
Abstract
Therapeutic vaccinations using human immunodeficiency virus (HIV) antigens in HIV-infected patients on antiretroviral therapy (ART) have so far been attempted with the purpose of inducing CTL response. However, they can also be useful as a strategy for activation of latent HIV reservoir, which is thought to be mainly comprised of latently infected HIV-specific memory CD4 cells, eventually leading to elimination of the virus. The present study was carried out to explore the ability of different HIV antigens to activate HIV replication as assessed by intracellular P24 detection as well as to induce T cell responses in terms of cytokine expression by flow cytometry after stimulation of PBMCs from HIV-infected patients. HIV antigens were found to be able to activate most of the CD4 T cells harboring proviral DNA. HIV-1 Pol and Env were responsible for induction of higher HIV replication in terms of both magnitude and frequency followed by Gag and Nef. As opposed to this, Pol and Env contributed to fewer numbers of polyfunctional CD8 cells desirable for elimination of HIV-infected cells in comparison to Gag and Nef. Thus, HIV antigens may provide a strategy for the activation of a latent reservoir. It was observed that HIV replication started as early as half an hour after in vitro activation indicating a stringent need for maintaining effective concentrations of antiretroviral drugs to prevent further spread of HIV during this process. HIV-infected cells were found to be responsible for higher IL-10 secretion after activation, which could also serve as one of the reasons for suppressed CD8 responses to Pol and Env as more HIV-infected CD4 cells would be secreting IL-10 in response to these antigens. Since IL-10 blockade helped to improve immune responses in terms of cytokine secretion, it should be considered in settings of therapeutic vaccination to improve CTL responses, which will ultimately limit the persistence of the viral reservoir.
H
The major reason responsible for the inability of HAART to eliminate the virus is its ineffectiveness against latent HIV residing in resting CD4 lymphocytes. Latent HIV persists in these cellular reservoirs and acts as a source for persistent HIV infection. Currently it is thought that the combination of agents that will reactivate latent viruses together with HAART is the desired approach toward possible viral eradication. Activation of latently infected cells results in increased expression of viral proteins on the surface of activated cells making them accessible to host immune surveillance. In addition, an intrinsic turnover of cells is accelerated by shortening their natural half-life or inducing death via activation-induced apoptosis. 9,10 Different approaches used for activation of latent infection so far include nonspecific stimuli such as anti-CD3 antibodies, cytokines [interleukin (IL-2, IL-7)], 11 –13 agents such as histone deacetylase inhibitors (valproic acid), 14,15 protein kinase C activators such as bryostatin, 16 –18 bacterial products such as HIV-1-reactivating protein factor, and lipopolysaccharide. 19,20 But to date, none of the clinical trials conducted with the intention of HIV-1 reactivation has succeeded in reducing latent reservoir significantly 21,22 and hence, newer approaches are being evaluated to determine their ability to activate latently infected cells.
The long lived HIV reservoir is mainly composed of HIV-1-infected memory CD4 cells. 23,24 These memory cells can be activated by exposing them to their cognate antigens, which further leads to activation of HIV harbored by them. The use of antigenic stimuli for activation of reservoir would target only the cells recognizing those antigens and, hence, would avoid the possibility of inducing cytokine storm and the associated adverse effects or prolonged depletion of T cells as seen after using nonspecific stimuli such as IL-2 or anti-CD3 antibodies. 11,12 The use of antigens for activating the latent HIV reservoir may potentially have an additional advantage of stimulating cytotoxic T lymphocyte (CTL) response simultaneously, which would cause lysis of the infected cells with reactivated HIV infection. Also, HIV-infected cells are most likely to consist of HIV-specific CD4 T cells as HIV has been found to preferentially infect HIV-specific CD4 cells. 25 Although it was reported by Douek et al. that HIV-infected HIV-specific CD4+ memory T cells constituted only between 1.3% and 10.4% (mean, 5.1%) of all infected CD4+ memory T cells in blood, it is likely to be a minimal estimate as the HIV specificity was determined in the said report by interferon (IFN)-γ secretion and expression of CD69, which may not represent all HIV-infected and HIV-specific cells as all HIV-infected cells may not produce IFN-γ or may not be fully activated. The reason for the preferential infection of HIV-specific CD4 cells is thought to be their prolonged close proximity to actively replicating HIV in the lymph nodes. 26,27 This finding suggests that HIV antigens can potentially be the most specific antigenic stimulus capable of activation of latently infected memory CD4 cells and, hence, may have important implications in settings of therapeutic vaccinations in which HIV antigens are administered.
