c coding for Pro), 23 but subsequent studies failed to confirm this association. 24,25 The rare P12A allele (allele g) was also associated with a significantly reduced risk of cardiovascular disease. 26,27 The rare H449H allele (allele t) has been unfavorably associated with obesity 28 and coronary heart disease (CHD) with a low consistency. 28 –31
The present study was undertaken to evaluate the association between PPARα and PPARγ polymorphisms and elements of the metabolic syndrome, lipodystrophy, or carbohydrate metabolism abnormalities in patients receiving HAART in the ANRS CO8 APROCO-COPILOTE Study.
Material and Methods
Study design
The ANRS CO8 APROCO-COPILOTE cohort is a prospective observational study that enrolled 1281 HIV-1-infected adults in 47 hospitals in France starting a protease inhibitor (PI)-containing antiretroviral regimen for the first time in 1997–1999. The APROCO COPILOTE–Metabolic Complications (APROCO-MC) substudy has been described elsewhere. 6 For this analysis, as shown in Fig. 1, these results were obtained in a subgroup of subjects who signed an informed consent for participating in the DNAbank in 2002 and have available data on metabolic syndrome at month 48. We checked that these subjects did not differ from those without DNA analyses for their main baseline characteristics. Signs of peripheral atrophy (fat wasting in the face, upper limbs, and lower limbs, prominent limb veins, and buttock atrophy), signs of fat accumulation (accumulation of fat in the face or breasts, buffalo hump, or increased waist size), and signs of metabolic syndrome according to NCEP-ATPIII criteria 9 and the International Diabetes Federation (IDF) 32,33 were recorded. Since almost 16% of the patients followed in the main APROCO Study had a metabolic syndrome, after 12–20 months of PI-based therapy 10 and because PPARα and PPARγ gene polymorphisms are related to these various metabolic abnormalities in the general population, we studied their relationship with metabolic abnormalities in this group of patients, with available clinical data at month 48.
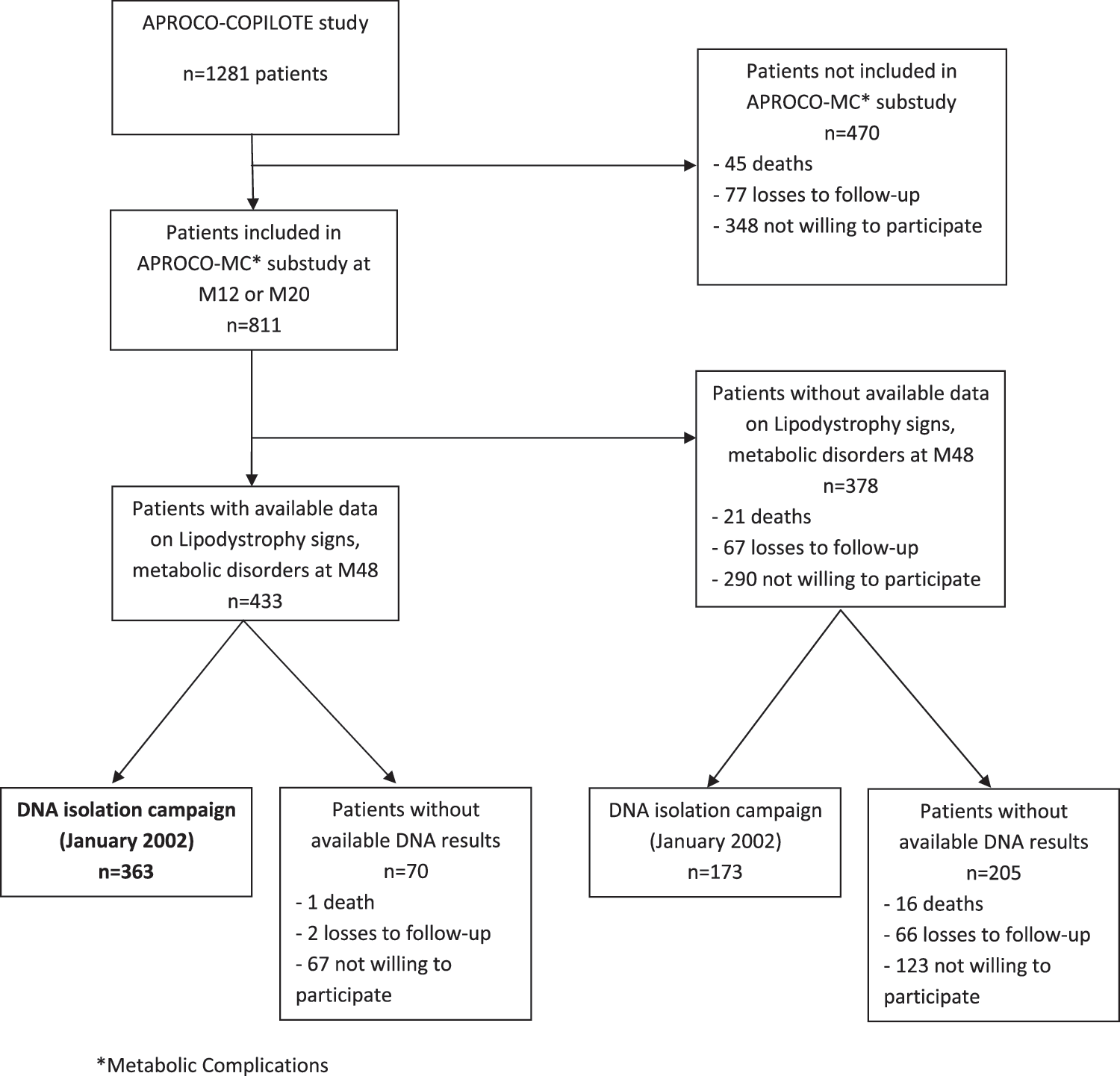
The ANRS CO8 APROCO-COPILOTE study.
