
Abstract
The human immunodeficiency virus type 1 (HIV-1) characteristics associated with mother-to-child transmission (MTCT) are still poorly understood. We studied a cohort of 30 mothers from Rwanda infected with HIV-1 subtype A or C viruses of whom seven infected their children either during gestation or soon after birth. CD4 counts and viral load did not significantly differ between nontransmitting mother (NTM) versus transmitting mother (TM) groups. In contrast to earlier studies we not only analyzed and compared the genotypic characteristics of the V1-V5 region of the gp120 envelope of viruses found in TM and their infected children, but also included data from the NTM. No differences were found with respect to length and number of potential N-glycosylation sites (PNGS) in the V1-V2 and the V1-V5 region. We identified that viruses with a PNGS on positions AA234 and AA339 were preferably transmitted and that viruses with PNGS-N295 showed a disadvantage in transmission. We also showed that the frequency of PNGS-N339 in the viruses of TM and infected children was significantly higher than the frequency in NTM in our cohort and in viruses undergoing sexual transmission while the frequency of PNGS-N295 in children was significantly lower than the frequency in TM and acute horizontal infections. Collectively, our results provide evidence that the presence of the PNGS-N339 site and absence of the PNGS-N295 site in the gp120 envelope confers an advantage to HIV-1 when considering MTCT.
Introduction
T
A number of studies focused on analyzing the molecular characteristics of viruses that get transmitted, either through MTCT or sexual intercourse, show that the majority of infections are characterized by the presence of a highly homogeneous viral population. 2 These viruses are predicted to exclusively utilize the CCR5 coreceptor (termed R5 viruses) for infection even when the mothers are shown to carry CXCR4 using variants (termed R5/X4 or X4 viruses), 3 indicative of a strong R5 selection pressure. This is somewhat surprising in children given the predominance of CXCR4 expressing naive CD4+ lymphocytes in the circulation, a consequence of limited antigen exposure. 4,5
Virus selection during MTCT may be influenced by the binding affinity of the gp120 envelope glycoprotein to a number of cell receptors. A higher affinity for the CD4 receptor or the CCR5 coreceptor may be of advantage for the infecting virus particle. A low net charge within the V3 variable loop of gp120 and the presence of a potential N linked glycosylation site (PNGS) on position 301 of the V3 loop 6 –8 are variables that have been associated with higher CCR5 affinity and being the variants preferentially transmitted. 9,10
In MTCT the HIV-1 variants transmitted to children tend to be more resistant to maternal plasma at time of delivery in comparison to viruses from the mother. 11,12 Some studies suggest that the properties and function of maternal antibodies (Abs) in MTCT can prevent perinatal transmission or mediate viral selection. 13 A number of gp120 envelope characteristics have been associated with resistance against neutralizing Abs, such as the V1-V2 region of the envelope, which has been postulated to regulate neutralization sensitivity by occluding conserved epitopes such as the CD4 and/or coreceptor binding sites. 14,15 The V1-V2 length of subtype C viruses has been shown to positively correlate with resistance to broadly cross-neutralizing sera. 16 –21
It has been postulated that HIV-1 transmission can be enhanced by binding of the gp120 glycan moiety to the C-type lectin DC-SIGN, which is expressed at high levels on immature dendritic cells (DCs) and facilitates infection of CD4+ T lymphocytes either locally or in distal lymph nodes. 22 –24 DC-SIGN expressed by a subset of B cells in the tonsils and blood may also contribute to the infection of CD4+ lymphocytes. 25 If this mode of transmission plays a role in MTCT, selection of the transmitting virus may be driven by more efficient binding of the virus to DC-SIGN.
Here we study a group of 30 pregnant mothers from Rwanda infected with either HIV-1 subtype A or C viruses of whom seven transmitted virus to their children. No correlation was identified between viral loads or CD4+ lymphocyte counts for transmitting mother (TM) or nontransmitting mother (NTM). We analyzed the gp120 envelope sequences for TM, NTM, and infected children. From this analysis we identified two potential glycosylation sites, PNGS-N234 and PNGS-N339, which were associated with viruses that are preferentially transmitted from mother to child and a disadvantage in transmission for viruses with PNGS-N295.
Materials and Methods
Study population
This is a subanalysis as part of a clinical study performed in Kigali, Rwanda as previously described. 26 In brief, the study is composed of a group of 30 HIV-1-infected women from Rwanda and their children. Plasma samples were obtained from both mothers and children at time of delivery and at week 16. Seven children tested HIV-1 positive at 16 weeks after delivery. All the women were given a single dose of nevirapine at the start of labor to reduce peripartum transmission of HIV-1. All children were breastfed during the 16-week period.