The present study was conducted with the objective of identifying HIV antigens capable of reactivation of HIV infection in CD4+ T cells. It was hypothesized that HIV will be reactivated after activation of cells by their cognate antigens thereby leading to increased expression of P24 intracellularly. Simultaneously, the functionality of CD4 and CD8 cells including HIV-infected (P24 positive) cells was assessed by estimating intracellular cytokine expression in response to these antigens, which would contribute to containment of the HIV-infected cells after reactivation. Seventeen antiretroviral treatment (ART)-naive HIV-infected patients (F=8, M=9) were enrolled with a median CD4 T cell count of 692 cells/mm3 (range 379–1157) and plasma viral load ranging from undetectable to 783,000 copies/ml (median:17,781 copies/ml). The study was approved by the Institutional Ethics committee, and all participants gave written informed consent.
An intracellular cytokine assay was performed for functional assessment of the cells and intracellular P24 was detected as a marker of HIV replication after stimulation of peripheral blood mononuclear cells (PBMCs) with HIV as well as non-HIV recall antigens. HIV antigens used were peptide pools spanning the HIV-1 Gag (Cat. no. 3993), Pol (Cat. no.6208), Env (Cat. no. 9499), and Nef (Cat. no. 5189) regions from the NIH AIDS Research and Reference Reagent Program, Division of AIDS, NIAID. Non-HIV recall antigens were also used so as to estimate the relative contribution of antigens from common pathogens, which can lead to increased HIV replication.
These antigens included PPD (Beacon diagnostics) and a pool of peptides from influenza, EBV, and cytomegalovirus (FEC: MabTech). PPD, a complex mixture of proteins, and FEC, a pool of HLA class I-restricted peptides, were included in the assays as PPD has been shown to induce specific CD4 responses in 6-h stimulation protocols and FEC has been shown to induce CD4 responses through cross-presentation in different previous studies. 28 –31 Briefly, PBMCs (1– 2×106) were incubated for 2 h with the antigens (5 μg/ml) at 37°C in 5% CO2.; brefeldin A (5 μg/ml, Sigma-Aldrich) was added for 4–6 h to accumulate intracellular cytokines. Anti-CD107a- PECy5, a degranulation marker, was added to the cells along with the peptides. Cells were washed with phosphate-buffered saline (PBS) containing 0.5% bovine serum albumin (BSA) (wash buffer), and fixed by resuspending the cells in lysing solution (BD Biosciences) for 10 min at room temperature. The cells were then washed once with wash buffer and permeabilized by resuspending in 0.2 ml of permeabilizing solution 2 (BD Biosciences), and incubated for 10 min at room temperature. Cells were subsequently washed with wash buffer and stained with anti-CD3-APCCy7, anti-CD4-PETR, anti-IFN-γ-PECy7, anti-IL-10-PE, anti-IL-2-APC, anti HLA DR-APC, anti CCR7- PECy7 (BD Pharmingen), and anti-P24-FITC antibody (KC57-FITC, Beckman Coulter) for 30 min at room temperature. The cells were then washed twice and resuspended in the fixative (3% formaldehyde). At least 30,000 gated lymphocytes per sample were acquired using a FACSaria (BD Biosciences), and analyzed using FACS Diva software. Unstimulated PBMCs were used as negative control. Responses more than twice the response in negative control cells were considered positive. The results were expressed as net percentage of P24-positive CD4+ T or CD8+ T (identified as non-CD4) cells and calculated by subtracting the percentage of negative control response from the percentage of Ag-specific response.