Laboratory measurement
Levels of serum glucose, triglycerides, and HDL cholesterol were determined by the routine methods used in the clinical laboratory located in each center. A glucose tolerance test (OGTT) was also performed after an oral load of 75 g of glucose. Glycemia was measured at 120 min time. According to the 1998 World Health Organization (WHO) diagnostic criteria, 34 impaired fasting glucose was defined as a fasting blood glucose level of ≥6.1 and <7.0 mmol/liter (≥110 and <126 mg/dl) with OGTT<7.8 mmol/liter (<140 mg/dl), impaired glucose tolerance was defined as a 2 h blood glucose level of ≥7.8 and <11.1 mmol/liter (≥140 and <200 mg/dl) by the OGTT, the fasting blood glucose being <7.0 mmol/liter (<126 mg/dl), and diabetes mellitus was defined as a fasting serum glucose level of ≥7.0 mmol/liter (≥126 mg/dl) or a 2 h serum glucose concentration of ≥11.1 mmol/liter (≥200 mg/dl) by the OGTT. For the statistical analysis, patients with an impaired fasting glucose and patients with an impaired glucose tolerance were pooled.
Genomic DNA was isolated from peripheral blood leukocytes by standard methods. 35 Genotyping was carried out on an Applied Biosystems 7900 sequence detection system with Taqman probes for allelic discrimination, as described by the supplier. PCR amplification was carried out in the presence of two probes with different reporter dyes attached to their 5′ ends and a fluorescent quencher at the 3′ ends. One probe was complementary to the wild-type DNA strand and the other to the DNA strand with the mutation.
Statistical analysis
The variables of interest were the serological distribution of each polymorphism PPARα and PPARγ, which do not distinguish patients with one or two alleles of interest. Comparisons were made between groups defined by the presence or absence of various clinical signs and were determined by the chi square test. Dichotomous outcomes for lipids, glucose, blood pressure, and waist circumference were chosen according to the definitions of MS. 9,32,33 Models were adjusted for the following baseline characteristics: HIV-1 RNA, CD4 cell count, AIDS status, history of treatment (naive or not), and demographic data (gender, age, and HIV transmission category). Analyses were performed with SAS Software (version 9.1). Due to multiple testing (20 tests/polymorphism), significance with a p-value above 0.0025 should be considered with caution.
Results
Among 1281 patients initially included in the cohort, 811 participated in the APROCO-MC substudy and 363 had available DNA to be included in this analysis (Fig. 1). As the selection reduced the number of patients, we compared baseline characteristics according to whether patients had or did not have data for this analysis (n=363 vs. n=918). Patients included in the analysis did not differ from those excluded in terms of antiretroviral history (antiretroviral naive status: 47% vs. 44%, p=0.31, respectively), AIDS status (23% vs. 19%, p=0.08), median baseline CD4 levels (IQR): 256 (120–418)/mm3 vs. 279 (128–423)/mm3, and HIV RNA: 4.5 log10 cp/ml (3.6–5.2) vs. 4.5 (3.8–5.2), as well as data at M48: median CD4 levels (IQR): 518 (355–690)/mm3 vs. 496 (357–689)/mm3, and HIV RNA (IQR): 1.5 (1.4–2.3) log10 cp/ml vs. 1.7 (1.4–2.7) log10 cp/ml, or median duration on the various antiviral therapies (IQR): 50 (48–71) months vs. 50 (48-67) months. Among the 363 DNA analyzed, the distribution of the various genotypes was as follows: 1% were homozygous for the rare alleles L162V and P12A, 2% for H449H, 88% were homozygous for the wild allele L162V, 84% for P12A, and 80% for H449H; 11% were heterozygous for L162V, 16% for P12A, and 18% for H449H. Multivariate analysis did not evidence significant associations of the tested characteristics with the studied polymorphisms (Table 1).