Viral load and CD4+ cell count assessment
Blood plasma samples were analyzed for HIV-1 viral loads using the Roche diagnostics RNA PCR quantification assay version 1.5, with a lower limit of detection of 400 copies/ml. Immunologic testing of mothers was performed in two laboratories (National HIV/AIDS Reference Laboratory, Kigali, Rwanda and the Joint Clinical Research Center JCRC, Kampala, Uganda).
HIV-1 RNA extraction
HIV-1 RNA was isolated from blood plasma samples according to the method of Boom et al. 27
Children's infection status at birth and week 16
Plasma samples taken at birth were tested in a reverse transcriptase polymerase chain reaction (RT-PCR) amplification assay covering the HIV-1 gp120 V3 region. Initially, viral RNA was converted to cDNA using Superscript III Reverse Transcriptase (Invitrogen) and the 3′ primer V3Not (GCG CGG CCG CCC CCT CTA CAA TTA AAA CTG TG), followed by PCR using Taq DNA Polymerase (Roche), 3′ primer 3′ V3Not and 5′ primer 5′ V3Not (GCG CGG CCG CAC AGT ACA ATG TAC ACA TGG). A nested PCR was subsequently performed using 5′ primer 5′ KSI (ATA AGC TTG CAG TCT AGC AGA AGA AGA) and 3′ primer 3′ KSI (ATG AAT TCT GGG TCC CCT CCT GAG GA) and the PCR product was analyzed by agarose/ethidium bromide electrophoreses. At week 16 the children's HIV-1 infection status was determined using standard viral load assessment as described above.
Amplification of the V1-V5 envelope region
To amplify the V1-V5 envelope region viral RNA was converted into cDNA using the Superscript III Reverse Transcriptase Kit (Invitrogen) and 3′ primer A1262 (CTG ACG GTA CAG GCC AGA CAA TTA TTG TC) followed by a PCR using the Expand Long Template PCR system (Roche), according to the manufacturer's instructions. For the first PCR 5′ primer A1053 (GAA AGA GCA GAA GAC AGT GGC AAT GA) and 3′ primer A1262 were used, and for the second PCR the 3′ primer was A1322 (TCT TGG GAG CAG CAG GAA GCA C) and the 5′ primer was A0358 (GAG GAT ATA ATC AGT TTA TGG GA).
Sequencing and analysis
PCR products were sequenced using the ABI PRISM Big Dye Terminator 1.1 Cycle Sequencing Kit and the ABI 3730 XL DNA Analyzer (Applied Biosystems). Sequences were assembled using the CodonCode Aligner program (CodonCode Corporation). Alignments were performed taking the translation codons into account and the sequences were translated into amino acids using the BioEdit program (Ibis Therapeutics). Analysis of the amplified region was performed utilizing the neighbor-joining (N-J) method of MEGA version 4. Positions containing an alignment gap were included for pairwise sequence analysis.
Subtype determination
To determine HIV-1 subtype the V1-V5 envelope PCR products were sequenced using primers 5′ A0385 (GAG GAT ATA ATC AGT TTA TGG GA), 3′ A1322 (TCT TGG GAG CAG CAG GAA GCA C), 5′ A1360 (GAG CCA ATT CCY ATA CAT TAT TG), and 3′A1191 (ATG GGA GGG GCA TAC ATT GC). The Subtype Reference Alignments of the Los Alamos Database (
Cloning and sequence analysis
V1-V5 envelope PCR products were cloned into the pCRII-TOPO vector (Invitrogen). For each subject and time point between 5 and 15 clones were sequenced using the primers 5′ SP6 (ATT TAG GTG ACA CTA TAG), 3′ T7 (TAA TAC GAC TCA CTA TAG G), 5′ A1360, and 3′ A1191. The number of PNGS was determined using the tool N-GlycoSite of the HIV sequence Database (
Direct sequencing of the C2-C3 region
To expand the group of NTM for comparison of the frequency of PNGS-N295 and PNGS-N339 we performed direct sequencing on the V1-V5 envelope PCR products of 10 additional mothers encompassing amino acids 245 to 370 (HXB2 numbering) using 5′ primer (GCG CGG CCG CAC AGT ACA ATG TAC ACA TGG) and 3′ primer (AT TTC TAA GTC CCC TCC TGA).