HIV replication as indicated by P24 expression showed an increase after antigenic stimulation of CD4 cells (Fig. 1a–e). Env (n=14/17; 0.5–5.5%) and Pol (n=12/17; 0.6–5.1%) antigens were found to be responsible for significantly higher P24 expression in terms of magnitude and frequency (p<0.05) than other HIV and non-HIV antigens followed by Gag (n=9/17; 0.5–3.7%) and Nef (n=6/17; 0.4–1.1%) (Fig. 1g). The background P24 expression was observed to be between 0.1 and 1.7% in these patients. Various studies have also shown increased HIV replication in resting CD4 cells after incubation with HIV Env antigens previously. 32 –35 Since increased HIV replication poses a risk of viral rebound and increase in the pool of HIV-infected cells after activation, it would be essential to attempt such an intervention under the cover of an effective or even intensified HAART regimen to limit the further spread of HIV infection in the cells.

CD4 % expressing P24 after stimulation with different HIV antigens. Peripheral blood mononuclear cells (PBMCs) were stimulated with HIV antigens Gag
Only four and three of 17 ART-naive patients showed increased p24 expression after stimulation with FEC (0.7–1.1%) and PPD (0.5–3.9%) antigens, respectively. Hence it is possible that although HIV infects predominantly HIV-specific cells, CD4 memory cells of other specificities may also become infected with HIV. It could be one of the reasons for the transient rise in viral load after other vaccinations attempted in HIV-infected patients. 36 PPD has also been shown to induce increased expression of HIV RNA and production of HIV p24 in HIV-infected monocytes 37 and transient HIV viremia has been observed following PPD skin testing in HIV-infected individuals who are PPD skin test-reactive (Garcia, 1998, unpublished).
Since resting CD4+ T cells from peripheral blood of aviremic HIV-infected individuals treated with HAART harbor predominantly latent provirus, 38,39 virally suppressed HIV-infected patients receiving antiretroviral therapy were also enrolled in the study to determine the activation of latent HIV after antigenic stimulation. Eighteen (F=10, M=8) virally suppressed HIV-infected patients on ART for more than 2 years were enrolled. These patients had a median CD4 T cell count of 635/mm3 (range 451–1026/mm3) and undetectable viral loads. Unstimulated cells showed no background P24 expression in 14/18 patients and 0.1% in only four patients. Increase in P24 expression as observed over the background after stimulating cells with these antigens is shown in Fig. 1h. Pol (n=18/18; 0.1–1.3%), Env (n=18/18; 0.1–0.7%), and Gag (n=17/18; 0.1–0.7%) peptides were responsible for significantly higher P24 expression (p<0.05) as compared to non-HIV antigens followed by Nef (n=12/18; 0.1–1.3%), FEC (n=8/18, 0.1–0.2%), and PPD (n=2/18, 0.1%).
To determine whether the cells showing activation of HIV represented resting cells, the cells were also stained for expression of HLA DR in the experiments as resting CD4+ T cells are recognized as CD4+ HLA DRlow. 40 HLA DR being a late activation marker is unlikely to be expressed on cells after 6–7 h stimulation and, hence, was included for identification of nonactivated resting cells. It was observed that P24 expressing CD4 cells ranging from 37% to 97% (median 90%) were HLA DR negative indicating that activated cells showing HIV replication were mainly resting CD4 cells (Fig. 1f). Although lymph tissues represent the ideal samples for conducting such studies on activation of latent infection, it is not always feasible to obtain samples from lymph tissues from patients. Hence many of the previous studies have used PBMCs as a source of resting CD4 cells for conducting studies on latent HIV. CD4 cells expressing CCR7, a homing marker for secondary lymphoid tissues, represent cells present in lymphatic tissue sites, which constitute the major HIV reservoir. Hence simultaneous expression of CCR7 in P24-positive cells after stimulation with different antigens was also determined and 18.5–86.4% (median 53.8%) of CD4 cells expressing P24 were found to be CCR7 positive, indicating the ability of antigens to induce HIV replication from an HIV reservoir (Fig. 1f).
