
Abstract
The incidence of herpes zoster remains high in HIV-infected patients despite the use of combined antiretroviral therapy (cART). We wished to assess varicella-zoster virus (VZV)-specific cell-mediated immune (CMI) responses in HIV-infected adults on cART. VZV-specific CMI responses were assessed using lymphocyte proliferative responses, cytokine production (IL-2, TNF-α, and IFN-γ), and interferon-γ ELISPOT assays in 103 HIV-infected adults and 30 healthy controls. HIV-infected patients were analyzed according to their current and nadir CD4 cell count and their use of cART. A multivariate analysis was performed to identify factors associated with VZV-specific CMI responses. HIV-infected patients had lower VZV-specific CMI responses than healthy controls. Patients with a CD4 T cell count <100/μL had almost no detectable responses whereas those with a current CD4 T cell count >300/μL and suppressed viral replication on cART had responses similar to those of healthy controls. In multivariate analysis, factors significantly associated with VZV-specific CMI responses were the absence of a previous AIDS-defining event and higher CD4 cell counts, in particular central and effector memory CD4 T cell counts. HIV-infected patients with a history of AIDS or low CD4 cell counts have impaired VZV-specific CMI responses, and remain at risk for herpes zoster.
Introduction
T
Herpes zoster is due to the reactivation of varicella-zoster virus (VZV) in patients with impaired VZV-specific cell-mediated immune (CMI) responses, as occurs with age or immune suppression. 12 Indeed, improvement of VZV-specific CMI following vaccination in elderly and bone marrow transplant recipients has been associated with a decrease in the incidence and severity of herpes zoster. 13,14
Limited data regarding VZV-specific CMI responses and their restoration under cART are available among HIV-infected adults. We therefore performed a cross-sectional study among HIV-infected adults with different levels of immune suppression, with or without cART, to assess their VZV-specific CMI responses, and to identify factors associated with the restoration of such responses. VZV-specific immune responses were assessed by lymphocyte proliferative assays, quantification of TH-1 cytokine production, and gamma-interferon (IFN-γ) ELISPOT assays in the presence of VZV antigens.
Materials and Methods
This cross-sectional study was performed between January 2005 and July 2006 in the Department of Infectious Diseases of Saint-Louis Hospital in Paris, France.
We consecutively enrolled HIV-infected adults at different levels of CD4 cell counts, with or without cART. All patients were screened for anti-VZV IgG antibodies and we enrolled only patients with detectable VZV antibodies. We defined four groups of HIV-infected patients: group 1: patients with a current CD4 T cell count <100 cells/μL with or without cART; group 2: patients with a current CD4 T cell count >300 cells/μL on cART but with a nadir CD4 cell count <100 cells/μL; group 3: patients with a current and nadir CD4 T cell count >300 cells/μL on cART; and group 4: patients with a current and nadir CD4 T cell count >300 cells/μL but naive to cART. We wished to enroll 15 to 30 patients per group.
Thirty HIV-negative healthcare workers with positive anti-VZV IgG antibodies were enrolled as healthy controls (group 5).
Ethics review
The study protocol was approved by the ethics committee of Paris Saint-Louis and all participants gave written informed consent.
Plasma viral RNA assay
Plasma HIV-1 RNA levels were measured by an ultrasensitive PCR assay (Ultrasensitive Amplicor HIV-1 1.5 Monitor, Roche) with a lower limit of quantification of 50 copies/mL.
VZV serology
Quantitative detection of VZV IgG antibodies was performed using the enzyme-linked immunosorbent assay Captia VZV IgG (Trinity Biotech, Jamestown, NY), according to the manufacturer's recommendations.