Comparison of the frequency of PNGS-N295 and PNGS-N339
The percentages of glycosylation on pos. AA295 and AA339 (HXB2 numbering) for subtype A and C were calculated separately for the envelope sequences from the group of NTM, children infected in uterine (C2),
29
children infected by breastfeeding (C3),
30
breastfeeding TM (TM2),
31
acute horizontal infections (contr.1),
32
–34
and a random selection of chronic infections acquired by horizontal transmission from the Search Interface of the HIV Sequence Database (contr.2) (
Results
High viral loads and low CD4+ T cell counts were not the only determinants for HIV-1 transmission
Viral loads and CD4+ T cell counts were measured and compared for the 23 NTM versus seven TM (Table 1) (Fig. 1). Four mothers gave birth to HIV-1-positive children as determined by positive HIV-1 RT-PCR reactions. This indicates three of 26 mothers (11.5%) were infected either intrapartum or during early breastfeeding, fitting with known HIV-1 transmission rates when mothers are given nevirapine at time of delivery. 36 The TM were not found exclusively in the group with high viral loads and low CD4+ T cell counts. Three out of the seven TM had CD4+ cell counts >500 cells/μl, and three of seven had viral loads lower than 10,000 copies/ml, with one mother in both categories. These data suggest that factors other than viral load and CD4+ cell counts can influence HIV-1 transmission.

Comparison of CD4 T cell counts and viral loads in transmitting mothers (TM) and nontransmitting mothers (NTM). The CD4 T cell counts in cells/μl
TM, transmitting mothers; NTM, nontransmitting mothers who have been studied in detail.
Phylogenetic analyses of sequences of mothers and children
The V1-V5 region of the gp120 envelopes of NTM and TM was sequenced. Out of the seven TM two (29%) were infected with subtype C viruses while five (71%) were infected with subtype A viruses. Of the group of 23 NTM, six (26%) were infected with subtype C and 15 (65%) with subtype A viruses and for two mothers the virus subtype was undetermined.