The time course of P24 expression was also studied to determine time kinetics of HIV replication and expression of antigens after activation in HIV-infected cells. PBMCs were stimulated with antigens and harvested at 0.5, 1, 2, 3, and 6 h and after overnight incubation. It was observed that P24 expression was initiated within half an hour of stimulation and reached a peak in 2–3 h. The percent of CD4 cells expressing P24 plateaued after 2–3 h up to overnight incubation (Fig. 2a). A previous study carried out with PBMCs has also shown intracellular expression of HIV proteins as early as within 2 h after transfection of PBMCs in vitro with HIV followed by activation. 41 These results confirmed that HIV reactivation from proviral latency in PBMCs is a very rapid process and hence there is a need for maintaining effective antiretroviral concentrations to block viral replication completely to prevent further spread of virus to other cells in such attempted reactivation.

The cells expressing P24 at 6 h after stimulation with antigens represented cells infected with HIV in vivo, as a single round of HIV replication takes about 1.5 days. 42 To confirm that the p24 expressing cells are the cells not infected ex vivo, the cells were treated with 100 nM/ml of atazanavir during stimulation to prevent secondary infection cycles. Atazanavir selectively inhibits the HIV type 1 (HIV-1) protease enzyme required for the processing of viral gag and gag-pol precursor polyproteins. The interruption of this latter step in the HIV life cycle prevents the efficient processing of viral structural proteins, thus preventing the formation of an infectious and mature viral particle. However, the precursor protein (p55) formed has been shown to be detectable by KC57 (anti-P24 antibody). 43 Hence while conducting experiments for determining the time course for P24 expression, atazanavir was added in a duplicate set of wells and the cells were harvested at different time points. It was observed that there was no difference in P24 expression at different time points in cells incubated with and without atazanavir, indicating that the P24 expressing cells were not infected de novo (Fig. 2a).
To determine the proportion of CD4 cells infected with HIV that were activated by HIV antigens, the HIV viral burden in PBMCs was determined by estimating proviral DNA copies and comparing this to the total percentage of P24+ CD4 cells after stimulation with HIV antigens. To determine the proviral DNA copies, DNA was extracted from PBMCs using a QIAamp DNA mini kit (QIAGEN, Courtaboeuf, France), according to the manufacturer's instructions. The total DNA in extracts was quantified using readings at 260 nm (Nanodrop, Labtech, Ringmer, UK). The elutes were diluted in distilled water to test 1 μg of total DNA per PCR, which was considered to be equivalent to 150,000 cells. 44 The TaqMan PCR was performed targeting a conserved consensus region in HIV-1 gag. The sequences of the forward and reverse primers and HIV-1 TaqMan probe were 5′-ACCCATGTTTACAGCATT ATCAGAAG-3′, 5′- GCTTGATGTCCCCCTACTGTATTT- 3′, and 5′- AGCCACCCCACAAGATTTAAACACCATGT-3′, respectively. The primers, probe, and standards were validated by an in house method for determining HIV viral load at NARI.
