
Abstract
HIV-1 is capable of mimicking the ligand of integrin α4β7 by displaying a tripeptide mimotope on the V2 region. Through this mimicry HIV can bind the α4β7 integrin and get carried through the lymphocyte proliferation signaling pathway, cell-to-cell adhesion and can migrate to gut-associated lymphoid tissues. The same tripeptide motif was suggested to be the epicenter of neutralization in laboratory strains of HIV-1. In this study, we compared the α4β7 binding sites of two HIV-1 subtypes prevalent in China and found that the tripeptide binding domain of α4β7 was more diverse in subtype B’ strains than in CRF07_BC.
I
There is a mounting body of evidence suggesting that HIV-1, among other viruses including simian immunodeficiency viruses (SIV), rotavirus, and cytomegalovirus (CMV), mimics the signaling machinery and transport-mediating ability of integrin to bind lymphocytes. In addition, the composition of the amino acids at the surface of HIV-1 gp120 can impact the conformational structure of the viral envelope and the propensity to trigger the neutralizing immune response. 3,5,6 Intriguingly, the activated form of α4β7 integrin on the surface of CD4+ T cells can bind to both its natural ligand and the conserved tripeptide domain of HIV-1 glycoprotein gp120. This binding plays an important role in the early phases of HIV-1 infection regardless of the route of transmission. 2,3,7 During this stage, infected CD4+ lymphocytes migrate into the lamina propria where the replication of HIV-1 yields a high viral load and results in a massive depletion of CD4+ cells. 8 The center of the mimicry is located in a conserved tripeptide domain of the HIV gp120 V2 loop, which resembles natural ligands of the α4β7 integrin. The conservancy of the integrin α4β7 ligand and its molecular similarity with the host imply that HIV exploits the integrin–ligand binding mechanism to pass though the mucosa. In this manner, HIV can elude detection by the human immune system to migrate from the gut or the genital tract to become systemic. The mechanism of HIV infection through this alternative, possibly complementary HIV-CD4 receptor pathway remains unclear. In addition, the distribution of the tripeptide binding site of the integrin α4β7 on the surface of different HIV-1 subtypes and circulating recombinant strains has thus far not been described. To achieve this, it would require a dataset of nonredundant sequences of each subtype and circulating recombinant forms with associated epidemiological information. In this study, we compared the V2-V4 region of two clades (CRF07_BC and B’) prevalent in China to determine whether there is a difference in conservancy of the α4β7 integrin-specific mimotope on the surface of the HIV envelope.
We selected a total of 275 stored blood samples previously collected between 2003 and 2007 from HIV-1-infected individuals in Henan, Shanxi, Sichuan, and Xinjiang provinces in China (Fig. 1). The risk factor for HIV transmission was known for each patient. Samples from Henan and Shanxi (n=48 and 63, respectively) were obtained from former blood donors infected with HIV through unregulated unsanitary commercial blood collection, 9 while samples from Sichuan and Xinjiang provinces were obtained from intravenous drug users (n=28 and 136, respectively). All blood samples were originally tested by ELISA (BioMerix, France) and confirmed HIV-1 positive by Western blot (Genelabs Diagnostics, Singapore) at their respective provincial laboratories. The study was approved by the institutional review board of the National Center for AIDS/STD Control and Prevention (NCAIDS, China CDC) and was conducted in accordance with human experimentation guidelines.

Map of China. The provinces where samples were collected are shown in gray. The number of samples collected in each province is shown. The subtype of samples collected in each province is shown in bold.
The RNA and DNA were extracted from plasma and whole blood, respectively, using QIAGEN extraction kits following the manufacturer's instructions (QIAGEN, Co., Ltd, Hilden, Germany; RNA: QIAamp Viral RNA Mini Kit; DNA: QIAamp DNA Blood Mini Kit). The synthesis of cDNA from RNA was carried out using 2.5 units of avian myeloblastosis virus reverse transcriptase in a 25 μl reaction mixture containing 2.5 μl 10×One Step RNA PCR buffer, 5 μl MgCl2 (25 mM), 2.5 μl dNTP mixture (10 mM), 0.5 μl RNase inhibitor (40 units/liter), and 0.5 μl primer (20 μM). 10 All subsequent cDNA and DNA amplifications were performed as previously described by Liu et al., using the TaKaRa Ex-Taq (Takara Bio Inc, Tokyo, Japan). 10 PCR amplicons were sequenced using the ABI 3100 Genetic Analyzer with the BigDye Terminator chemistry (BigDye V3.1; Applied Biosystems, Foster City, CA). The nucleotide sequences from this study are available in GenBank under accession number HQ292336–HQ292610.