We cloned and sequenced the V1-V5 regions of eight NTM and seven TM and their infected children. Phylogenetic analysis of the sequences of the V1-V5 regions of mothers and children confirmed the relatedness of the virus circulating in the respective mother–child pairs (data not shown). Four of the seven children were infected in utero of whom child 091 demonstrated two separate variants (Fig. 2A). All other infected children had only one HIV-1 variant while the viral quasispecies from the TM contained multiple variants (Fig. 2B). The intrapatient mean genetic distances of the V1-V5 region were analyzed (Fig. 3A). The mean genetic distance in NTM was 0.022 (95% CI 0.007–0.037; n=8) and was similar to that of TM [0.025 (95% CI 0.004–0.046; n=6)] (p=0.82) or from the children [0.007 (95 CI 0.003–0.012; n=6)] (p=0.12); however, the mean intrapatient diversity in the children was significantly lower than in the TM (p=0.007), which was in agreement with the transmission of one virus variant in six out of seven children and two variants from one child.
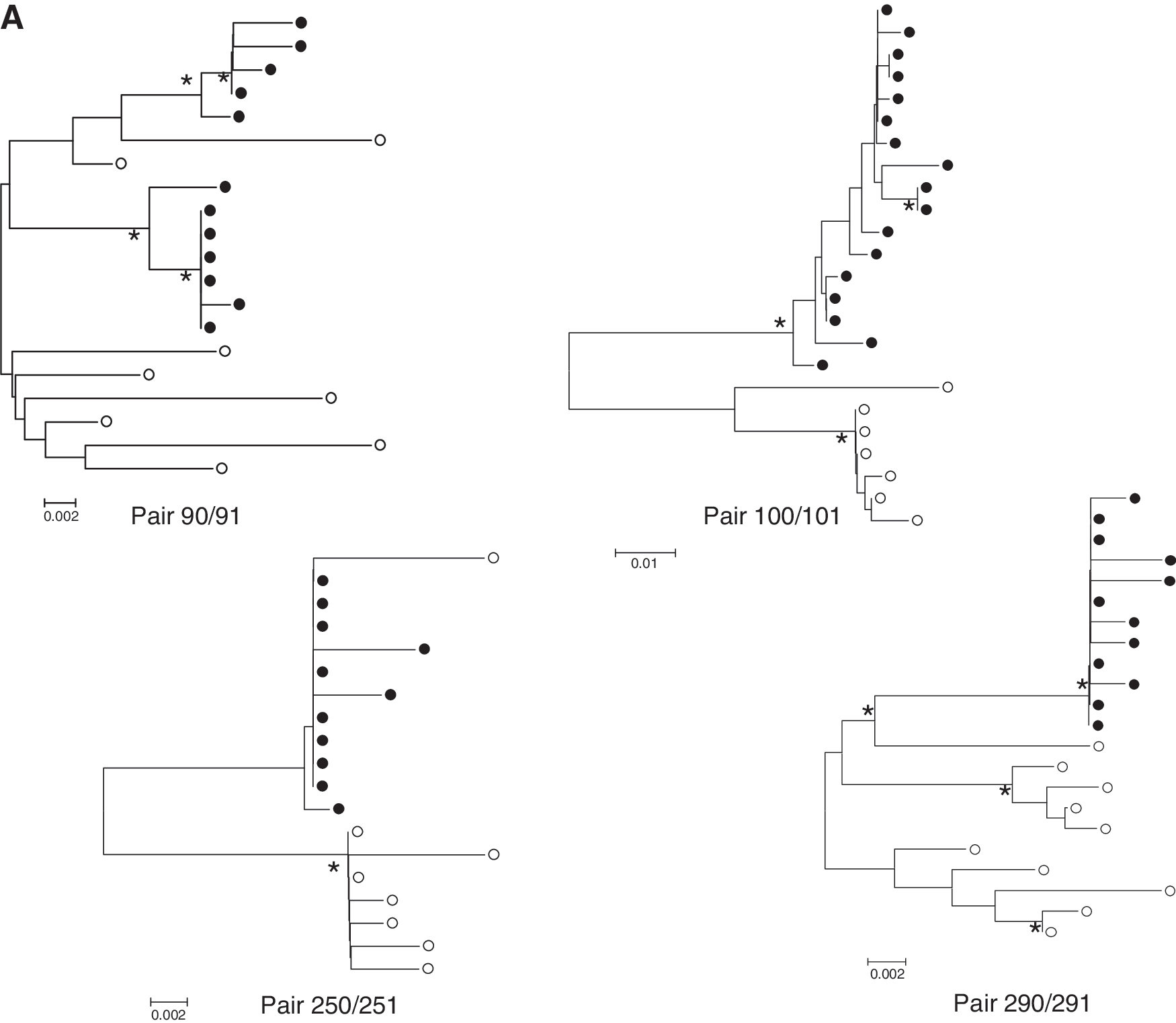
Phylogenetic analysis of V1-V5 envelope sequences. The unrooted neighbor-joining trees show sequences of seven mother–child pairs.
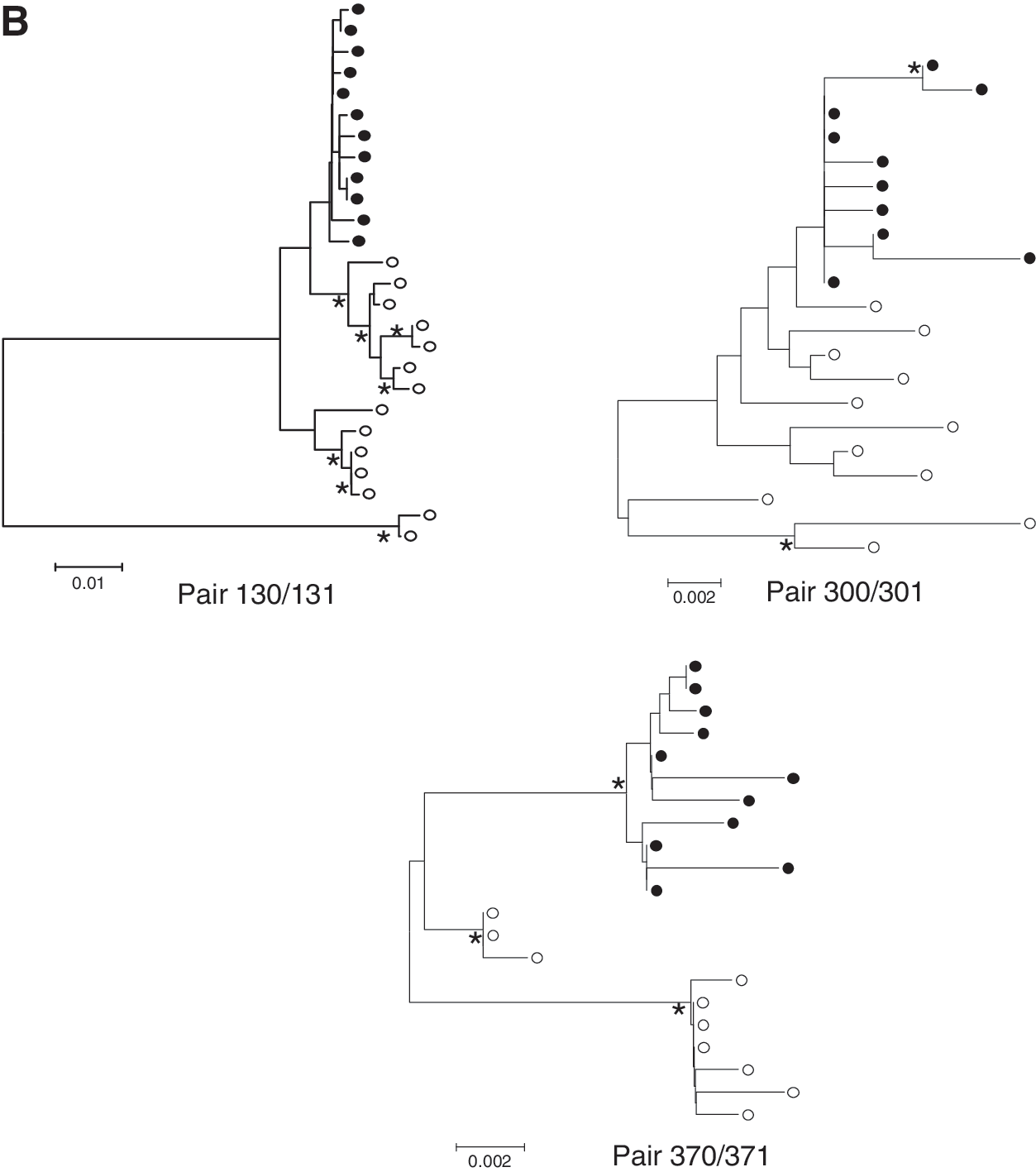
Viral characteristics between NTM, TM, and children.
V3 regions with a PNGS on position AA301 and a charge of +3 or +4 are transmitted
We next analyzed the number of PNGS and the overall V3 charge in the three groups. PNGS-N301 (HXB2 numbering), which is associated with CCR5 usage, was found in the majority of clones from TM (100% in 6/7 and 90% in 1/7) and NTM (100% in 6/8, 25% and 0% in 1/8), and was present in 100% of the clones of all children. The mean V3 charges of viruses from NTM varied between 1.89 and 4.03 (average 3.17) and for TM between 2.58 and 5.00 (average 3.69) and was found to be narrower at between 3.00 and 3.91 (average 3.40) in the children (Fig. 3B).
No differences in V1-V2 and V1-V5 length and number of PNGS
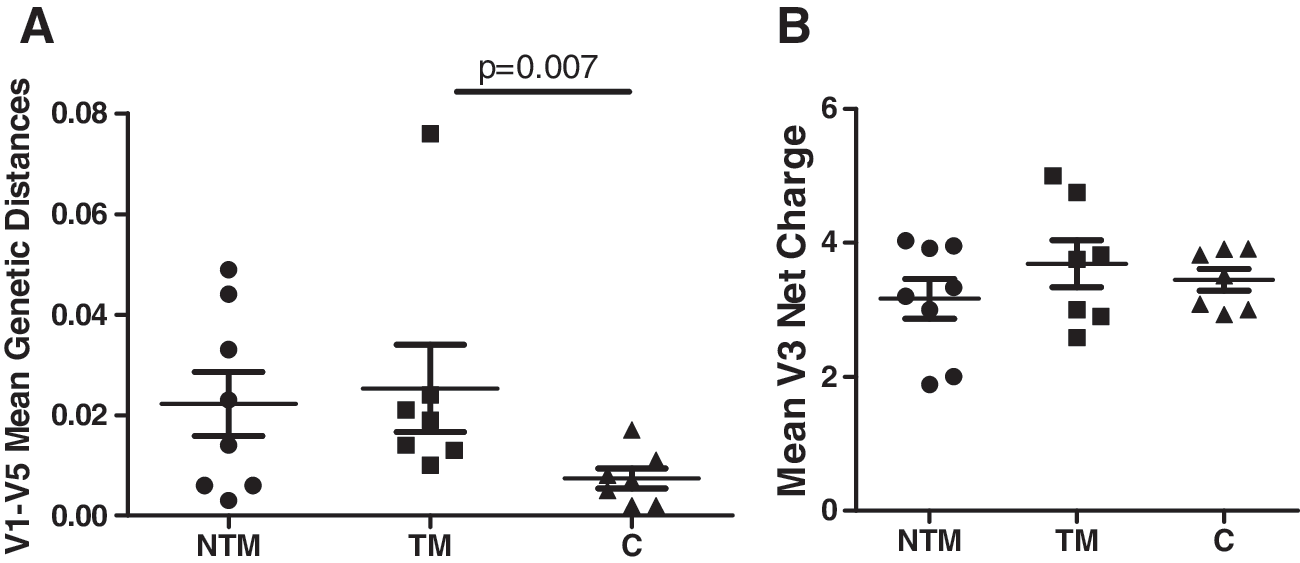