Thermocycling conditions were 2 min at 50°C and 10 min at 95°C, followed by 40 cycles at 95°C for 15 s, and 60°C for 1 min. Amplification and data acquisition were carried out using the 7900HT Fast Real-time PCR system (Applied Biosystems). The log10 of the number of targets initially present was proportional to the cycle threshold (CT) and was determined using the standard curve. The standard curve was linear between 10 and 10,000 copies of input DNA, with an r 2 value of 0.97. Final results were reported as the % CD4 cells harboring proviral DNA copies by using the percentage of CD4+ T lymphocytes in PBMCs as estimated by flow cytometry using the following formula: % CD4 cells harboring proviral DNA copies=100×(No. of proviral DNA copies per 150,000 PBMCs/(150,000×% CD4+ T lymphocytes in PBMCs/100). The percentage of CD4 cells containing proviral DNA was calculated considering that each cell contains only one proviral DNA copy. The percentages of CD4 cells containing proviral DNA were almost similar to the total percent CD4 cells expressing P24 after stimulating them with HIV antigens, indicating that most of the cells containing proviral DNA were activated using HIV antigen (Fig. 2b). The lower values obtained by gag real time PCR may be an underevaluation of the actual proviral DNA load as gag real time PCR has been found to be less sensitive than LTR PCR in previous studies. 45,46 This may be due to greater possibilities of mutations in the gag region as compared to the LTR region.
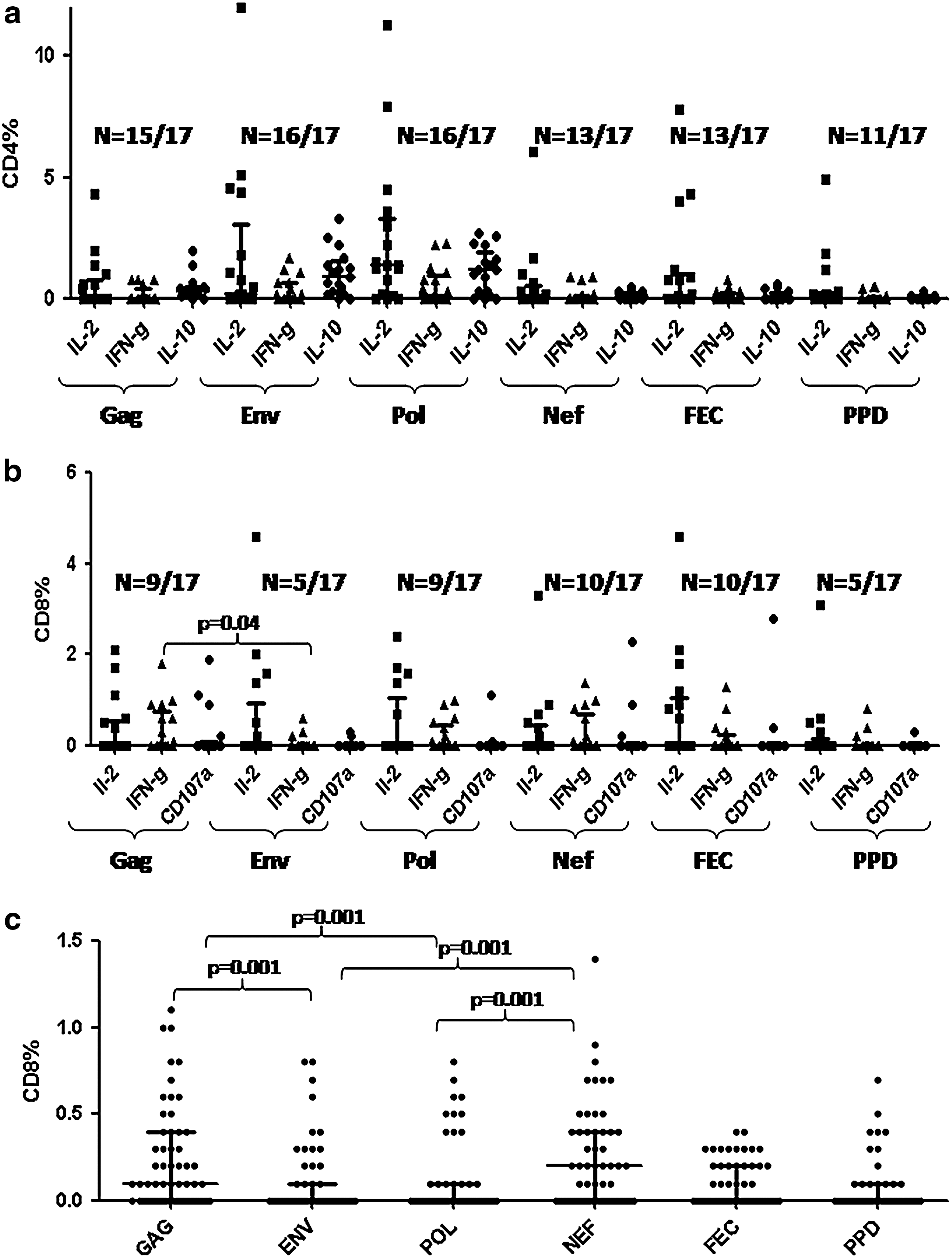