Sequences generated by this study encompassing the V2-V4 region were aligned with HIV-1 reference sequences representing all previously documented subtypes (A, B, C, D, F, G, H, J, K) and circulating recombinant forms CRF07_BC and CRF08_BC, using the GeneCutter tool available at the Los Alamos HIV sequence database (
As depicted in Fig. 2, all 275 samples were classified into two well-defined clades with significant bootstrap values (>75%). Consistent with previous molecular epidemiological surveys of HIV-1 in China, 11 sequences from former blood donors in Henan and Shanxi provinces formed a single subcluster of the B subtype, and were denoted as subtype B’. In addition, as shown in two previous reports, 12,13 sequences obtained from intravenous drug users in Sichuan and Xinjiang clustered with CRF07_BC reference sequences (Fig. 2). This preliminary analysis provided us with two distinctly different phylogenetic subtypes of HIV-1 to perform a detailed cross-clade comparison of the V2-V4 region of subtype B’ and CRF07_BC. As previously stated, the V2-V4 region encompasses the putative α4β7 mimotope domain, which mimics the binding site of integrin. The rationale, results, and possible applications for each comparison are outlined as follows.

Neighbor-joining tree of the V2-V4 sequences. The phylogenetic tree was constructed using the neighbor-joining method with the F84 substitution model. Filled circles mark the B’ sequences sampled from Henan, open circles mark the B’ sequences sampled from Shanxi, filled triangles mark the CRF07_BC sequences sampled from Sichuan, and open triangles mark the sequences from Xinjiang. The reliability of nodes was tested using the bootstrap method with 1000 replicates. The bootstrap values for different clades were B’: 98%; B: 89%; D: 95%; 07_BC: 86%; 08_BC: 81%; C: 100%; K: 100%; F: 68% (F1: 93%; F2: 97%); H: 100%; G: 100%; J: 100%; A: 99% (A1: 91%; A2: 100%).
First, we estimated the evolutionary distance between the two clades to determine whether the difference within the α4β7 mimotope domain of B’ and CFR07_BC reflects the overall difference between the two clades. A phylogenetic analysis using the nucleotide sequences within the V2-V4 regions revealed that the branch lengths between operational taxonomic units (OTUs) of B’ and their common ancestral node (CAN) were significantly higher than those of the CRF07_BC OTUs and their respective CAN. Indeed, the difference of median distances between OTUs to their respective clade CAN was statistically significant (Fig. 3A, Mann–Whitney test, p<0.0001). This suggested that the HIV-1 subtype B’ epidemic in Shanxi and Henan provinces was more heterogeneous than in Xinjiang and Sichuan where CRF07_BC constitutes the predominant circulating viruses.