To identify differences between the virus variants in the NTM and TM of children we analyzed the envelope sequences. We compared the size and the number of PNGS of the V1-V5 region and the V1-V2 region. The mean length of the V1-V5 region of amplified products from the NTM was 342.1 (95% CI 336.6–347.5) amino acids, in the TM 342.3 (95% CI 337.0–347.5) amino acids (p=0.95), and in the children 340.0 (95% CI 334.8–345.1) (p=0.70) amino acids (Fig. 4A). The mean number of PNGS in the V1V5 region of the viruses in the NTM was 22.0 (95% CI 20.5–23.6), in the TM 22.2 (95% CI 20.6–23.7) (p=1.00), and in the children 23.0 (95% CI 21.2–24.8) (p=0.39) (Fig. 4B). The mean size of the V1-V2 of NTM was 73.0 (95% CI 66.4–79.7) amino acids, in the TM 68.8 (95% CI 64.9–72.8) (p=0.24) and in the children 68.6 (95% CI 64.5–72.7) (p=0.22). Overall, we did not find significant differences between the NTM, the TM, or the children in length and number of PNGS in either the V1-V2 or V1-V5 envelope regions.
V1-V5 envelope length and number of potential N-glycosylation sites (PNGS) in NTM, TM, and children. No significant differences in V1-V5 length
Selective transmission of viruses with a PNGS on N234 and N339
To further identify envelope PNGS differences we mapped the sites in NTM, TM, and children. Six PNGS were found conserved in all variants for all groups (HXB2 numbering N136, N156, N198, N262, N332, and N447), with three sites showing a difference between the groups (Table 2). PNGS-N234 (in C2) was present in all of the children's clones, in the majority of the clones of TM (86%), and less abundant in the NTM (62%). Only 60% of the clones of TM 290 had a PNGS at position N234, whereas 100% of her child's (291) clones possessed this PNGS, suggesting an advantage in transmission. PNGS-N295 (between C2 and V3) was present in all NTM clones, the majority of clones in four/seven TM, and in four/seven children, which would suggest a disadvantage for transmission; however, when the PNGS is present transmission can occur. Six of seven children demonstrated