Table 2 presents the percent distribution, among the 363 DNA analyzed, of the rare alleles according to the presence or the absence of various signs of the metabolic syndrome. As shown in Table 2, there was no difference in the serological distribution between patients with the metabolic syndrome and patients who did not have the syndrome. However, there was a difference in the frequency of the rare allele of PPARα L162V when comparing patients with high triglycerides (≥150 mg/dl) with subjects with normal triglycerides (<150 mg/dl): 7% vs. 14%, p=0.041.
Results are numbers with rare allele/total number of patients in the category.
WC, high waist circumference (≥103 cm in men or ≥88 cm in women for NCEP and ≥94 cm in men or ≥80 cm in women of European origin); TG, high triglycerides (≥150 mg/dl or 1.70 mmol/liter); HDL, low HDL cholesterol (<40 mg/dl or 1.00 mmol/liter in men or <50 mg/dl or 1.30 mmol/liter in women); HFG, high fasting glucose (≥100 mg/dl or 5.55 mmol/liter); HBP, high blood pressure (≥135/80 mm Hg); MS, metabolic syndrome; ND, not determined.
Number of ND in PPARα L162V: 6 (HBP), 7 (WC NCEP, WC IDF), 10 (MS IDF), 12 (TG), 13 (HDL, HFG, MS NCEP). Number of ND in PPARγ P12A: 8 (HBP), 10 (WC NCEP, WC IDF), 12 (MS IDF), 15 (TG), 16 (HFG), 18 (MS NCEP), 20 (HDL). Number of ND in PPARγ H449H: 12 (WC NCEP, WC IDF), 13 (HBP), 16 (MS IDF), 20 (TG), 21 (HFG), 24 (HDL).
Table 3 presents the percent distribution among the 363 DNA analyzed of the rare alleles according to the presence or the absence of signs of lipodystrophy. As shown in Table 3, the rare allele for PPARγ P12A was present in 12% of patients with normal glucose metabolism, 11% of patients with IGT or IFG, and 35% of patients with diabetes (p=0.014). The rare allele for PPARα L162V was found in 15% of patients free of any sign of lipohypertrophy and 8% patients with at least one sign of lipohypertrophy (p=0.04), while the rare allele for PPARγ H449H was found in 14% of patients free of any sign of lipoatrophy and 23% patients with at least one sign of lipoatrophy (p=0.05).
Normal, fasting glucose level <6.1 mmol/liter. IFG/IGT, impaired fasting glucose [fasting blood glucose level of ≥6.1 and <7.0 mmol/liter (ge;110 and <126 mg/dl) and <7.8 mmol/liter (<140 mg/dl) by OGTT]/impaired glucose tolerance [2 h blood glucose level of ≥7.8 and <11.1 mmol/liter (≥140 and <200 mg/dl) by the OGTT]. Diabetes, fasting serum glucose level of ≥7.0 mmol/liter (≥126 mg/dl) or a 2 h serum glucose concentration of ≥11.1 mmol/liter (≥200 mg/dl) by the OGTT.
Not done in PPARα L162V: glucose metabolism (N=15, 13%), lipoatrophy (N=2, 8%), lipohypertrophy (N=2, 8%). Not done in PPARγ P12A: glucose metabolism (N=31, 27%), lipoatrophy (N=6, 25%), lipohypertrophy (N=6, 25%). Not done in PPARγ H449H: glucose metabolism (N=32, 28%), lipoatrophy (N=9, 38%), lipohypertrophy (N=9, 38%).
Discussion
This study focuses on the association of three polymorphisms of PPARα and PPARγ with the metabolic disorders present in treated HIV-infected patients. Our results do not exclude any other association of some polymorphisms important for lipoprotein and carbohydrate metabolism, such as lipids carriers, enzymes, inflammation factors, or oxidative stress as already suggested. The association of the polymorphisms studied with the metabolic syndrome was studied with the syndrome defined according to the most popular definitions. 9,32,33 Since these definitions are still under discussion, we also studied these relationships with individual components of the syndrome. To the same extent, the diagnosis of lipoatrophy and lipohypertrophy was done on clinical observations and is therefore subject to some subjectivity. This represents clear limitations in the interpretation of our results.