Flow cytometry analysis
Lymphocyte immunophenotyping was performed on fresh whole blood EDTA samples by direct three- or four-color immunofluorescence, using appropriate monoclonal antibodies (Becton Dickinson, San Jose, CA and Coulter-Beckman-Immunotech, Villepinte, France) as previously reported. 15 Ten thousand gated lymphocytes were analyzed with a Facscalibur analyzer (Becton Dickinson, San Jose, CA). Lymphocyte gate purity was ≥98%. Percentages and absolute counts of competent (CD28+), naive (CD45RA+CCR7+), central memory (CD45RA−CCR7+), effector memory (CD45RA−CCR7−), terminally differentiated (CD45RA+CCR7−) CD4 and CD8 T cells subsets, naive (CD19+CD27−), and memory (CD19+CD27+) B cells, and (CD3−CD56+) NK cells were assessed using standardized procedures with the following antibodies: CD45-PerCP, CD3-FITC, CD4-FITC, -APC, or -PerCP, CD8-APC or -PE, CD28-PE, CD45RO-PE, DR-PE, CD56-PE, CD19-FITC (Becton Dickinson, San Jose, CA), CD8-PerCP-Cy5.5, CD45RA-APC, CD27 PE (Pharmingen), CCR7-PE (R&D), and CD5-PE-Cy5 (Immunotech Coulter Beckman, Villepinte, France).
Lymphocyte proliferative responses (LPRs)
Peripheral blood mononuclear cells (PBMCs) isolated from sodium heparinate peripheral blood samples by gradient density centrifugation were cultured in triplicate wells for 7 days in RPMI medium supplemented with 10% human AB serum (Biowest, Abcys, Paris) in the presence of PHA (0.5 μg/mL; Murex, UK) and interleukin (IL)-2 (10 IU/mL; Boehringer, France) as a positive control, VZV antigens (VZV ROD infected cell extracts), and uninfected cell extracts as negative control (1/200, Tebu-bio, Le Perray en Yvelines, France). LPRs were quantified by incorporation of tritiated labeled thymidine (Specific activity 185 GBq/mmol, 1 μCi/well) (Amersham, Buckinghamshire, UK) added during the last 18 h of culture.
Results were expressed as mean counts per minute (cpm) per 105 cells and as stimulation index (SI) calculated for each sample as the ratio of cpm in VZV-stimulated versus mock-stimulated cultures. A stimulation index equal or greater than 3 associated with a cpm count ≥3000 defined a positive antigen-specific response. 6
IFN-γ ELISPOT assays
Fresh PBMCs were stimulated with VZV antigen 1/50 (Tebu-bio, Le Perray en Yvelines, France), control antigen 1/50 as negative control and PHA 0.5 μg/mL (Murex) as positive control. PBMCs (2×105 cells) were added in triplicate to a 96-well ELISPOT plate (Millipore) coated with antihuman IFN-γ antibody (Diaclone) for 40 h at 37°C. Antigen-specific spot forming cell (SFC) frequencies were measured with an automated microscope (Zeiss, Munich, Germany) and expressed per 106 PBMCs. Responses were considered positive if superior or equal to 25 SFC/106 PBMCs detected after subtracting the background. 16
Cytokine production
PBMCs were also cultured for 48 h with either VZV antigens, a combination of anti-CD3 (5 μg/mL) and anti-CD28 (0.5 μg/mL) monoclonal antibodies (Coulter Beckman) as positive control or medium alone as negative control. Supernatants were collected and stored at −80°C until quantification of TH-1 [IL-2, IFN-γ, tumor necrosis factor (TNF)-α] and TH-2 (IL-4, IL-5, IL-10) cytokine production using a CBA kit (Becton Dickinson) combining a “sandwich” immunoassay and flow cytometry. 17 Standard curves ranging from 0 to 5000 pg/mL were established for IL-2, IFN-γ, TNF-α, IL-4, IL-5, and IL-10, respectively. Cytokine production was expressed as delta (Δ) pg/mL defined as Δ=(production in stimulated wells) – (production in unstimulated wells). Patients with a Δ>10 pg/mL of IFN-γ were considered as responders, as were patients with Δ>5 pg/mL for the other cytokines.
Statistical analysis
Results are expressed as the median (interquartile range) for continuous variables and N (%) for categorical variables. The effects of continuous covariates were studied using the Student's t-test or the Wilcoxon test when appropriate. Fisher's exact tests were used for categorical variables. All p-values were adjusted for multiple comparisons using the Hochberg method. 18
Predictors of positive ELISPOT and LPR responses were assessed by logistic regression, restricted to HIV-infected patients (groups 1 to 4). For each outcome (response to ELISPOT and LPR), factors achieving a p-value <0.05 in the univariate analysis were included in the multivariate analysis. A backward stepwise variable selection procedure was used to remove factors with p-value >0.05. Analyses were carried out using the R 2.4.0 statistical package [R Development Core Team; R Foundation for Statistical Computing, Vienna, Austria (
Results
Patients characteristics
Patients characteristics are shown in Table 1.
p-values are reported for comparisons between all HIV-infected patients (group 1+2+3+4) and healthy controls (group 5).
cART, combined antiretroviral therapy; CDC, Centers for Diseases Control and Prevention; IQR, interquartile range; NR, not relevant.