PNGS-N339 in all virus variants, which was present in the majority of clones from TM (five/seven) and was found in only two/eight NTM. PNGS-N339 was absent in all the clones of mother 370 and the corresponding child 371. Only 13% of the clones of mother 250 held PNGS-N339, whereas her child (251) had 100% of viruses carrying this PNGS. The above findings suggest that variants with PNGS-N234 and/or PNGS-N339 were preferentially transmitted and that variants with PNGS-N295 had a disadvantage.
# clones, number of clones with a PNGS/number sequenced.
PNGS, potential N-glycosylation sites.
To verify our above findings we compared our results with data from different sources (Search Interface of the HIV Sequence Database) where we could compare sequences of subtype A or C viruses from children infected in utero (C2/N=40), from children infected through breastfeeding (C3/N=40), as well as from mothers who transmitted the virus by breastfeeding (TM2/N=83). Data from the virus populations of NTM were not available in the database. We performed a direct sequence analysis of 10 additional NTM from our cohort to increase the overall study group size. Viral sequences from acute infection (AI) patients of subtype A and C were included to verify if this PNGS is associated with recently sexual intercourse-transmitted viruses (control 1/N=90). We also included a random selection of sequences from drug-naive African individuals of subtype A and C (control 2/N=318) to verify the prevalence of the PNGS at position 295 and 339 in individuals other than mothers or children (Fig. 5A and B, respectively). If more than one sequence of an individual was available we calculated the consensus of these sequences. We matched the percentages of individuals positive for the PNGS to equivalent proportions of subtype A and C in the groups of TM and children that we investigated (C1 and TM1). To determine significance we performed a Supervised Comparison of the Subjects with the children groups C1, C2, and C3 as the reference group for the G standard (Fig. 5).