Functionality of CD4 cells responding to these antigens was also studied simultaneously by detecting intracellular cytokines, namely IL-10, IL-2, and IFN-γ (Fig. 3a–c). Although all antigens could induce cytokine secretion in CD4 cells in ART-naive patients (Fig. 4a), the frequency and magnitude of response were more against Pol (n=17/17; IL-2: 0.1–11.3%, IFN-γ: 0.1–2.3%, IL-10: 0.1–2.7%) and Env (n=16/17; IL-2: 0.1–5.1%, IFN-γ: 0.2–1.7%, IL-10: 0.2–3.3%), suggesting more activated CD4 cells, specific to these antigens, acting as targets for HIV infection in vivo. Although a previous study reported Gag to be responsible for maximum CD4 T cell responses followed by Pol, and Env, 47 we have observed more Env and Pol specificity among the CD4 cells responding to antigenic stimulus.
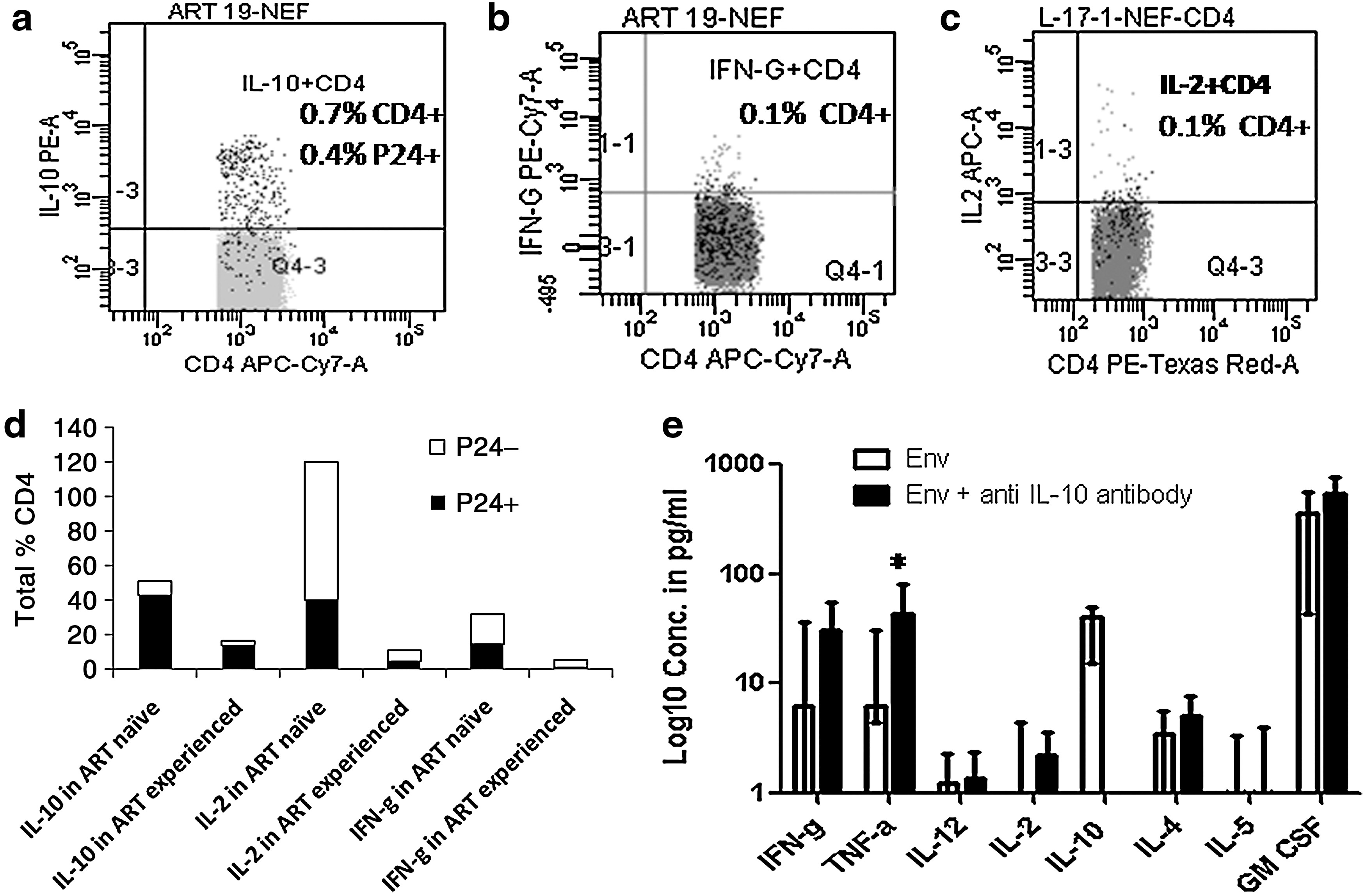
The assessment of the cytokine expression pattern of HIV-infected (P24 expressing) and HIV-uninfected (P24-negative) CD4 cells showed that all the three cytokines were secreted by HIV-uninfected as well as HIV-infected cells showing retention of functionality of the cells even after HIV infection. However, interestingly IL-10 was secreted predominantly by HIV-infected cells (Fig. 3d). It has been observed previously that IL-10 expression was 155-fold higher in infected (P24+) cells than in uninfected cells after in vitro infection of PBMCs. 48 Mitogen-stimulated PBMCs from HIV-infected individuals have also been shown to produce more IL-10 than PBMCs from HIV-negative controls 49 Plasma IL-10 levels have been observed to increase in parallel with viremia in patients who interrupted therapy after therapeutic vaccination and a population of circulating HIV-1-specific IL-10-secreting cells was expanded within the CD4+ T cell subset. 