Between January 2005 and July 2006, 103 HIV-infected adult patients (groups G1 to G4) and 30 HIV-uninfected controls (group G5), all seropositive for VZV, were enrolled in the study.
HIV-infected patients were on average 11 years older than controls, and more frequently males (p<10−4 for both comparisons). Also, HIV-infected patients had lower CD4 cell counts and anti-VZV IgG titers than controls, but had a more frequent history of zoster (p=10−4, p=0.02, and p<10−4, respectively).
By definition, patients in group 1 had the lowest CD4 cell counts, but 86% were receiving cART although plasma HIV RNA levels were below 50 cp/mL in only 29% of them. Patients in the other groups had all CD4 cell counts >300 cells/μL, but only patients in groups 2 and 3 were receiving cART with full suppression of HIV replication in plasma. The median duration of cART was 3, 27, and 18 months in groups 1, 2, and 3, respectively. As expected according to their low nadir or current CD4 cell count, patients in groups 1 and 2 experienced AIDS-defining events and zoster more frequently than patients in groups 3 and 4 (p<10−4 and p=0.18, respectively).
Phenotypic characteristics of T and B lymphocytes
CD4 and CD8 T cell and B cell subsets in the different groups of patients and controls are shown in Fig. 1.

(
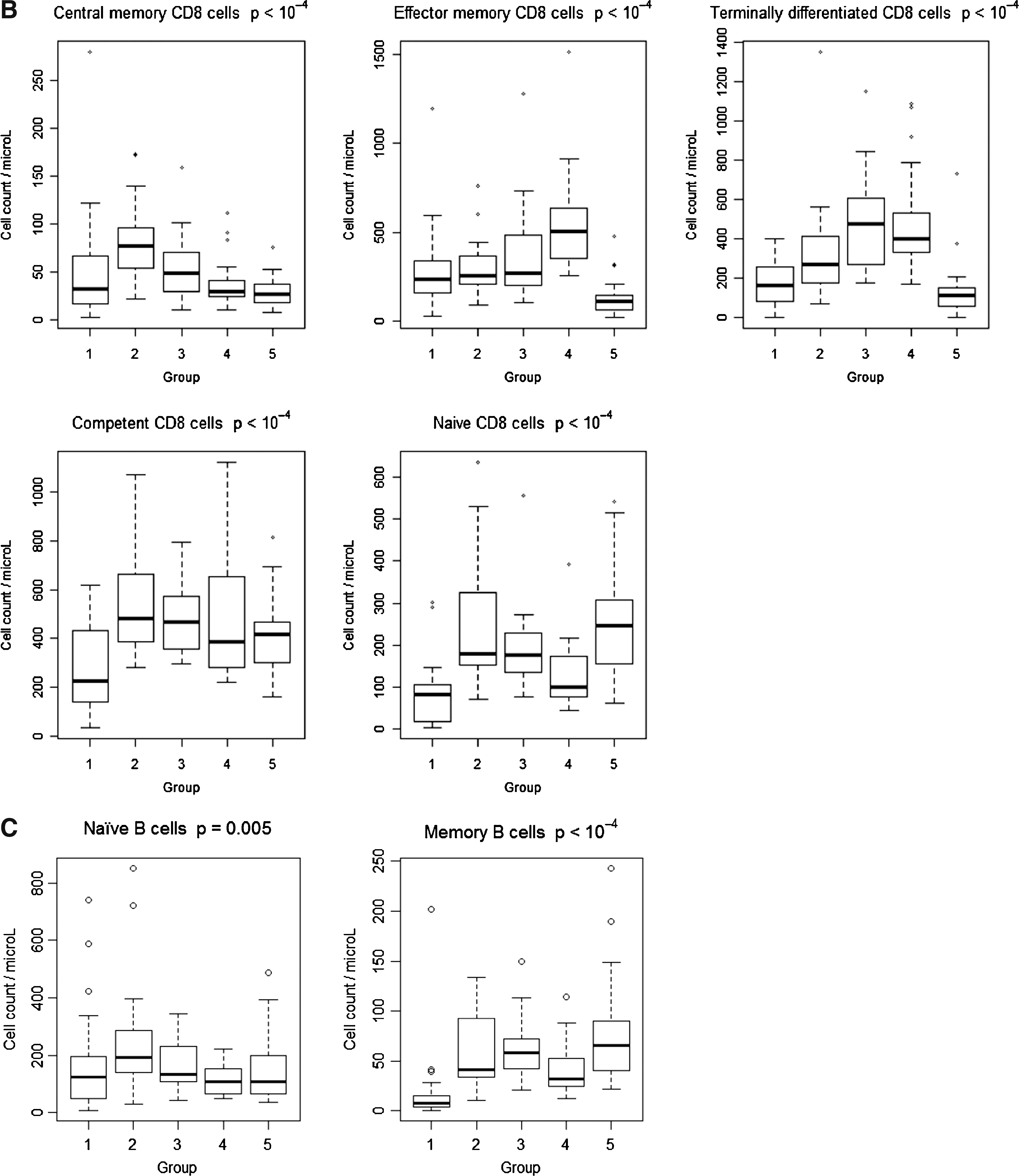
Lower CD4 T cells and T cell subsets were seen in all HIV-infected patient groups as compared to controls (p<10−4) except for terminally differentiated CD4 T cells. Naive CD8 T cells were also lower in HIV-infected patients as compared to controls, but with a concomitant increase of total, effector memory and terminally differentiated subpopulations (p<10−4).
Median absolute counts of B cells were in the normal range in each group, with a decrease of memory subset observed in group 1 (p<10−4). NK cell counts were in the normal range in each group but with a significant decrease in HIV-infected patients as compared to controls (p=0.002; data not shown).
LPRs to VZV antigens