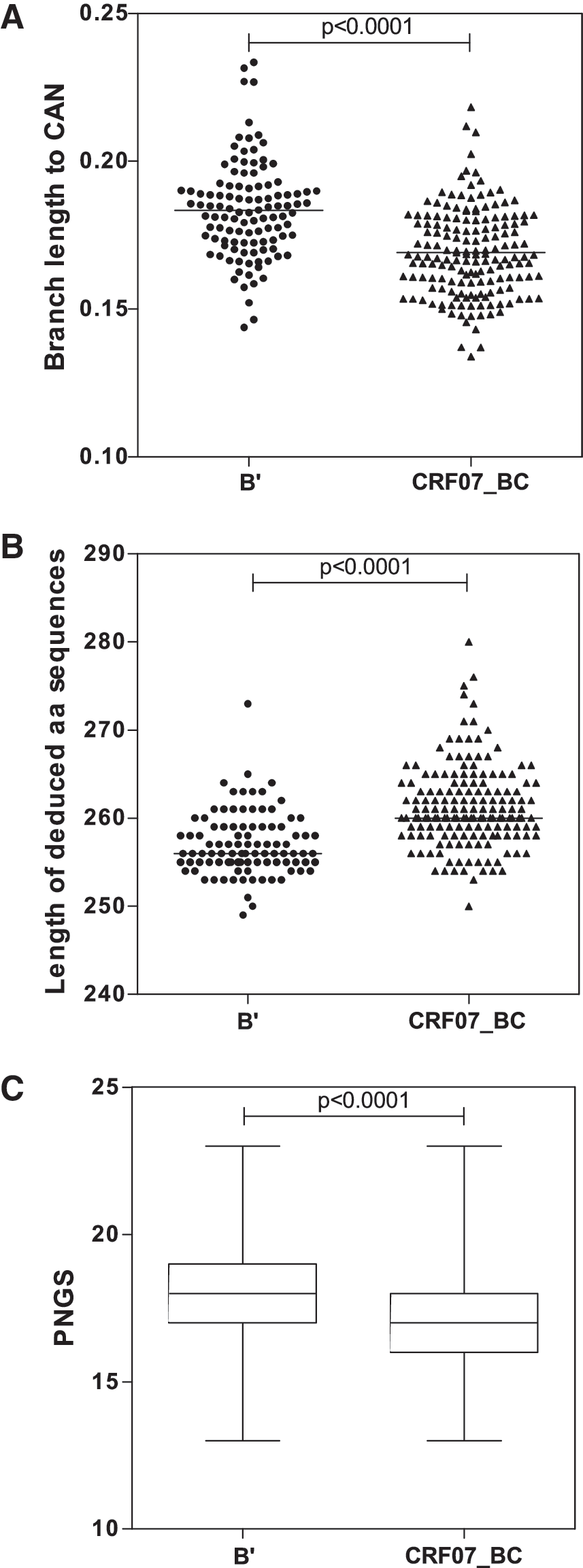
Comparative analysis of
Second, the composition of the amino acid within the envelope of HIV-1 can impact the conformational structure and the propensity of the virus to trigger the neutralizing immune response. 5,6,14 In addition, insertions and deletion-type mutations could significantly affect the function of the viral envelope and its ability to bind cellular receptors. Therefore, the lengths of the deduced amino acid sequences of the V2-V4 regions were determined and analyzed using the Mann–Whitney statistical test. As shown in Fig. 3B and C, there was a statistically significant difference between subtype B’ and CRF07_BC when the length of the deduced amino acid sequences and the number of putative N-glycosylation sites were compared (p<0.0001). This may suggest that the conformational structure of the two subtypes may be different, and could probably have some relevance in virus–cell interaction.
Finally, the analysis of tripeptides at positions 179–181 relative to the envelope amino acid sequence of HIV-1 HXB2 revealed that of 111 sequences classified as subtype B’, 51% displayed a leucine, an aspartic acid, and an isoleucine (LDI, Leu-Asp-Ile) and 28% displayed a leucine, an aspartic acid, and a valine (LDV, Leu-Asp-Val) within the putative integrin α4β7 ligand site (Fig. 4A). In contrast, 95% of sequences classified as CRF07_BC displayed LDI at positions 