50 Hence there it might be possible that HIV-infected CD4 cells could have contributed to this IL-10 secretion. IL-10 has been found to be induced by HIV-1 Tat in mice 51 as well as in human monocytic cells, 52 which could partially explain IL-10 secretion by HIV-infected cells. IL-10 is known to suppress the immune response 53 and contribute to viral persistence. 54 Our study showed that HIV-infected CD4 cells could themselves be a source of IL-10 and may play a role in their own persistence.
CD8 response was also assessed in ART-naive patients by determining intracellular cytokines (IL-2, IFN-γ) secreted by CD8+ cells as well as expression of the degranulation marker (CD107a) as a marker of cytolytic activity. It was observed that all antigens were responsible for eliciting the CD8 response (Fig. 4b). Although IL-2 was found to be secreted by CD8 cells more or less to same extent (0.2–4.6%) after stimulation with all the antigens, the magnitude of IFN-γ and CD107a expression was higher after stimulation with Gag [0.1–1.8% (n=8/17) and 0.2–1.9% (n=4/17), respectively] and Nef pools [ranging from 0.1–1.4% (n=6/17) and 0.2–2.3% (n=3/17), respectively] than with Env [0.2–0.6% (n=3/17) and 0.2–0.3% (n=2/17), respectively] and Pol [0.1–0.9% (n=6/17) and 0.1–1.1% (n=2/17), respectively]. Gag and Nef have also been reported in previous studies to induce IFN-γ and CD107a secreting CD8 responses more frequently than Env and Pol peptides using similar assays. 55,56 Similar percentages of polyfunctional CD8 as defined by CD8 cells expressing two or more of these markers were significantly higher after stimulation with Gag and Nef (p 0.05) antigens than those with Env in these patients (Fig. 4c).