As shown in Table 2, the proportion of patients with a positive LPR and the magnitude of the response (SI) were significantly lower in patients with HIV infection as compared to controls (p=0.0006 and p=0.005, respectively). Interestingly, among HIV-infected patients, the proportion of patients with positive LPRs in groups 3 and 4 (CD4 T cells >300/μL with or without cART) was not statistically different from that of controls (p=1). Only patients with low nadir or current CD4 T cell counts had impaired LPRs.
p-values are reported for comparisons between all HIV-infected patients (group 1+2+3+4) and group 5.
SI, stimulation index, calculated for each sample as the ratio of cpm in VZV-stimulated versus mock-stimulated cultures; cpm, counts per minute; SFC, spot-forming cells; cART, combined antiretroviral therapy.
Gamma-Interferon ELISPOT responses to VZV antigens
The proportion of patients with production of IFN-γ in the presence of VZV antigens detected by ELISPOT assays was significantly lower in HIV-infected patients than in controls (Table 2). Indeed, only 4% of patients in group 1, 30% in group 2, 37% in group 3, and 19% in group 4 had a positive IFN-γ ELISPOT responses compared to 67% of controls. Only patients in group 3 had responses not significantly different from those of controls (p=0.3).
Cytokines production after VZV antigenic stimulation
Production of TH-1 cytokines (IFN-γ, IL-2, TNF-α) is shown in Fig. 2. IFN-γ and IL-2 were produced by 90% of controls in response to VZV stimulation, whereas their production was significantly lower in HIV-infected patients (p=0.005 and p=0.0006 for IFN-γ and IL-2, respectively). HIV-infected patients from groups 1, 2, and 4 had low response rates, and only patients from group 3 had responses that were not significantly lower than controls (p=0.4 and p=0.2, respectively). Production of TNF-α was limited in all groups, almost no production of IL-4 and IL-5 was detected, and IL-10 was produced by 37% of controls but less than 5% of HIV-infected patients overall (data not shown).