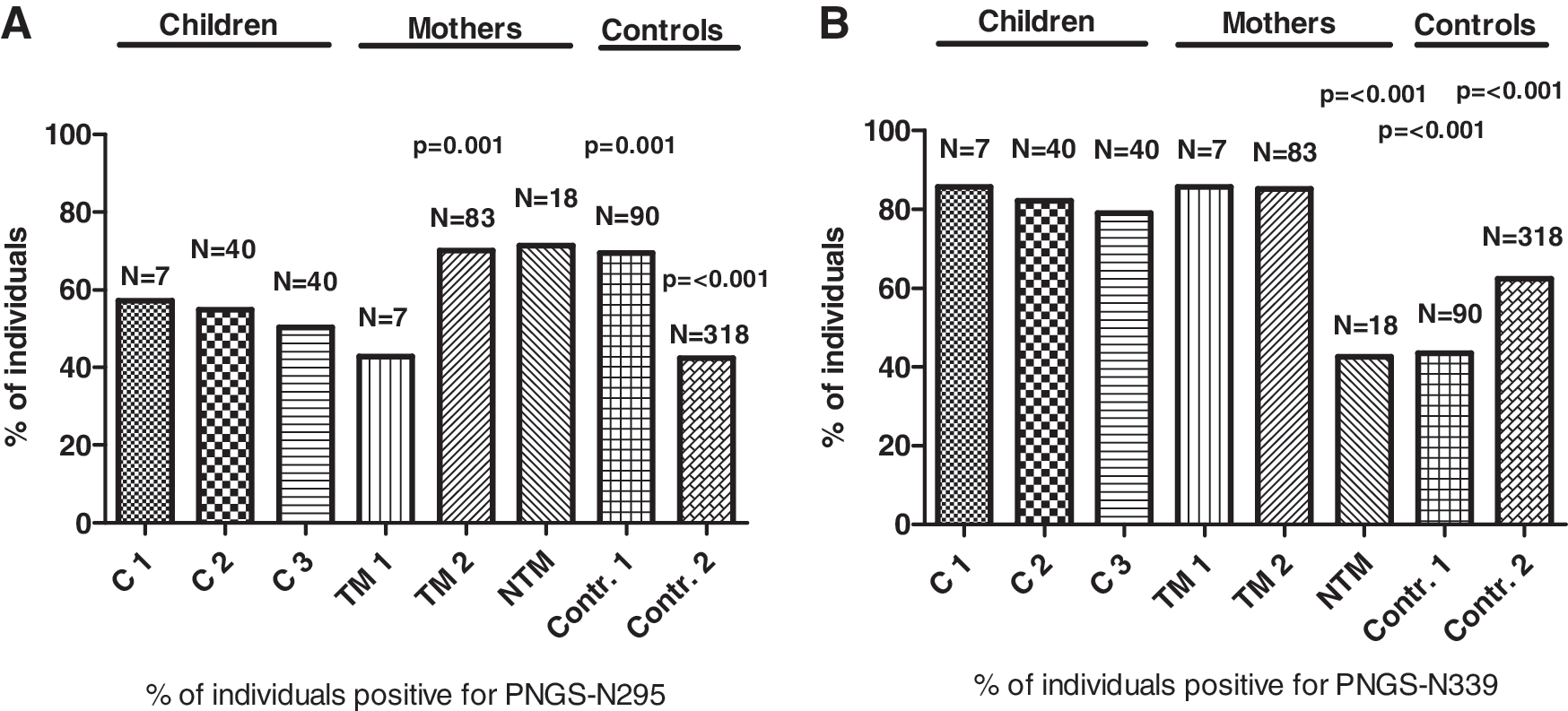
PNGS percentages at positions
Due to the fact that sequences of groups C2, C3, and TM2 did not encompass PNGS-N234 we could not analyze this PNGS. When analyzing further PNGS-N295 we found it to be present in 50–70% children (C1, C2, C3), which was significantly reduced when compared to TM2 (70%) and the acute horizontal transmissions (control 1) (69%) (both p=0.001), which is in concordance with a disadvantage in MTCT for envelopes carrying PNGS-N295 (Fig. 5A). For PNGS-N339 we found this site to be present in over 79–86% of the consensus sequences for both TM and children, whereas for NTM and control 1 and control 2 the frequency was significantly reduced (43%) (p≤0.001), 44% (p≤0.001) and 51% (p≤0.001), respectively, indicating a strong positive selection in MTCT for virus variants with a PNGS on position AA339 in the C3 region of the viral envelope (Fig. 5B).