The frequency distribution of the various polymorphisms is in agreement with those observed elsewhere in general populations and subjects suffering from various metabolic disorders. 11,13,20,23,31,36 –40 No difference in the frequency distribution of the three rare alleles studied was observed in the two groups of patients defined by the presence or the absence of the metabolic syndrome. Some studies report a relationship between the presence of the P12A polymorphism and insulin resistance, with an apparent protection of the rare allele toward the development of adiposity or insulin resistance. 36,37 However, in a large French population study, individual polymorphisms failed to demonstrate any relationship with the metabolic syndrome while the influence of a haplotype combining four PPARγ polymorphisms, including P12A and H449H, was suggested. 38
Another PPAR polymorphism was also studied and no clear association of this polymorphism with lipodystrophy was observed because of a lack of power. For the PPARα polymorphism, we may point out that when we considered the individual signs of the metabolic syndrome, there was a trend toward a different frequency distribution of the PPARα gene L162V rare allele in the presence or absence of high fasting triglycerides. This PPARα gene polymorphism has been shown in the Framingham population to be associated with higher concentrations of LDL cholesterol, apolipoprotein B, and apolipoprotein C-III. 11
Here we show that the rare allele seems to be less frequent in the case of triglycerides above 150 mg/dl. It was previously shown, in young healthy volunteers, that the rare allele was associated with an increase in fasting triglyceride concentrations. 39 However, there is obviously an interaction between the PPARα gene polymorphism and diet. In an article taking into consideration polyunsaturated fatty acid (PUFA) intake, it was shown that the effect of the rare allele was different in low PUFA consumers and high PUFA consumers. 40 Moreover, some functional studies demonstrated that PPAR α agonists (omega 3 fatty acids) have different effects in transcriptional activation by the allelic variants of PPARα. 41 –48 It may be assumed that the lifestyle of our population may interact with the gene polymorphism to explain these unexpected results. In addition, we observed that the rare allele for PPARα L162V seems to be less frequent in the presence of lipohypertrophy than in the absence of any sign of lipohypertrophy. If this is confirmed, this would suggest that our HIV-infected population behaves differently than noninfected subjects. However, no data on diet in our population are available. This is clearly a limitation of our study.
The rare allele frequency distribution of P12A was slightly different between the three groups of patients defined by their glucose tolerance (p=0.014) with a higher frequency of this rare allele in diabetics (35%) than in normal subjects (12%) and IFG/IGT (11%). In a previous Japanese-American study, the rare allele was found in 9.3% of subjects with normal glucose tolerance vs. only 2.2% in patients with diabetes. 13 This association was reproduced in two large population studies from Finland and Japan. 41 –44 However, several other studies failed to confirm this association. 18 –21,43
The most common finding is that the P12A polymorphism is associated with a greater insulin sensitivity. 13 In addition, subjects with obesity experienced a more pronounced difference in insulin sensitivity in normal 18,22 and in HIV-infected subjects. 45 In an Asian population, 31 the P12A polymorphism was associated with fasting glucose concentrations, whereas the H449H polymorphism was related to the risk of diabetes, while the effect of P12A on fasting glucose was modified by diabetes status. Obviously our findings of an increased frequency of the rare P12A allele in the presence of diabetes do not fit with what was observed in noninfected populations. However, in the HIV-infected subjects, the effect of the P12A polymorphism could differ between insulin sensitivity and diabetes. On the other hand, expression of the mutant protein in vitro has produced results that do not fit with most of the epidemiological studies. As a matter of fact, Masugi et al. 49 have shown that the ability of the P12A mutant protein to mediate adipogenesis in response to thiazolidinedione is impaired, suggesting a deleterious influence of this variant on muscle sensitivity. In addition, our observation of an association between the presence of the rare H449H allele and the presence of lipoatrophy would indicate that our HIV-infected population behaves differently than noninfected subjects. However, we might speculate that infection, treatment, and/or lifestyle interact with the PPARs gene, modulating its effects.
One major limitation of this study was the potential lack of power for the analysis of rare alleles, leading us to cautiously interpret these results in the context of multiple testings (i.e., 20 tests per polymorphism) when p values are above 0.0025 (0.05/20 according to the Bonferroni rule).
In conclusion, this study failed to demonstrate clear associations between studied PPARs polymorphisms and the presence or absence of components of the metabolic syndrome. A tendency was observed for some of these polymorphisms, which could be the basis for further studies with larger numbers of subjects.
Footnotes
Author Disclosure Statement
In the past 3 years, G Chêne has received consulting fees (Scientific Committee) from Roche. She has had scientific responsibilities in projects for which her institution or a non-profit society received specific grant support from Gilead, Tibotec, Boehringer Ingelheim, GlaxoSmithKline, Roche, Pfizer, MSD, Bristol-Myers Squibb, Janssen, and ViiV Healthcare. All other authors report no competing financial interests exist.