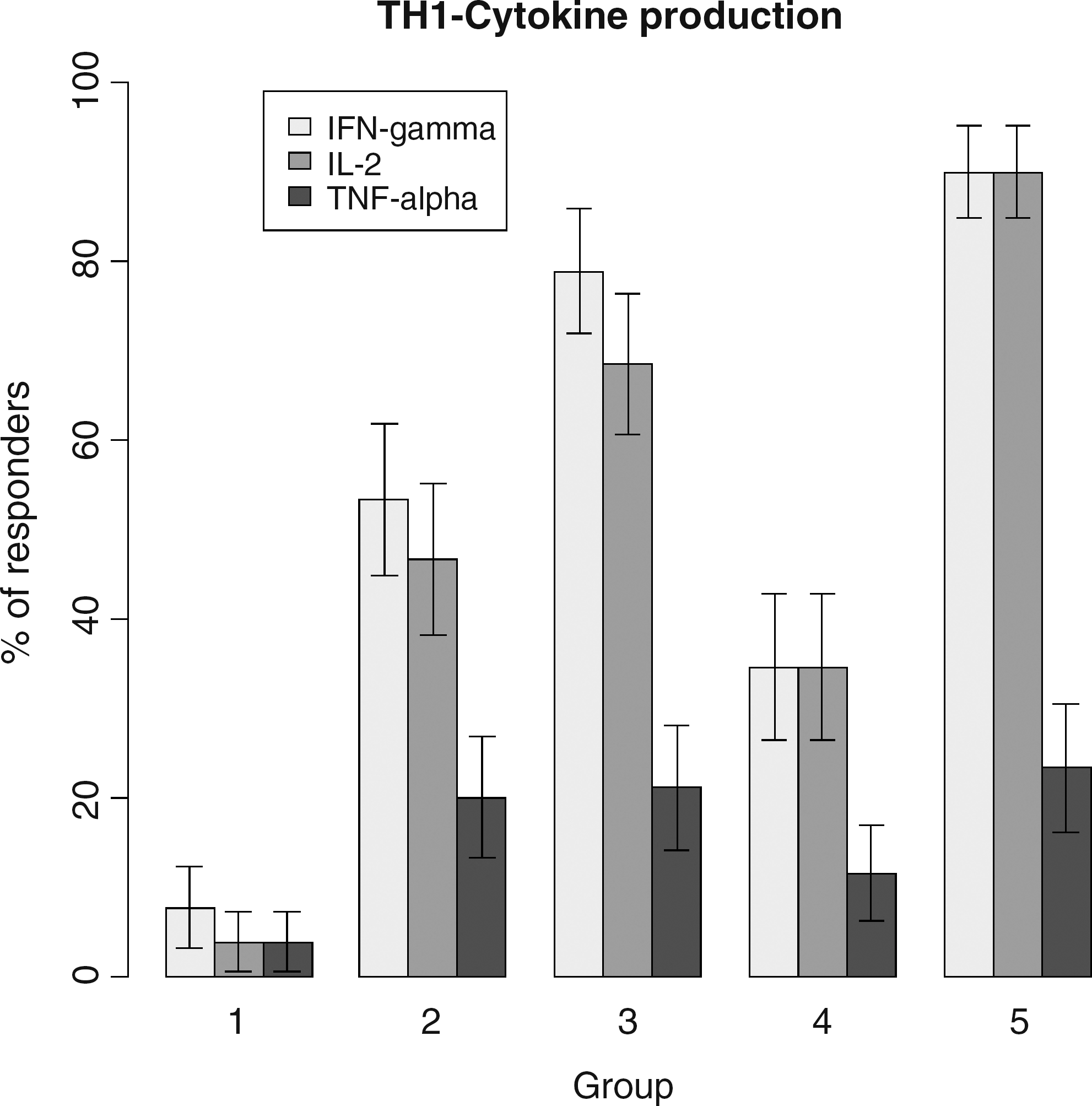
Proportions of HIV-infected patients (groups 1 to 4) and non-HIV-infected controls with gamma-interferon (IFN-γ), interleukin-2 (IL-2), and tumor necrosis factor (TNF)-α cytokine production in peripheral blood mononuclear cell culture supernatants following varicella-zoster virus (VZV) antigenic stimulation.
Predictors of VZV-specific T cell proliferation and IFN-γ ELISPOT responses
Univariate and multivariate analyses were performed to determine predictors of a positive LPR and IFN-γ ELISPOT response in HIV-infected patients (group G1 to G4, n=103).