179–181. There was little variation in the CRF07_BC integrin binding site, which displayed only three alternative tripeptide motifs at the 179–181 domain (Fig. 4A). Conversely, there was a higher degree of heterogeneity for subtype B’ sequences with 16 different tripeptide variants in the integrin α4β7 binding domain. To quantify the degree of the diversity in the 179–181 domain, the Shannon diversity indices 15 were calculated. The Shannon diversity index for subtype B’ was higher (4.272) than that of CRF07_BC (1.278), suggesting a marked difference in the composition of the putative mimotope of the α4β7 domain in the V2-V4 region of the HIV-1 subtype B’ and CRF07_BC (Fig. 4A). The diversity estimation between subtypes B’ and CRF07_BC in the 179–181 domain was comparable with the analysis of a single amino acid at positions 179 and 181 (Fig. 4B and C, respectively). The conservation of the integrin binding site in HIV-1 CRF07_BC may be indicative of an adaptation of CRF07_BC in intravenous drug users in Xinjiang and Sichuan provinces. However, all CRF07_BC (n=164) and 110/111 of subtype B’ sequences contained an aspartic acid at position 180. This aspartic acid residue was suggested to be critical for the neutralization resistance phenotype in HIV-1 strains in vitro. 16
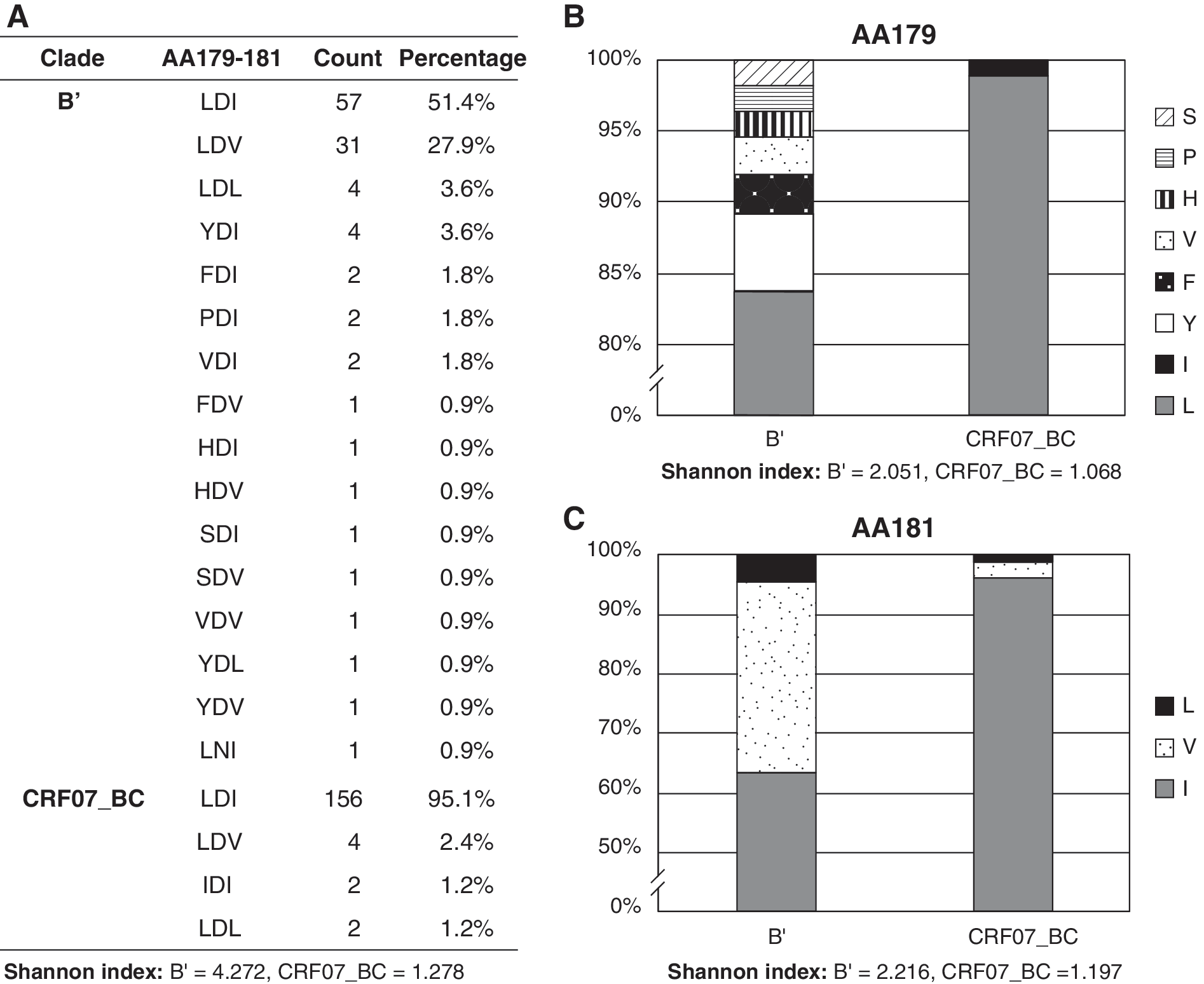
Comparative analysis of the amino acid composition of the putative binding site of the α4β7integrin.