Polyfunctional T cells are considered important in controlling HIV infection as they were preferentially observed in long-term nonprogressors (LTNP) 57 and also in vaccinated macaques with better control of the disease. 58 The reason for less IFN-γ and CD107a expression as well as a smaller number of polyfunctional T cells in response to Env and Pol antigens could be the defective help provided through CD4 cells as more numbers of CD4 cells responding to these antigens were HIV infected and IL-10 secreting.
To determine the effect of IL-10 blockade in improving cytokine secretion, cells were incubated with HIV-1 Env with and without anti-IL-10 antibody (BD: 10 μg/ml) for 3 days. The cytokine concentrations were determined in the supernatants by the Bioplex protein array system using Th1/Th2 cytokine kit (Cat. No. M50-00005L3, Bio-Rad, USA). Most of the cytokines tended to increase after IL-10 blockade, although this was not significant statistically, except for tumor necrosis factor (TNF)-α, which increased significantly (p=0.047) (Fig. 3e). IL-10 blockade has also been shown to improve CTL responses in different previous studies done in vitro as well as in vivo. 50,59 Hence IL-10 blockade should be considered while attempting activation of the HIV reservoir to counter the immunosuppressive effect of IL-10 and to improve CTL response, which would help in the elimination of HIV-infected cells.
ART-experienced patients also showed findings similar to ART-naive patients, although the percentages of P24 expressing CD4 cells as well as cytokine-secreting CD4 and CD8 cells were comparatively less (data not shown). This was expected because HIV-specific immune response decreases after initiation of ART in the absence of continuous antigenic stimulation and also the frequency of resting CD4+ T lymphocytes harboring latent HIV-1 decreases in a biphasic fashion with an initial rapid decay within 3 months and a slower phase of decay thereafter. 60 Since CTL response decreases significantly after starting ART, CTLs may not play any role in controlling HIV-infected cells with replicating viruses in HIV-infected patients on prolonged ART. Hence, such interventions for reactivating HIV infection, if attempted soon after viral suppression but before waning of the CTL response, might prove helpful in the better control of HIV replication.
Thus, it was seen that HIV antigens could activate HIV replication in CD4 cells including resting CD4 cells as well as in CD4 cells representing a cellular reservoir. HIV replication was observed to be a very rapid process starting within half an hour after activation, indicating a stringent need for maintaining effective concentrations of antiretroviral drugs or even intensification of therapy to prevent further spread of HIV. Use of antigens for such a purpose may provide additional advantages as they may also induce a CTL response, which will help in the clearance of virally infected cells, as may happen with therapeutic vaccinations. It was observed that Pol and Env were responsible for maximal P24 expression versus Gag and Nef, which induced polyfunctional responses after stimulation. However, since the CTL response diminishes after prolonged ART, reactivation of latent infection should be attempted soon after viral suppression. Also after activation, HIV-infected cells were found to be responsible for secretion of IL-10, which may affect the CTL response, ultimately resulting in immunosuppression. IL-10 blockade was found to result in increasing concentrations of cytokines, especially TH1, and may be used along with therapeutic vaccination to improve the CTL immune response.
Footnotes
Acknowledgments
We would like to thank the laboratory staff, especially Ms. Poonam Kadam, Ms. Sujata Sahoo, and Subodh Shukre, and the clinic staff, including Ms. Prajakta Tayade, Ms. Ayesha Momin, and Mr. Narayan Panchal, for their help during the experiments and for recruiting patients in the study. We specially thank the study participants for their time and participation in this study. This study was supported by intramural funding from the Indian Council of Medical Research.
Author Disclosure Statement
No competing financial interests exist.