Fifty-three patients were responders and 50 non responders using the LPR assay. In the univariate analysis (data not shown), we found a significant association between a positive LPR and the following parameters: age (p=0.03), male sex (p<0.002), patients receiving cART (p=0.02), no prior history of an AIDS-defining event (p<0.01), higher CD4 T cell counts (p<10−4), CD8 T cell counts (p<10−4), and B cell counts (p=0.02). In the multivariate analysis (Table 3a), only the absence of a prior AIDS-defining event (OR 9.8, p<10−4) and CD4 T cells (OR: 1.9 per 100 cells/μL increase, p<10−4) were associated with a positive LPR. When CD4 T cell subsets were incorporated in the model instead of total CD4 T cells (Table 3b), the absence of a prior AIDS-defining event remained significantly associated with a positive LPR (OR 20.0, p<10−4), as well as central memory CD4 T cells (OR 1.2 per 10 cell/μL increase, p<0.008) and effector memory CD4 T cells (OR 1.2 per 10 cell/μL increase, p=0.02).
CDC, Centers for Disease Control and Prevention.
CD4 T cell subsets were included in the model instead of total CD4 T cell counts.
CDC, Centers for Disease Control and Prevention.
For IFN-γ ELISPOT assays, 23 patients were responders (22.3%) and 80 non responders (77.7%). In the univariate analysis, we found a significant association between a positive ELISPOT response and the following parameters: patients with a plasma HIV RNA <50 copies/mL (p=0.03), no prior history of an AIDS-defining event (p=0.003), and higher CD4 T cell counts (p<10−4) and B cell counts (p=0.003). In the multivariate model (Table 3 ), only the absence of prior AIDS-defining event (OR 6.5, p=0.007) and CD4 T cells (OR: 1.3 per 100 cells/μL increase, p=0.008) were associated with a positive LPR. When CD4 T cell subsets were incorporated in the model instead of total CD4 T cells, the absence of a prior AIDS-defining event remained significantly associated with a positive ELISPOT response (OR 5.4, p=0.01), as well as central memory CD4 T cell counts (OR 1.1 per 10 cell/μL increase, p=0.005).
Discussion
In this study, using three different assays (LPR assays, IFN-γ ELISPOTs, and cytokine production in PBMC culture supernatants), we have observed impaired VZV-specific CMI responses among HIV-infected patients as compared to healthy controls. These data confirm and extend previous reports in children with HIV infection in whom VZV-specific CMI were low before the initiation of cART and did not significantly improve over the course of more than 3 years on cART. 19 Interestingly, none of the five children who developed herpes zoster had VZV CMI in that study. 19 Similarly, HIV-infected adults of less than 50 years were also found to have significantly lower VZV CMI than age-matched healthy controls in another study. 14
When assessing the predictors of VZV-specific CMI in our study, we found that both the level of CD4 T cells and a history of an AIDS-defining event were strongly associated with these responses. Also, when analyzing CD4 T cell subsets, central memory CD4 T cells, and to a lesser extent effector memory CD4 T cells, were significantly associated with VZV-specific CMI.
These findings were not unexpected since specific memory T cells, mainly TH-1 CD4 T cells, are crucial to control VZV reactivations. 12,20 Also, low CD4 T cell counts and a history of AIDS have been identified as strong predictors of herpes zoster in HIV-infected patients. 8 –11 Finally, patients with the lowest nadir CD4 counts, most of whom also experienced an AIDS-defining event in our study (patients in groups 1 and 2), had a frequent history of herpes zoster (43%) (Table 1). Indeed, in patients with low CD4 counts (<100 cells/μL) such as those from group 1 in our study, less than 5% had detectable VZV-specific CMI responses despite the use of cART in 86% of them (Table 2 and Fig. 2), and these patients therefore remain potentially at high risk of herpes zoster.