The central hypothesis of this investigation was that the putative mimotope of the integrin α4β7 could differ depending on the subtypes of HIV-1 strains and their propensity to infect host cells. It has been shown that the process by which lymphocytes migrate to gut-associated lymphoid tissue is mediated by integrin α4β7. 3 HIV exploits this process by mimicking the natural ligand of integrin, which allows HIV to bind the surface of lymphocytes in order to migrate into the general circulation. During this process, HIV could infect CD4+ cells through the traditional virus–receptor pathway. However, the mechanism of HIV infection through the integrin and other alternative pathways has yet to be elucidated. Previous studies have suggested that different HIV-1 subtypes could have different selective pressures against the envelope transmission and during the course of infection. 6,17 The V1-V2 domain of the envelope has been shown to contain epitopes that confer the neutralization sensitive or resistant phenotype to HIV strains. 5,14,18 –21 Most recently, O'Rourke et al. 16 showed that the substitution of aspartic acid (D) with asparagine (N) at the center of the recognition motif of α4β7 (LDI/V) in V2 can drastically change the viral phenotype from resistant to becoming sensitive to neutralization. Unlike most laboratory strains, which can readily be blocked in vitro by recombinant soluble CD4 and CD4-immunoglobulin hybrid molecules, wild-type HIV-1 primary isolates are naturally resistant to both cellular and humoral neutralization. 18,22 –24 Envelopes of the majority of strains that are responsible for the global HIV-1 pandemic have evolved to resist antibody-mediated neutralization, exhibiting a tier 2–3 level of resistance against heterologous plasma pools and monoclonal antibodies. 25 Most of these viruses contain an aspartic acid residue at the center of the ligand of α4β7 integrin; however, this is also true for their more sensitive tier 1 counterparts. Furthermore, in studies of neutralization escape from autologous antibodies in natural infection, the aspartic acid is conserved and does not appear to play a direct role in this process. 26 –28 Indeed, substitution of this residue is very rare in known HIV-1, HIV-2, and SIV sequences, and often leads to a decrease in viral infectivity in vitro. 16 Whether in natural infection the aspartic acid at the center of the ligand of α4β7 integrin could confer HIV-1 the antineutralization phenotype remains unclear. Assuming that in real life the aspartic acid at position 180 is indeed responsible for the resistance to neutralization as described by O'Rourke et al., 16 it should be expected that the majority of HIV-1 strains that have established a successful epidemic in China and worldwide should have an aspartic acid at this critical V2 position. All but one of the 275 sequences of this study contained an aspartic acid at position 180, consistent with the existence of strict structural constraints on this region of the V2 domain.
Taken together, the data suggest that neutralization resistance is determined by multiple factors in addition to the aspartic acid at position 180, but this residue may be involved in maintaining envelope conformation. Therefore, new vaccines designed to use the α4β7 motif in V2 to trigger neutralization sensitivity should take into account not only the high conservation of aspartic acid but also the differences in the other residues across different subtypes.
Footnotes
Acknowledgments
This work was supported by the Chinese 973 Program (2005CB522903) and International Cooperative Grant (2009DFB30420) from the Ministry of Science and Technology of China. Additional support was provided by the National Major Projects for Infectious Diseases Control and Prevention (2008ZX10001-004, 010) from the Ministry of Health of China. The authors would like to thank Professors Robin Ryder and Cynthia Derdeyn, Dr. Juris Grasis, and Neville Kisalu for invaluable comments and critical review of the manuscript.
The author(s) have made the following declarations about their contributions: Conceived and designed the experiments: X.H., K.H., J.C., Y.S.; performed the experiments: R.X., Y.H.S., M.J., J.C., Y.F., H.X.; analyzed the data: X.H., J.L.M., J.W., Y.F., Y.S.; wrote the paper: J.L.M., X.H., Y.S. All authors reviewed and agree with the final content of the paper.
Author Disclosure Statement
No competing financial interests exist.