Discussion
From our V1-V5 gp120 sequence analysis of viruses preferentially transmitted between mothers and their children and in comparison to nontransmitting viruses we identified a higher frequency of PNGS at positions AA234 and AA339 and a lower frequency of PNGS at position AA295 in transmitted viruses. We confirmed the results of preferential transmission of viruses with PNGS at position N339 and the disadvantage in transmission for viruses with PNGS-N295 in MTCT through analyzing additional cohorts of MTCT as well as in individuals in their acute infection period and infected through sexual intercourse. We should emphasize that for PNGS-295 the observation is more group based than at the individual transmission pair level.
In sexual transmission HIV-1 subtype A and C variants with shorter envelope and fewer PNGS have been shown to be more efficiently transmitted and more susceptible to neutralizing antibodies present within the donor. 37 –39 One report concludes that variants transmitted from mother to child did not have shorter V1-V5 regions but did have fewer PNGS, 11 while another study showed no association between transmitted variants and the length of the variable loops or the number of PNGS. 40 We did not find differences in the number and the length of the V1-V2 and the V1-V5 regions between NTM, TM, and children in our cohort. Probably resistant viruses are preferentially transmitted from mother to child but it is not clear how changes in length of the envelope and in the number of PNGS correlate with MTCT. 11,12,37,41,42
A low overall net charge and a PNGS at N301 within the gp120 V3 variable loop have been heavily associated with CCR5 usage. 7 In agreement with other reports that show selection of CCR5 using variants in transmission we found that all transmitted viruses contained PNGS-N301 and that the V3 charge of these viruses was restricted to +3 and +4, although the viruses of both mothers' groups had V3 charges ranging from +2 to +5. The majority of studies to date have reported on viruses with low V3 charges being preferentially transmitted and here we describe a narrow range in V3 charges within children even when mothers contain quasispecies of viruses with lower charges. This could result from such viruses being the major variants in the TM, but could also reflect a less stable electrostatic interaction between gp120 and the array of receptors to which it binds and which may influence HIV-1 transmission, including CD4, CCR5, or DC-SIGN. 39,43 PNGS N197, N230, N241, N301, N332, N386, N392, and N397 (HXB2 numbering) have been identified as candidates to play a role in DC-SIGN binding and may therefore exert such an influence through modulating this specific interaction. 44,45
The selection for viruses with PNGS-N339 in viruses undergoing MTCT and not in acute horizontal infections could be explained by neutralizing antibody selection. Children will possess antibodies from the mothers whereas individuals sexually exposed to HIV-1 will not have such antibodies and can therefore be infected with viruses lacking PNGS-N339. The function of PNGS-N339 in the envelope of the children remains to be determined; however, PNGS-N339 is situated in the α2-helix region of C3 and several reports have demonstrated that residues within the α2-helix of subtype C viruses are under strong positive selection and that unique mutational patterns in the α2-helix and acquisition of length in gp120 hypervariable domains are associated with resistance to autologous neutralization of HIV-1 subtype C. 46 The disadvantage in MTCT for PNGS-N295 can be explained by the generation of resistance against neutralizing antibodies similar to 2G12. 47
The further clarification of mechanisms of HIV-1 MTCT and the viruses involved will provide insights into which gp120 envelope regions may need to be targeted in a vaccine aimed at restricting transmission. Therapeutic vaccination of HIV-1-positive mothers during their pregnancy against such variants may have the benefit of restricting MTCT during all possible transmission periods and allow for lower rates of infection. We demonstrate that PNGS-N234 and PNGS-N339 within the gp120 envelope is associated with the risk of viral transmission from mother to child. Knowledge concerning which viruses are preferentially transmitted may aide in the future design of HIV-1 vaccine immunogens aimed at restricting MTCT.
Footnotes
Acknowledgments
The authors would like to thank the participants for their study involvement and Dr. J.M. Ruijter for assistance with the statistical analysis.
Accession numbers
GenBank accession numbers JN187678–JN187853 and JN097489–JN097554.
Author Disclosure Statement
No competing financial interests exist.