On the other hand, patients from group 4 who were antiretroviral naive with more than 300 CD4 T cells/μL (median CD4 cell count of 425 cells/μL) exhibited VZV-specific LPR similar to controls. However, IFN-γ ELISPOT responses and TH-1 cytokines production were significantly impaired as compared to controls (Table 2 and Fig. 2). These patients had persistent HIV replication that could explain these impaired responses, but HIV replication itself was not found to be associated with impaired VZV-specific CMI in our study, contrary to a previous report. 19 The confounding effect of cART on CD4 cell counts and HIV replication might explain these differences. Indeed, another study found no association between high HIV viral load and the incidence of herpes zoster, once the use of cART for more than 90 days was included in a multivariate model. 8
Of interest, central memory CD4 T cells in these untreated HIV-infected patients were lower than in patients with similar CD4 T cell counts receiving cART (groups 2 and 3), and could therefore explain impaired VZV-specific CMI (Fig. 1). Interestingly, patients without a history of AIDS, with high CD4 counts on cART (median 535 cells/μL), were found to have the best VZV-specific CMI, not significantly different, although lower, than those of healthy controls. Indeed, 90% of these patients from group 3 had positive LPR (Table 2), and more than 70% produced TH-1 cytokines in response to VZV antigens (Fig. 2). IFN-γ ELISPOT responses, although not statistically different from those of controls, were also lower (37% vs. 67%, respectively, p=0.3). These differences might explain why HIV-infected patients remain at higher risk of herpes zoster even after several months on cART with high CD4 cell counts and well suppressed HIV replication.
Due to the cross-sectional design of our study we were unable, however, to correlate these ex vivo measurements of VZV-specific CMI with clinical endpoints, i.e., the occurrence of herpes zoster. This is a clear limitation to our results that cannot be used to identify patients with HIV infection that might be at highest risk of VZV reactivation.
Previous studies were able to demonstrate an increase in VZV-specific CMI following vaccination in elderly patients, the magnitude and duration of which paralleled the clinical effect of the vaccine. 13 Unfortunately, the assays performed to measure VZV-specific CMI also failed to identify a specific level for any immune response that was predictive of protection against herpes zoster. Similar findings were reported in bone marrow transplant recipients immunized with inactivated varicella vaccine. 14,21
One study also showed an increase in VZV antibody following vaccination that also correlated with vaccine efficacy. 13 The assay was based on a quantitative ELISA method to detect antibodies to VZV glycoproteins purified from VZV-infected human fibroblasts. 22 We did not find any association in our study between VZV antibody titers and LPR or IFN-γ ELISPOT responses, although VZV antibody titers were lower in HIV-infected patients as compared to controls, but we did not use the same assay (Table 1).
Our study also had other limitations. It was a relatively small study and differences seen between HIV-infected patients and controls might be also explained, at least in part, by demographic differences: indeed, controls were younger and more frequently females.
Age in particular has been associated with VZV-specific CMI, and older age (above 50 years) is associated with impaired CMI. 23 In immunocompetent individuals with prior varicella VZV-specific CMI peaks in early adulthood around 34 years of age. 24 Although controls were 11 years younger than HIV-infected patients in our study (29.8 vs. 40 years, p<10−4), median ages in both groups were on both sides of this peak and should not have significantly affected the results.
Another potential bias in our study resulted from repeated exposure to VZV antigens in controls who were healthcare workers, and which may have boosted their VZV-specific CMI. 20 A significant proportion of our HIV-infected population also had exposure to VZV antigens since 31% of them reported a previous episode of herpes zoster, yet these two types of reexposure may not have the same booster effect on cell-mediated responses.
Finally, we could not establish VZV-specific CMI as the correlate of protection against VZV reactivation and there is the possibility that other factors such as cytotoxic responses that were not assayed in this study might correlate better.
In conclusion, our study confirmed that HIV-infected patients have low VZV-specific CMI and remain potentially at risk for VZV reactivation and herpes zoster. Although HIV-infected patients with CD4 cell counts >500/μL on cART, no history of AIDS-defining events, and well suppressed HIV replication exhibited VZV-specific CMI responses similar to those of healthy controls, these responses were somewhat lower and it was not possible to identify a threshold above which the risk of VZV reactivation would be negligible. These patients would be, however, good candidates for VZV vaccination and recent studies suggest that VZV vaccination is safe in these patients and is associated with an improvement of VZV-specific CMI. 25,26 Larger studies would be needed to confirm the clinical efficacy of such a vaccine strategy.
Footnotes
Acknowledgments
This study was supported by grants from Glaxo Smith Kline and Sanofi Pasteur MSD laboratories, and from the Centre d'Etudes et de Recherches en Infectiologie. This study has been presented in part at the 15th Conference on Retroviruses and Opportunistic Infections, Boston, February 3–6, 2008, abstract 1015.
Author Disclosure Statement
No competing financial interests exist.